
Поиск по сайту | Диоды Д231, Д231А, Д231Б, Д232, Д232А, Д232Б, Д233, Д233Б, Д234Б – диффузионные, кремниевые. Основное предназначение – преобразование переменного напряжения с частотой до 1 кГц. Имеют жёсткие выводы и металлостеклянный корпус. Цоколёвка диодов написана на корпусе, как и их название. Электрические параметры Д231, Д231А, Д231Б, Д232, Д232А, Д232Б, Д233, Д233Б, Д234Б
Предельные характеристики диодов Д231, Д231А, Д231Б, Д232, Д232А, Д232Б, Д233, Д233Б, Д234Б
Рабочая частота (без ухудшения электрических параметров) – 1 кГц |
 ..+130°C
..+130°CДиод Д232 — DataSheet
Корпус диодов Д231, Д232, Д233, Д234Описание
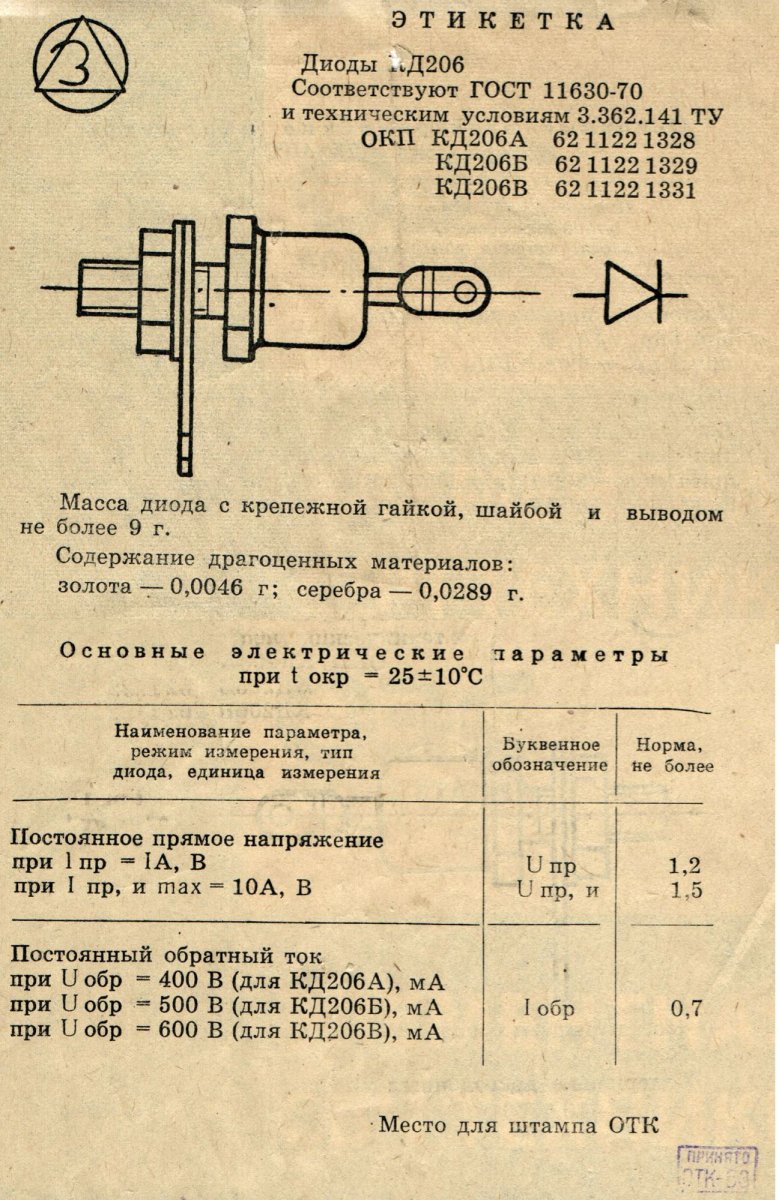
Диоды кремниевые, диффузионные. Предназначены для преобразования переменного напряжения частотой до 1,1 кГц. Выпускаются
в металлостеклянном корпусе с жесткими выводами. Тип диода и схема соединения электродов с выводами приводятся на корпусе. Масса диода с комплектующими деталями не более 18 г.
При креплении диодов усилие затяжки должно быть не более 196 Н·м (0,2 кгс·м). При этом запрещается прилагать к изолированному выводу усилие, превышающее 9,8 Н (1 кгс), что может привести к нарушению целостности стеклянного изолятора. Размеры радиатора
(теплоотвода) рассчитываются из условия, что диод является точечным источником теплоты, рассеивающим мощность 2Uпр,ср· Iпр,ср.
При последовательном соединении диодов рекомендуется применять диоды одного типа и шунтировать каждый резистором сопротивлением 10…15 кОм на каждые 100 В амплитуды обратного напряжения.
| Параметр | Обозначение | Маркировка | Значение | Ед. изм. |
| Аналог | Д232 | 10А400 | ||
| Д232А | 10А400 | |||
| Д232Б | 1N1069 | |||
| Максимальное постоянное обратное напряжение. | Uo6p max, Uo6p и max | Д232 | 400 | В |
| Д232А | 400 | |||
| Д232Б | 400 | |||
Максимальный постоянный прямой ток.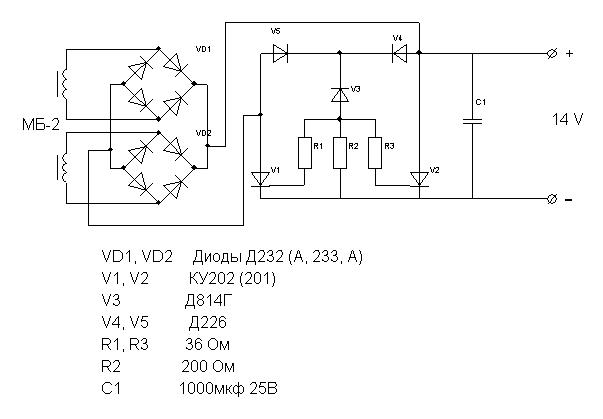 | Iпp max, Iпp ср max, I*пp и max | Д232 | 10 | А |
| Д232А | 10 | |||
| Д232Б | 5 | |||
| Максимальная рабочая частота диода | fд max | Д232 | 1.1 | кГц |
| Д232А | 1.1 | |||
| Д232Б | 1.1 | |||
| Постоянное прямое напряжение | Uпр не более (при Iпр, мА) | Д232 | 1 (10 А) | В |
| Д232А | 1 (10 А) | |||
| Д232Б | 1.5 (5 А) | |||
| Постоянный обратный ток | Iобр не более (при Uобр, В) | Д232 | 3000 (400) | мкА |
| Д232А | 3000 (400) | |||
| Д232Б | 3000 (400) | |||
| Время обратного восстановления — время переключения диода с заданного прямого тока на заданное обратное напряжение от момента прохождения тока через нулевое значение до момента достижения обратным током заданного значения | tвос, обр | Д232 | — | мкс |
| Д232А | — | |||
| Д232Б | — | |||
| Общая емкость | Сд (при Uобр, В) | Д232 | — | пФ |
| Д232А | — | |||
| Д232Б | — |
Описание значений со звездочками(*) смотрите в буквенных обозначениях параметров диодов.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Диод Д232А
Справочник количества содержания ценных металлов в диоде Д232А согласно паспорта на изделие и информационной литературы. Указано точное значение драгоценных металлов в граммах (Золото, серебро, платина, палладий и другие) на единицу изделия.
Содержание драгоценных металлов в диоде Д232А
Золото: 0,0021 грамм.
Серебро: 0 грамм.
Платина: 0 грамм.
Палладий: 0 грамм.
Источник информации: .
Фото диода Д232А:
Панель ламповая виды
Диод — электронный элемент, обладающий различной проводимостью в зависимости от направления электрического поля. Электрод диода, подключаемый к положительному полюсу источника тока, когда диод открыт (то есть имеет маленькое сопротивление), называют анодом, подключаемый к отрицательному полюсу — катодом.
О комплектующем изделии — Диод
Диод — видео.
Диод это полупроводниковый прибор основанный на PN-переходе. А если без теории, то диод в одном направлении пропускает ток, а в другом нет. Вот и все.
Как работает диод — видео.
В этом выпуске вы узнаете: что такое диод, принцип действия диода, как работает диод, что такое p — n переход; что такое прямой ток диода, что такое обратный ток диода; каково внутреннее сопротивление диода; что такое вольт- амперная характеристика диода; что такое пропускное и не пропускное напряжение диода; как работает диод в цепи постоянного тока, как работает диод в цепи переменного тока; как устроен плоскостной диод; какие существуют виды диодов; как устроен выпрямительный диод.
Характеристики диодов Д232А:
Купить или продать а также цены на Диод Д232А:
Оставьте отзыв о Д232А:
Диод Д232А
Справочник количества содержания ценных металлов в диоде Д232А согласно паспорта на изделие и информационной литературы. Указано точное значение драгоценных металлов в граммах (Золото, серебро, платина, палладий и другие) на единицу изделия.
Содержание драгоценных металлов в диоде Д232А
Золото: 0,0021 грамм.
Серебро: 0 грамм.
Платина: 0 грамм.
Палладий: 0 грамм.
Источник информации: .
Фото диода Д232А:
Панель ламповая виды
Диод — электронный элемент, обладающий различной проводимостью в зависимости от направления электрического поля. Электрод диода, подключаемый к положительному полюсу источника тока, когда диод открыт (то есть имеет маленькое сопротивление), называют анодом, подключаемый к отрицательному полюсу — катодом.
О комплектующем изделии – Диод
Диод – видео.
Диод это полупроводниковый прибор основанный на PN-переходе. А если без теории, то диод в одном направлении пропускает ток, а в другом нет. Вот и все.
Как работает диод – видео.
В этом выпуске вы узнаете: что такое диод, принцип действия диода, как работает диод, что такое p – n переход; что такое прямой ток диода, что такое обратный ток диода; каково внутреннее сопротивление диода; что такое вольт- амперная характеристика диода; что такое пропускное и не пропускное напряжение диода; как работает диод в цепи постоянного тока, как работает диод в цепи переменного тока; как устроен плоскостной диод; какие существуют виды диодов; как устроен выпрямительный диод.
Детали устройства Микросхема: DD1, DD2 – К155ЛА3. Диоды: Br1 Д232А; Br2 – Д242Б; D1 Д226Б. Стабилитрон: D2 КС156А.
1 Хитрый выпрямитель Выпрямитель предназначен для питания бытовых потребителей, которые могут работать как на переменном, так и на постоянном токе. Это например электроплиты, камины, водонагревательные устройства, освещение и т. п. Главное, чтобы в этих устройствах не было электродвигателей, трансформаторов и других элементов, рассчитанных на переменный ток. Устройство, собранное по предлагаемой схеме, просто вставляется в розетку и от него питается нагрузка. Вся электропроводка остается нетронутой. Заземление не нужно. Счетчик при этом учитывает примерно четверть потребленной электроэнергии.



2 Транзисторы: Т1 КТ848А, Т2 КТ815В, Т3 КТ315. Т1 и Т2 устанавливаются на радиаторе площадью не менее 150 см 2. Транзисторы устанавливаются на изолирующих прокладках. Конденсаторы электролитические: С1-10 мкф 400В; С мкф 50В; С мкф 16В; Конденсаторы высокочастотные: С2, С3 0.1 мкф. Резисторы: R1, R2 27 ком; R3 56 Ом; R4 3 ком; R5-22 ком; R6 10 Ом; R7, R8 1.5 ком; R9 560 Ом. Резисторы R3, R6 проволочные мощностью не менее 10 Вт, R9 – типа МЛТ-2, остальные резисторы МЛТ Трансформатор Tr1 любой маломощный 220/36 В. Наладка При наладке схемы соблюдайте осторожность! Помните, что низковольтная часть схемы не имеет гальванической развязки от электрической сети! Не рекомендуется в качестве радиатора для транзисторов использовать металлический корпус устройства. Применение плавких предохранителей обязательно! Вначале проверяют отдельно от схемы низковольтный блок питания. Он должен обеспечивать ток не менее 2 А на выходе 36 В, а также 5 В для питания маломощного генератора. Затем налаживают генератор, отключив силовую часть схемы от электросети (для этого можно временно отсоединить резистор R6). Генератор должен формировать импульсы амплитудой 5 В и частотой около 2 кгц. Скважность импульсов приблизительно 1/1. При необходимости для этого подбирают конденсаторы С2, С3 или резисторы R7, R8. Формирователь импульсов на транзисторах Т2 и Т3, если правильно собран, обычно наладки не требует. Но желательно убедиться, что он способен обеспечить импульсный ток базы транзистора Т1 на уровне А. Если такое значение тока не обеспечить, транзистор Т1 не будет в открытом состоянии входить в режим насыщения и сгорит за несколько секунд. Для проверки этого режима можно при отключенной силовой части схемы и отключенной базе транзистора Т1, вместо резистора R1 включить шунт сопротивлением в несколько Ом. Импульсное напряжение на шунте при включенном генераторе регистрируют осциллографом и пересчитывают на значение тока. При необходимости подбирают сопротивления резисторов R2, R3 и R4.
Затем налаживают генератор, отключив силовую часть схемы от электросети (для этого можно временно отсоединить резистор R6). Генератор должен формировать импульсы амплитудой 5 В и частотой около 2 кгц. Скважность импульсов приблизительно 1/1. При необходимости для этого подбирают конденсаторы С2, С3 или резисторы R7, R8. Формирователь импульсов на транзисторах Т2 и Т3, если правильно собран, обычно наладки не требует. Но желательно убедиться, что он способен обеспечить импульсный ток базы транзистора Т1 на уровне А. Если такое значение тока не обеспечить, транзистор Т1 не будет в открытом состоянии входить в режим насыщения и сгорит за несколько секунд. Для проверки этого режима можно при отключенной силовой части схемы и отключенной базе транзистора Т1, вместо резистора R1 включить шунт сопротивлением в несколько Ом. Импульсное напряжение на шунте при включенном генераторе регистрируют осциллографом и пересчитывают на значение тока. При необходимости подбирают сопротивления резисторов R2, R3 и R4. Следующей стадией является проверка силовой части. Для этого восстанавливают все соединения в схеме. Конденсатор С1 временно отключают, а в качестве нагрузки используют потребитель малой мощности, например лампу накаливания мощностью до 100 Вт. При включении устройства в электрическую сеть действующее значение напряжения на нагрузке должно быть на уровне В. Осциллограммы напряжения на нагрузке и на резисторе R6 должны показать, что питание её производится импульсами с частотой, задаваемой генератором. Если всё исправно, подключают конденсатор С1, только вначале емкость его принимают в несколько раз меньше номинальной (например 0.1 мкф). Действующее напряжение на нагрузке заметно возрастает и при последующем увеличении емкости С1 достигает 310 В. При этом очень важно внимательно следить за температурой транзистора Т1. Если возникает повышенный нагрев при использовании маломощной нагрузки, это свидетельствует о том, что Т1 либо не входит в режим насыщения в открытом состоянии, либо
Следующей стадией является проверка силовой части. Для этого восстанавливают все соединения в схеме. Конденсатор С1 временно отключают, а в качестве нагрузки используют потребитель малой мощности, например лампу накаливания мощностью до 100 Вт. При включении устройства в электрическую сеть действующее значение напряжения на нагрузке должно быть на уровне В. Осциллограммы напряжения на нагрузке и на резисторе R6 должны показать, что питание её производится импульсами с частотой, задаваемой генератором. Если всё исправно, подключают конденсатор С1, только вначале емкость его принимают в несколько раз меньше номинальной (например 0.1 мкф). Действующее напряжение на нагрузке заметно возрастает и при последующем увеличении емкости С1 достигает 310 В. При этом очень важно внимательно следить за температурой транзистора Т1. Если возникает повышенный нагрев при использовании маломощной нагрузки, это свидетельствует о том, что Т1 либо не входит в режим насыщения в открытом состоянии, либо
3 полностью не закрывается.![]() В этом случае следует вернуться к настройке формирователя импульсов. Эксперименты показывают, что при питании нагрузки мощностью 100 Вт без конденсатора С1, транзистор Т1 в течение длительного времени не нагревается даже без радиатора. В заключении подключается номинальная нагрузка и подбирается емкость С1 такая, чтобы обеспечить питание нагрузки постоянным напряжением 220 В. Емкость С1 следует подбирать осторожно, начиная с малых значений, так как увеличение емкости приводит к увеличению выходного напряжения (до 310 В, что может вывести из строя нагрузку), а также резко увеличивает импульсный ток через транзистор Т1. Об амплитуде импульсов тока через Т1 можно судить, подключив осциллограф параллельно резистору R6. Импульсный ток должен быть не более допустимого для выбранного транзистора (20 А для КТ848А). В случае необходимости его ограничивают, увеличивая сопротивление R6, но лучше остановиться на меньшем значении емкости С1. При указанных деталях устройство рассчитано на нагрузку 1 квт.
В этом случае следует вернуться к настройке формирователя импульсов. Эксперименты показывают, что при питании нагрузки мощностью 100 Вт без конденсатора С1, транзистор Т1 в течение длительного времени не нагревается даже без радиатора. В заключении подключается номинальная нагрузка и подбирается емкость С1 такая, чтобы обеспечить питание нагрузки постоянным напряжением 220 В. Емкость С1 следует подбирать осторожно, начиная с малых значений, так как увеличение емкости приводит к увеличению выходного напряжения (до 310 В, что может вывести из строя нагрузку), а также резко увеличивает импульсный ток через транзистор Т1. Об амплитуде импульсов тока через Т1 можно судить, подключив осциллограф параллельно резистору R6. Импульсный ток должен быть не более допустимого для выбранного транзистора (20 А для КТ848А). В случае необходимости его ограничивают, увеличивая сопротивление R6, но лучше остановиться на меньшем значении емкости С1. При указанных деталях устройство рассчитано на нагрузку 1 квт. Применяя другие элементы силового выпрямителя и транзисторный ключ соответствующей мощности, можно питать и более мощные потребители. Обращаем Ваше внимание на то, что при изменении нагрузки, напряжение на ней также будет существенно изменяться. Поэтому устройство целесообразно настроить и использовать постоянно с одним и тем же потребителем. Этот недостаток в определенных случаях может оказаться достоинством. Например, изменяя емкость С1 можно в широких пределах регулировать мощность нагревательных приборов.
Применяя другие элементы силового выпрямителя и транзисторный ключ соответствующей мощности, можно питать и более мощные потребители. Обращаем Ваше внимание на то, что при изменении нагрузки, напряжение на ней также будет существенно изменяться. Поэтому устройство целесообразно настроить и использовать постоянно с одним и тем же потребителем. Этот недостаток в определенных случаях может оказаться достоинством. Например, изменяя емкость С1 можно в широких пределах регулировать мощность нагревательных приборов.
4
5 Радиолюбительская алхимия Через устройство, согласно принципиальной электрической схемы, изображенной на рис.1, можно включить электрический чайник в розетку совершенно незаметно для электрического счетчика. Или подключить любой электрический прибор не требовательный к форме питающего напряжения достаточно большой потребляемой мощности. Если заведомо известна другая пониженная потребляемая мощность электрического прибора, то согласно справочным данным по полупроводниковым приборам можно подобрать и другие параметры радиодеталей для этой схемы. Как работает эта схема? После включения питания сетевое напряжение поступает одновременно на мощные диоды VD1 и первичные обмотки адаптеров для зарядки сотовых телефонов. Если в момент включения регулятора в сети оказалось напряжение отрицательной полярности, ток нагрузки протекает по цепи эмиттер-коллектор VT1. Если полярность сетевого напряжения положительная, ток протекает по цепи коллектор-эмиттер VT1. Значение тока нагрузки зависит от величины управляющего напряжения на базе VT1. Управляющее напряжение формируется генератором на логических элементах (микросхема К155ЛА3 или КМ155ЛА3). Частота генератора – 2кГц, скважность – 50%. Таким образом, наш электрический чайник превращается в высокочастотную (с точки зрения счетчика) нагрузку и электрический счетчик её не видит. Остается только в нужный момент открывать транзистор и счетчик начнет крутиться так как Вам надо. Параллельно нагрузке включается конденсатор (на схеме показан как С1) – это улучшит форму напряжения подаваемого на нагрузку.
Как работает эта схема? После включения питания сетевое напряжение поступает одновременно на мощные диоды VD1 и первичные обмотки адаптеров для зарядки сотовых телефонов. Если в момент включения регулятора в сети оказалось напряжение отрицательной полярности, ток нагрузки протекает по цепи эмиттер-коллектор VT1. Если полярность сетевого напряжения положительная, ток протекает по цепи коллектор-эмиттер VT1. Значение тока нагрузки зависит от величины управляющего напряжения на базе VT1. Управляющее напряжение формируется генератором на логических элементах (микросхема К155ЛА3 или КМ155ЛА3). Частота генератора – 2кГц, скважность – 50%. Таким образом, наш электрический чайник превращается в высокочастотную (с точки зрения счетчика) нагрузку и электрический счетчик её не видит. Остается только в нужный момент открывать транзистор и счетчик начнет крутиться так как Вам надо. Параллельно нагрузке включается конденсатор (на схеме показан как С1) – это улучшит форму напряжения подаваемого на нагрузку. Емкость придется подбирать экспериментально, рекомендую использовать только высоковольтные конденсаторы. Можно на выходе применить более мощный транзистор, например, КТ848А. Детали Чтобы не мучиться с блоком питания в данной конструкции применялись два адаптера для зарядки сотовых телефонов с напряжением на выходе 7 В и 5 В. В частности для питания микросхемы применялся отдельный блок или адаптер на 5 В. Не было необходимости в стабилитроне. Конденсатор С1 применить не менее 400 В. На транзисторы обязательно установить отдельные радиаторы. На VТ1 площадь не менее 200 кв.см. А на VТ2 площадь радиатора не менее 50 кв. см. Сопротивления R1, R2 и R3 типа МЛТ-2. Заменить можно VD1 на КД2989А, КД2989А1, КД2989Б, КД2989Б1, КД203М, КД203Л. При необходимости устанавливается стабилитрон КС407Гили КС451А. Схема собиралась навесным монтажом и заработала сразу.
Емкость придется подбирать экспериментально, рекомендую использовать только высоковольтные конденсаторы. Можно на выходе применить более мощный транзистор, например, КТ848А. Детали Чтобы не мучиться с блоком питания в данной конструкции применялись два адаптера для зарядки сотовых телефонов с напряжением на выходе 7 В и 5 В. В частности для питания микросхемы применялся отдельный блок или адаптер на 5 В. Не было необходимости в стабилитроне. Конденсатор С1 применить не менее 400 В. На транзисторы обязательно установить отдельные радиаторы. На VТ1 площадь не менее 200 кв.см. А на VТ2 площадь радиатора не менее 50 кв. см. Сопротивления R1, R2 и R3 типа МЛТ-2. Заменить можно VD1 на КД2989А, КД2989А1, КД2989Б, КД2989Б1, КД203М, КД203Л. При необходимости устанавливается стабилитрон КС407Гили КС451А. Схема собиралась навесным монтажом и заработала сразу.
6 Уважаемые читатели сайта, медики-радиолюбители! За год мне, как автору этой статьи, поступило более полусотни писем с просьбой разъяснить тонкости конструкции этого устройства. С учетом ваших вопросов и публикуется это дополнение.
С учетом ваших вопросов и публикуется это дополнение.
7 Сопротивления R1, R3-240 Ом, 0,25 Вт, R2, R4 10 ком (сп3-4), R5, R6 1кОм 2Вт, R7 300 Ом 2 Вт. С2, С3, С4, С5 электролитические 4,7 мкф 40 В, С6 0,01 мкф. С1 – самый основной конденсатор, т.к. он формирует синусоидальную форму высокочастотного тока на нагрузке. Подборка его начинается с самых малых емкостей, указанных в схеме. А при первичной настройке конденсатор С1 не устанавливается вовсе. Но тогда подключают нагрузку не более 60 Вт. Не включайте прибор без нагрузки! Ток нарастает мгновенно и мощный транзистор перегорает. Данная схема неприемлема для питания приборов, у которых имеется электродвигатель (но при более сложной схемотехнике и эта проблема устраняется). Вначале проверяют отдельно от схемы низковольтный блок питания микросхемы КМ155ЛА3 (генератора прямоугольных импульсов). Резистором R2 выставляют стабилизированное напряжение +5 В. И на этом вся регулировка заканчивается. Налаживание блока питания +7 В имеет свои сложности.![]() Резистором R4 так же выставляется стабилизированное напряжение +7 В. Затем может не хватать силы тока одного адаптера. Тогда подключают параллельно еще один адаптер плюс к плюсу, минус к минусу. Возможно, что таких подключений буде четыре, тогда и добьетесь силу тока в 1А на коллекторе транзистора КТ602БМ. Если на транзисторе (формирователе прямоугольных импульсов – по схеме КТ602БМ) нет тока, то необходимо добавить усилитель тока на транзисторе КТ315 любой серии. Схему лучше собирать навесным монтажом. И только после всех мучений вести монтаж на плате. На печатных платах необходимо токоведущие дорожки делать как можно шире. И не гонитесь за миниатюризацией прибора, так как по шинам протекают очень большие токи! Устройство до такой степени капризное, что не терпит даже смены нагрузки. В результате чего приходится все настраивать заново. Пайку всех элементов осуществлять заземленным паяльником не более 40 Вт и не более 3-х секунд, базу транзисторов паять в первую очередь. Диодный мост на Д203 неприхотлив и выдерживает любые издевательства.
Резистором R4 так же выставляется стабилизированное напряжение +7 В. Затем может не хватать силы тока одного адаптера. Тогда подключают параллельно еще один адаптер плюс к плюсу, минус к минусу. Возможно, что таких подключений буде четыре, тогда и добьетесь силу тока в 1А на коллекторе транзистора КТ602БМ. Если на транзисторе (формирователе прямоугольных импульсов – по схеме КТ602БМ) нет тока, то необходимо добавить усилитель тока на транзисторе КТ315 любой серии. Схему лучше собирать навесным монтажом. И только после всех мучений вести монтаж на плате. На печатных платах необходимо токоведущие дорожки делать как можно шире. И не гонитесь за миниатюризацией прибора, так как по шинам протекают очень большие токи! Устройство до такой степени капризное, что не терпит даже смены нагрузки. В результате чего приходится все настраивать заново. Пайку всех элементов осуществлять заземленным паяльником не более 40 Вт и не более 3-х секунд, базу транзисторов паять в первую очередь. Диодный мост на Д203 неприхотлив и выдерживает любые издевательства. Вполне возможно, придется обратиться за помощью к более опытному электронщику. И еще, если есть желание сделать механический прерыватель с частотой 1-2 кгц, то вообще нет необходимости в монтаже блоков питания генератора импульсов, каскадов формирования прямоугольных импульсов. Всем удачи и терпения при конструировании!
Вполне возможно, придется обратиться за помощью к более опытному электронщику. И еще, если есть желание сделать механический прерыватель с частотой 1-2 кгц, то вообще нет необходимости в монтаже блоков питания генератора импульсов, каскадов формирования прямоугольных импульсов. Всем удачи и терпения при конструировании!
8 А.Тумарев, UA3GLU Природа эффекта полостных структур Какова же физическая природа ЭПС? Было высказано немало предположений и гипотез; к сожалению, многие из них отдают экстрасенсурой, столь почему то модной среди интеллигенции в наши дни. Наибольшего внимания заслуживает теория ленинградского физика, доктора технических наук В.Ф. Золотарева, разработанная им еще ранее, а сейчас получившая убедительное экспериментальное подтверждение. В результате длительных совместных исследований мы охарактеризовали находку как “неизвестное ранее явление взаимодействия многополостных структур с живыми системами,заключающееся в том, что сопутствующие движению электронных потоков в твердых стенках полостей волны де Бройля образуют посредством интерференции макроскопическое поле многополостных структур, вызывающие изменения функционального состояния живых объектов, находящихся в этом поле”. Волны де Бройля присущи движущимся микрочастицам любого тела, в толще его скомпенсированы, на поверхности же проявляются в виде излучения, но настолько коротковолнового и сверхвысокочастотного, что приборами были уловлены лишь в виде дифракции, но тут же помогли науке: вспомним своеобразные портреты электронов и нейтронов, полученные на кристаллах и пленках именно с помощью волн де Бройля; никто не думал, что эти мизерные излучения могут как’то воздействовать на живое. И они не воздействовали ‘ во всяком случае возле плоских предметов. Зато у многополостных структур, где площадь поверхности твердых тел велика, к тому же многократно искривлена, волны де Бройля складываются, образуя, подобно музыкальным обертонам, гармоники с уже меньшими частотами. Так, удлиняясь и усиливаясь за счет взаимоналожения в ячейках, они образуют “пучности”‘максимумы стоячих волн де Бройля. Наталкиваясь на эти сами по себе пассивные преграды, нервные импульсы дают сбои, меняя свою частоту и скорость и вызывая не только кажущиеся ощущения, но порой и существенные физиологические изменения.
Волны де Бройля присущи движущимся микрочастицам любого тела, в толще его скомпенсированы, на поверхности же проявляются в виде излучения, но настолько коротковолнового и сверхвысокочастотного, что приборами были уловлены лишь в виде дифракции, но тут же помогли науке: вспомним своеобразные портреты электронов и нейтронов, полученные на кристаллах и пленках именно с помощью волн де Бройля; никто не думал, что эти мизерные излучения могут как’то воздействовать на живое. И они не воздействовали ‘ во всяком случае возле плоских предметов. Зато у многополостных структур, где площадь поверхности твердых тел велика, к тому же многократно искривлена, волны де Бройля складываются, образуя, подобно музыкальным обертонам, гармоники с уже меньшими частотами. Так, удлиняясь и усиливаясь за счет взаимоналожения в ячейках, они образуют “пучности”‘максимумы стоячих волн де Бройля. Наталкиваясь на эти сами по себе пассивные преграды, нервные импульсы дают сбои, меняя свою частоту и скорость и вызывая не только кажущиеся ощущения, но порой и существенные физиологические изменения. Своей энергии стоячие волны де Бройля не несут, и закон сохранения энергии ни в коей мере не нарушается. Поскольку волны де Бройля распространяются в физическом вакууме, ЭПС должен обладать всепроницающим действием. Именно это и наблюдаем мы при безуспешном перекрытии ЭПС любым экраном. Что такое широкополосный трансформатор?
Своей энергии стоячие волны де Бройля не несут, и закон сохранения энергии ни в коей мере не нарушается. Поскольку волны де Бройля распространяются в физическом вакууме, ЭПС должен обладать всепроницающим действием. Именно это и наблюдаем мы при безуспешном перекрытии ЭПС любым экраном. Что такое широкополосный трансформатор?
9 Самодельный трансивер на 144 мгц. Смеситель типовой – трансформаторы на колечках, диоды Вопрос в кольцах. Согласно справочнику 50 ВЧ работают до 30 МГц. 20 ВЧ – немного более. А вот на 144 МГц вопрос! Так вот, на этих кольцах трансформаторы потянут ли? В этих нескольких строчках заключены вопросы, ответы на которые, казалось бы, очевидны для большинства радиолюбителей. Бери кольцо, да мотай! Однако, не все так просто в понимании работы широкополосных трансформаторов. Пришлось собрать в уме все то, чему меня учили, свой практический опыт и дать ответ По поводу ферритовых колец в смесителе в данном случае, и вообще о широкополосных трансформаторах на длинных линиях (ШПТЛ), существует множество глубоких и устойчивых заблуждений. Здесь как раз одно из них! Хотя, если “потянуть ниточку, то концов в этом “клубочке заблуждений окажется очень много. Известно, что в старых распространенных радиолюбительских конструкциях всегда рекомендовались ферриты с проницаемостью А они ведь очень низкочастотные! Однако же в каком нибудь “Радио-76 они стоят и на входе и во всех смесителях. Что, авторы этих конструкций, известные радиолюбители, совершили ошибку? Отнюдь! Они то помнили и понимали, что энергия в ШПТЛ-ах передается не через перемагничивание сердечника, а непосредственно от элемента линии к элементу. Феррит здесь нужен для того, что бы повысить сопротивление линии для синфазных токов и в качестве “сборщика полей рассеивания. Т.е., поглотителя энергии, которая паразитно наводится вокруг линии. Я, например, в своих конструкциях на КВ часто использую ферритовые кольца НМ2000. Это не значит, что надо применять только такие ферриты. Я хочу сказать, что и с такими магнитопроводами трансформаторы вполне нормально работают в широкой полосе радиочастот.
Здесь как раз одно из них! Хотя, если “потянуть ниточку, то концов в этом “клубочке заблуждений окажется очень много. Известно, что в старых распространенных радиолюбительских конструкциях всегда рекомендовались ферриты с проницаемостью А они ведь очень низкочастотные! Однако же в каком нибудь “Радио-76 они стоят и на входе и во всех смесителях. Что, авторы этих конструкций, известные радиолюбители, совершили ошибку? Отнюдь! Они то помнили и понимали, что энергия в ШПТЛ-ах передается не через перемагничивание сердечника, а непосредственно от элемента линии к элементу. Феррит здесь нужен для того, что бы повысить сопротивление линии для синфазных токов и в качестве “сборщика полей рассеивания. Т.е., поглотителя энергии, которая паразитно наводится вокруг линии. Я, например, в своих конструкциях на КВ часто использую ферритовые кольца НМ2000. Это не значит, что надо применять только такие ферриты. Я хочу сказать, что и с такими магнитопроводами трансформаторы вполне нормально работают в широкой полосе радиочастот. Какие же условия должны соблюдаться для того, чтобы трансформатор был именно на длинных линиях? 1) Его обмотки должны представлять собой длинные линии с известным волновым сопротивлением. Проще говоря, все “обмотки трансформатора должны быть сделаны из параллельных или слегка скрученных проводов с одинаковыми расстояниями между ними. Конструкции трансформаторов, которые выполнены “традиционным способом (первичная обмотка на одной части кольца, вторичная на другой) НЕ РАБОТОСПОСОБНЫ! В этом можно убедиться, сделав простой эксперимент. Намотайте трансформатор на кольце с коэффициентом трансформации 1:1 или 1:2 (эти цифры еще один повод для обсуждения) и нагрузите на соответственный эквивалент нагрузки, сделанный, например, из резистора МЛТ-2. В первом случае – это 50 Ом, а во втором Ом. Подайте на трансформатор постоянный сигнал небольшой мощности с любого современного трансивера, используя его, как ГСС. Так вот, когда трансформатор намотан “тради-
Какие же условия должны соблюдаться для того, чтобы трансформатор был именно на длинных линиях? 1) Его обмотки должны представлять собой длинные линии с известным волновым сопротивлением. Проще говоря, все “обмотки трансформатора должны быть сделаны из параллельных или слегка скрученных проводов с одинаковыми расстояниями между ними. Конструкции трансформаторов, которые выполнены “традиционным способом (первичная обмотка на одной части кольца, вторичная на другой) НЕ РАБОТОСПОСОБНЫ! В этом можно убедиться, сделав простой эксперимент. Намотайте трансформатор на кольце с коэффициентом трансформации 1:1 или 1:2 (эти цифры еще один повод для обсуждения) и нагрузите на соответственный эквивалент нагрузки, сделанный, например, из резистора МЛТ-2. В первом случае – это 50 Ом, а во втором Ом. Подайте на трансформатор постоянный сигнал небольшой мощности с любого современного трансивера, используя его, как ГСС. Так вот, когда трансформатор намотан “тради-
10 ционным способом, то он дает КСВ на входе, равный БЕСКОНЕЧНОСТИ! А когда ваш трансформатор по конструкции – истинный ШПТЛ, то КСВ будет около 1 и в широком диапазоне частот. Опыт можно повторить с различными ферритами. Такой эксперимент очень показателен, его можно проделать не выходя из дома, на своем рабочем столе; 2) ШПТЛ должен быть нагружен по входу и выходу на АКТИВНЫЕ нагрузки равные примерно волновому сопротивлению линий из которых он сделан. Типовой пример: Наш брат-радиолюбитель применяет для “симметрирования антенн огромные по величине ферритовые кольца возле полотна. Однако, описанный выше эксперимент с активными нагрузками показывает, что колечко диаметром в мм выдерживает мощность в 100 Вт и не нагревается! Так где же правда? Правда в том, что антенна (диполь или рамка) имеет низкое активное сопротивление ТОЛЬКО на одной единственной частоте, частоте первой гармоники антенны. Высокие активные сопротивления, которые имеются на четных гармониках, на практике неприменимы. Низкоомные резонансы на нечетных верхних гармониках попадают уже не в радиолюбительские диапазоны. А на остальных частотах ВСЕГДА будут присутствовать значительные реактивности.
 Они вызывают сильный нагрев кольца и поэтому оно должно иметь большую поверхность охлаждения, т.е., быть БОЛЬШИМ. К примеру, в импортных стоваттных трансиверах на выходе ПА стоят микроскопические ферритовые бинокли. И… НИЧЕГО! Это не из-за того, что они сделаны из диковинного материала. Просто одно из требований к выходной нагрузке для таких трансиверов – что бы она была АКТИВНОЙ. (Другое требование 50 Ом). Следует опасаться тех публикаций, где рекомендуют мотать строго определенное число витков для ВЧ трансформатора. Это признак еще одной “болезни сознания – квазирезонансного использования ШПТЛ-а. Вот оттуда “ростут ноги у легенды о необходимости применять ВЧ ферриты. Но Широкополосности-то уже НЕТ! Теперь про упомянутые 1:1 и 1:2… В школьном курсе физики коэффициент трансформации – это соотношение витков первичной и вторичной обмоток. Т.е., соотношение входных и выходных напряжений. Почему же у радиолюбителей этот параметр превратился “по умолчанию в коэффициент трансформации сопротивлений? Да потому, что трансформация сопротивлений более важна в нашей среде. Но не следует доходить до абсурда! Вот разговор подслушанный в эфире два радиолюбителя обсуждают как сделать трансформатор с 50 на 75 Ом. Один предлагает мотать его с соотношением витков 1:1,5. И когда им кто-то робко возражает, в ответ слышны только обвинения в технической неграмотности. И подобное случается на каждом шагу! А это всего лишь – ТЕРМИНЫ! Получается, что великий закон сохранения энергии для них не действует и можно при напря-
Они вызывают сильный нагрев кольца и поэтому оно должно иметь большую поверхность охлаждения, т.е., быть БОЛЬШИМ. К примеру, в импортных стоваттных трансиверах на выходе ПА стоят микроскопические ферритовые бинокли. И… НИЧЕГО! Это не из-за того, что они сделаны из диковинного материала. Просто одно из требований к выходной нагрузке для таких трансиверов – что бы она была АКТИВНОЙ. (Другое требование 50 Ом). Следует опасаться тех публикаций, где рекомендуют мотать строго определенное число витков для ВЧ трансформатора. Это признак еще одной “болезни сознания – квазирезонансного использования ШПТЛ-а. Вот оттуда “ростут ноги у легенды о необходимости применять ВЧ ферриты. Но Широкополосности-то уже НЕТ! Теперь про упомянутые 1:1 и 1:2… В школьном курсе физики коэффициент трансформации – это соотношение витков первичной и вторичной обмоток. Т.е., соотношение входных и выходных напряжений. Почему же у радиолюбителей этот параметр превратился “по умолчанию в коэффициент трансформации сопротивлений? Да потому, что трансформация сопротивлений более важна в нашей среде. Но не следует доходить до абсурда! Вот разговор подслушанный в эфире два радиолюбителя обсуждают как сделать трансформатор с 50 на 75 Ом. Один предлагает мотать его с соотношением витков 1:1,5. И когда им кто-то робко возражает, в ответ слышны только обвинения в технической неграмотности. И подобное случается на каждом шагу! А это всего лишь – ТЕРМИНЫ! Получается, что великий закон сохранения энергии для них не действует и можно при напря-
11 жении на входной обмотке, предположим 1 Вольт, подавая на 50- тиомный вход трансформатора мощность 20 мвт, на 75-тиоммном выходе снимать уже 30 мвт. Вот такой “вечный двигатель получается! Здесь всего-то лишь надо помнить, что коэффициент трансформации сопротивлений находится в квадратичной зависимости от коэффициента трансформации напряжений. Другими словами, трансформатор 1:2 будет трансформировать сопротивление 50 Ом в 200 Ом, а трансформатор 5:6 сопротивление 50 Ом в 75 Ом. Почему я написал 5:6, а не 1:1,2? Вот здесь один шаг до конструкции. Как уже говорилось, ШПТЛ должен мотаться линией. А линия это два или несколько сложенных вместе и слегка скрученных провода. Волновое сопротивление такой линии зависит от диаметра проводов, расстояния между их центрами и шага скрутки. Для трансформации 50 Ом в 75 Ом необходимо использовать линию из ШЕСТИ проводов и, если нет требования к симметрированию, соединить эти провода по схеме на рис.1 Рис.1 Как вы заметили, схема тоже нарисована по особому, не как обычный трансформатор. Такое изображение лучше отражает суть конструкции. Привычное схемное изображение, рис.2, и, соответственно, “традиционная конструкция автотрансформатора с однослойной обмоткой и отводом от 0,83 общего количества витков при практических испытаниях “на столе показывает гораздо худшие результаты по широкополосности. Рис.2
12 По конструктивным и эксплуатационным соображениям нежелательно также делать ШПТЛ с укороченным участком одной из линий (рис.3). Несмотря на то, что это позволяет легко делать любые, даже дробные, коэффициенты трансформации. Такое решение приводит к появлению неоднородности в линии, вследствие чего ухудшается широкополосность. Рис.3 Интересный вопрос: “Какие предельные коэффициенты трансформации можно получить в ШПТЛ? Особенно интересно найти ответ на этот вопрос тем, кто “заболел идеей сделать широкополосный апериодический ламповый усилитель мощности, где необходимо трансформировать сопротивление порядка 1..2 ком со стороны лампы в сопротивление 50 Ом. Эксперимент “на столе дает довольно интересный результат. Опять здесь все зависит от конструкции обмоток. К примеру, если сделать “традиционный трансформатор или автотрансформатор с коэффициентом трансформации, предположим, 1:10, нагрузить его на положенное активное сопротивление, равное 5 ком и промерить КСВ на пятидесятиомной стороне, то от результата волосы могут встать дыбом! А если в добавок снять АЧХ, то будет понятно, что от широкополосности ничего не осталось. Имеется один явный, довольно острый резонанс, обусловленный индуктивностью. Рис.4
13 Эту больную тему можно было бы еще развивать до бесконечности, но Все затмила конструкция широкополосного симметрирующего трансформатора на трансфлюксоре (двухдырочном ферритовом сердечнике) показанная на рис.4, которую мне удалось “подсмотреть в импортной антенне для телевизора типа “усы. Изображение на рисунке, конечно, схематическое – на самом деле обмотки состоят из нескольких (3…5) витков. Долго с недоумением я рассматривал его конструкцию, пытаясь понять систему намотки. Наконец удалось нарисовать расположение “обмоток. Вот уж пример использования истинных длинных линий! Если бы я не знал, что это линии, то подумал бы, что я сумасшедший! Особенно эта красная короткозамкнутая обмотка Но, почему же мы не удивляемся в случае, когда, например в кабельном U- колене, необходимо соединить в одной точке оплетку с двух концов коаксиального кабеля. Тоже ведь ЛИНИЯ! При настольном эксперименте на эквивалент нагрузки этот микротрансформатор, предназначенный для работы на частотах в сотни мегагерц, показал великолепные результаты на значительно более низких частотах, вплоть до диапазона 40 м и при полной мощности трансивера. Короче! Делайте свой смеситель на любом кольце с НЧ ферритом. Испытаете – напишите! Экспериментируйте смелее! Сергей Макаркин, RX3AKT Всем известно, что время суток довольно значительно влияет на работоспособность мозга. Проявляется это в колебаниях способностей человека к восприятию и усвоению информации. Теперь эти колебания научно доказаны учеными из австралийского университета Аделаиды. Из их исследований ясно, что чем ближе к ночи, тем все более производительно начинает функционировать мозг человека. Экспериментаторы доказали, что способность мозга, например, управлять движениями рук зависит от времени суток. Ночные показатели сильно отличались от дневных. Безусловно, это открытие чрезвычайно полезно, поскольку позволяет стимулировать мозговые функции и может использоваться для реабилитационной терапии больных с инсультом для скорейшего восстановления познавательных и мыслительных способностей. Раньше считалось, что человек лучше всего работает утром. Теперь неврологи утверждают, что лучше всего человеческий мозг работает по вечерам, а наихудшее время для работы это утренние часы. В свою очередь, исследователи из Стэнфорда выяснили, что человеческий мозг лучше всего работает в условиях полной тишины, несмотря на то, что многие утверждают, что с трудом перено-
14 сят тишину, и им просто необходим хоть какой-нибудь звуковой фон. Например, если человеку проигрывать музыку с паузами и в это время снимать показатели работы мозга, то именно переходы между музыкальными частями помогают мозгу создать идеальные условия для работы человеческого мозга, и пик мозговой деятельность приходится именно на периоды тишины между музыкальными фрагментами. Исследователи из Университета штата Иллинойс изобрели новые микрочипы, которые позволят любой электронике, питающейся от аккумуляторов, работать до 100 раз дольше. Всё объясняется просто: изменится технология изготовления микрочипов. По прогнозам, смартфонам и ноутбукам подзарядка не потребуется в течение нескольких недель. Пользователям эта новость пришлась по душе, ведь хлопотно постоянно заряжать аккумуляторы, да и зачастую просто забываешь об этом, вспоминая как всегда в самый неподходящий момент. В состав нынешних чипов памяти входят тонкие металлические полоски. Через них ток пропускается каждый раз, когда устройство обращается к хранимым в памяти данным. Главным образом это и расходует заряд батареи. По новой технологии металл заменят углеродные нанотрубки. Исследователи из Иллинойского университета полагают, что в этом случае чипы станут гораздо компактнее, а соответственно потери энергии сократятся. Руководитель исследования, профессор Эрик Поп, даёт еще более дальновидные прогнозы. Он говорит, что в будущем для питания мобильных устройств будет достаточно кинетической энергии, солнечного света или тепла человеческого тела. Только додумался вытравить фрактальную антенну в виде свастики имено для качера, но так они и лежат. Но то что качер это импульсный широкополосный передатчик и приемник, это факт. Я давно начал подозревать что все дело в антеннах и именно в широкополосных. Может есть наработки, в широкополосных игольчатых антеннах. Просто они однонаправленные, хотя бы намек. Рефлектор в антенне фокусирует только одну частоту. Рефлектор для широкополосной антенне (импульсной), должен фокусировать на линейном проводнике. Или по другому. Трансформатор Тесла в первичке генерирует импульсный ток, но во вторичке он уже генерит ток в резонансе, то есть переменный. Можно ли перенести импульсный ток на
15 растояние. возбудив переменный ток допустим через один метр. Как эти широкополсные антенны этот ток отыграют. Посмотрел фильм о фрактальных антеннах, предоставленный ОТВ. Прошла аналогия. А что если вытравить на плате, не просто фрактальную антенну, а русскую руну. Облучить ее импульсным шумовым генератором (качером) и снять с нее спектр частот? А еще можно попробовать вытравить на плате копию “кругов на полях” и то же снять с этой картинки спектр. Интересно, что получится? А вообще то можно посмотреть на ютубе фильм “HAARP Климатическое оружие “. Так вот там, все делается тоже самое, что мы делаем на батарейках, только на коленке. Мы сделали первичку трансформатора Тесла, + импульсный шумовой генератор (качер), причем работающий в УКВ диапазоне. Практически, это та же микроволновка. Только там стоит магнетрон или ламповый диод, фактически тот же разрядник, что у Теслы только с постоянным магнитом. А мы это дело сделали на транзисторах. Но приложения качера, не перестают удивлять. А теперь повод для повторения схемы и генератора “infiniti”, выше она приложена. Сетевой кабель на тороидальной магнитной антенне. Я его быстренько сделал, благо дело все есть. Подключил к восьми жилам диоды помоему Д223 на 150 вольт, импульсные, и сразу Ц- шку, измерить, что он дает, на всех жилах
16 сразу.а дает он странности. Вольтаж дает нормально по постоянному току. К примеру 15 вольт. А вот ток постоянный он не видит. Нет тока! Но Ц-шку выбивает. Может это у меня так? Я не знаю. Может кто попробует? Деталей помоему, на сто рублев. Ко всем участникам форума. С конденсатором проще, он чего то копит и выдает уже нормальный ток. Правда конденсатор от конденсатора отличается. Не на каждом это происходит. Привет wowan, там кстати есть разница как подключать диод… на схеме у меня справа, а у тебя может оказаться и слева и нужно ещё перевернуть=) хотя я думаю ты пробовал по разному… я прост сказал на всякий случай дак вот о чем я… подключил я обычный блокинг (1,2в) к этим ферритам что на схеме… запустил и подключил к одной из пар светодиод… добился регулировкой резистора резонанса (хотя я не уверен что это он… осциллограф не работает) диод засветился а потребления с 20 милиампер упало до 0 хотя это наверно потому что у меня в тестере больше чувствительности не хватает… дак вот собственно стал подключать к остальным парам светодиоды и также тестер показывал 0! а яркость первого ничуть не менялась от подключения полного набора светодиодов… но картина другая если поменять полярность светодиодов… тогда при подключении больше одного светодиода начинали заметно угасать… то есть как по классике… если кому интересно могу выложить схему и видео с экспериментом… Нет мне не нужно. Я твой генератор отработал в деталях, под уже технологию. Я их сделал уже десять штук самых разных. Упростил до некуда. В итоге получилось, алюминиевая шинка и на ней два транзистора, навесной монтаж и вход и выход. Все генераторы запускаются почти от нуля вольт и выше. То есть практически лежит куча генераторов, вставил их в разъемы и исследуй. Теперь делаю стенд, для иследования,уже индуктивностей и конденсаторов. Конденсаторы тоже бывают разные, и похоже не каждый конденсатор подходит к этому делу. Сразу видно что конденсатор, на пятьдесят вольт и качер на пятнадцать вольт, не совместимы. Конденсатор должен иметь запас, хотя бы раз в десять, чем мы видим на осцилографе. Потом полярные и неполярные тоже работают по разному, пока не понятно как. Емкость конденсатора тоже похоже от чего то зависит, я ставил на максимальную емкость, но это похоже не выход. Ну и наконец диоды, тоже отдельная песня. Я почему качеры настругал десяток. Качеры “свергетеродинные диоды”, а простые диоды тоже своеобразно работают.
17 Потом я попробовал индуктивность, дроссель с зазором. Причем зазор – резиновая пленка от презерватива. На 50 герц он работал замечательно. А тут, никаких дрочек. Вибрации никакой. Хотя витки на кольцевом феррите не затянутые изолентой пищат замечательно причем в тон контрольного радиоприемника. Ну вобщем надо репу чесать. Никто не поможет. Походил, подумал. Все таки придется вернуться к клещам на датчике холла, для измерения тока в таких цепях. Они правда под пятерку тысяч, но меряют эти “невидимые” токи довольно хорошо. Я тогда помоему, один из первых замерил токи в катушках качера. Они оказались гораздо больше, чем показывали все эти Ц- шки и тестеры. Я тогда и сказал, кто сделал качер – это уже “сверхединичный прибор”. А вот снимать с него нормальные токи получается только сейчас. И то не совсем.хотя эти лампочки и светодиоды, кто как не изгалялся, уже все делали. А этот ток оказывается снимается на конденсаторы и аккумуляторы и только потом превращаются в обычные токи. Причем именно в постоянный ток. Да и конденсаторы заряжаются далеко не сразу. Им надо определенное время. Для зарядки. И определенное время для разрядки. Правда это школьная задача. В емкость, “бассейн” вливается десять труб, а выливается из одной трубы. Надо расчитать оптимальный расход воды. Так и электрический ток, можно сказать воды. «Вода» течет не только по «трубам» проводам, но и обменивается энергией (трансформацией) через окружающее индуктивности пространство. Не освоил вставлять фотки извините мучаюсь. Тем более у меня люникс. Дело не в операционной системе, дело в том, что нажимая кнопку [ i m g ], мы тем самым должны вставить адрес объекта (картинки) в Интернете, который был бы постоянно доступен. Любой сервер хранения картинок удовлетворяет этой цели. Адрес вида file:///home говорит о том, что объект находится на локальном диске автора публикации. Понятно, что никто другой ничего не увидит… а чем плохая модель? к тому же она не касается токов смещения только. Это картинка того как проявляется вещество. Только вот не очень я согласен с тем материя непрерывно телепортируется на шаг сетки эфира, а не движется цельным куском. Автор счи-
18 тает что материя постоянно создается и уничтожается каждый миг. И состоит в каждый новый момент из новых частиц эфира. Хотя я и сам так раньше думал. Ведь в этом случае выходит поле и намерение первичны, материя вторична. От таких дум депресняк находит… Начинаешь понимать насколько ничтожны попытки понять мир, и его законы. Таки человек это кучка микробов в одном чехле, блять… Вот видишь, как плохо без признания иных ортогоналей. Не исчезает и появляется из ниоткуда, а переныривает из одной ортогонали в другую. Надо полагать, всегда соседнюю. Что у нас в точности и воспринимается пульсацией массы. Конечно, всегда с максимально возможной скоростью. Память же о намерениях – это и есть инерция массы вовлечённого в этот бег. И плоский срез картины – как волны ряби, бегущие по поверхности раздела воды и воздуха. Только без трения, понятно. Да на уровне нано как раз с трением хреновастенько и будет. Мы всё время ищем поперечную волну – их ведь не бывает без границы сред. Но именно граница этих сред существует в каждой точке пространства. С пороговой проницаемостью, похоже. Иначе, с продольными волнами, возникает полная жопа: пульсации зарядов – в образе продольно движущихся зарядовых градиентов. А где место тому же Э/М полю, которое мы ловим сотовыми? Потому поперечные волны нам верный друг. Проблема только в одном – найти эту сраную границу раздела сред. Без которой всегда прав суровый Ацюковский. Он же в вихри свои полез не от хорошей жизни. А потому, что Левашова не читал, и сам не думал на эти темы. Хоть и общался с эфиродинамиком Ивановым на постоянной основе. Эвклидова трехмерность вокруг нас сыграла плохую шутку. Ну и что? Уделали же её и Лобачевский, и Гаусс, и Риман. Но они разве не ввели контробандно новую ортогональ – кривизну пространства в каждой точке? Ввели, засранцы! Но не объявили об этом честно и прямо. Так что никакие мы не революционеры. Просто называем вещи привычными именами. И не стыдимся.
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в cookie-файлах может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
pEGL1-D232A | ATCC
Продукт предоставляется «КАК ЕСТЬ», и на жизнеспособность продуктов ATCC ® дается гарантия в течение 30 дней с даты отгрузки, при условии, что покупатель хранил продукт и обращался с ним в соответствии с информацией, содержащейся в информационном листе продукта, веб-сайт и сертификат анализа. Для живых культур ATCC перечисляет состав сред и реагенты, которые оказались эффективными для продукта.Хотя другие неуказанные среды и реагенты также могут давать удовлетворительные результаты, изменение в протоколах, рекомендованных АТСС и / или депонентом, может повлиять на извлечение, рост и / или функцию продукта. Если используется альтернативный состав среды или реагент, гарантия ATCC на жизнеспособность больше не действует. За исключением случаев, прямо изложенных в настоящем документе, никакие другие гарантии любого рода, явные или подразумеваемые, не предоставляются, включая, помимо прочего, любые подразумеваемые гарантии товарной пригодности, пригодности для конкретной цели, производства в соответствии со стандартами cGMP, типичности, безопасности, точности. , и / или ненарушение прав.
Этот продукт предназначен только для лабораторных исследований. Он не предназначен для каких-либо терапевтических целей на животных или людях, для употребления в пищу людьми или животными или для каких-либо диагностических целей. Любое предлагаемое коммерческое использование запрещено без лицензии ATCC.
Хотя ATCC прилагает разумные усилия для включения точной и актуальной информации в этот лист продукта, ATCC не дает никаких гарантий или заявлений относительно ее точности.Цитаты из научной литературы и патентов приводятся исключительно в информационных целях. ATCC не гарантирует, что такая информация была подтверждена как точная или полная, и заказчик несет исключительную ответственность за подтверждение точности и полноты любой такой информации.
Этот продукт отправляется при условии, что клиент несет ответственность и принимает на себя все риски и ответственность в связи с получением, обращением, хранением, утилизацией и использованием продукта ATCC, включая, помимо прочего, принятие всех соответствующих мер безопасности и мер предосторожности при обращении для минимизации риск для здоровья или окружающей среды.В качестве условия получения материала заказчик соглашается с тем, что любые действия, предпринимаемые с продуктом ATCC и любыми дочерними продуктами или модификациями, будут осуществляться в соответствии со всеми применимыми законами, нормативными актами и руководящими принципами. Этот продукт предоставляется «КАК ЕСТЬ» без каких-либо заверений или гарантий, за исключением случаев, прямо указанных в настоящем документе, и ни при каких обстоятельствах ATCC, ее материнские компании, дочерние компании, директора, должностные лица, агенты, сотрудники, правопреемники, правопреемники и аффилированные лица не несут ответственности за косвенные , особые, случайные или косвенные убытки любого рода в связи с использованием продукта покупателем или возникшие в результате его использования.Несмотря на то, что прилагаются разумные усилия для обеспечения подлинности и надежности материалов, находящихся на хранении, ATCC не несет ответственности за ущерб, возникший в результате неправильной идентификации или искажения таких материалов.
Дополнительные сведения об использовании этого продукта см. В соглашении о передаче материалов (MTA). MTA доступен на сайте www.atcc.org.
1p8s – Протеопедия, жизнь в 3D
Структурные особенности
Эволюционное сохранение
Проверка, как определено ConSurfDB.Вы можете прочитать объяснение метода и полные данные, доступные на ConSurf.
Резюме публикации из PubMed
Аргиназа – это двухъядерный металлофермент марганца, который гидролизует l-аргинин с образованием l-орнитина и мочевины. Трехмерные структуры D128E, D128N, D232A, D232C, D234E, h201N и h201E аргиназ I были определены методами рентгеновской кристаллографии для выяснения роли лигандов металлов первой оболочки в стабильности и каталитической активности фермент.Эта работа представляет собой первое рассечение биядерного марганцевого кластера на основе структуры с использованием сайт-направленного мутагенеза и рентгеновской кристаллографии. Замена металлических лигандов ставит под угрозу каталитическую активность фермента либо за счет потери, либо за счет разрушения металлического кластера или нуклеофильного гидроксид-иона, соединяющего металл. Однако замена металлических лигандов или уменьшение занятости Mn (2 +) (A) или Mn (2 +) (B) не ставит под угрозу сродство фермента к субстрату, что отражается в K (M), которое остается относительно неизменным для эта серия вариантов аргиназы.Это предполагает наличие неметаллического сайта связывания субстрата l-аргинина в прекаталитическом комплексе Михаэлиса, как было предложено на основе анализа структуры нативного фермента (Kanyo, ZF, Scolnick, LR, Ash, DE и Christianson, DW (1996) Nature 383, 554-557).
Структурное и функциональное значение лигандов металлов первой оболочки в биядерном марганцевом кластере аргиназы I., Cama E, Emig FA, Ash DE, Christianson DW Biochemistry. 1 июля 2003 г .; 42 (25): 7748-58. PMID: 12820884 [1]
Из MEDLINE® / PubMed®, базы данных U.С. Национальная медицинская библиотека.
См. Также
Список литературы
- ↑ Cama E, Emig FA, Ash DE, Christianson DW. Структурное и функциональное значение лигандов металлов первой оболочки в биядерном марганцевом кластере аргиназы I. Биохимия. 1 июля 2003 г .; 42 (25): 7748-58. PMID: 12820884 doi: http: //dx.doi.org/10.1021/bi030074y
Ключевые остатки, ответственные за повышение каталитической эффективности липазы Lip от Thermomyces lanuginosus, выявленные с помощью стратегии комплементарной белковой инженерии.
Journal of Biotechnology 188 (2014) 29–35Списки содержания доступны на ScienceDirect
Домашняя страница журнала Journal of Biotechnology: www.elsevier.com/locate/jbiotec
Ключевые остатки, ответственные за повышение каталитической эффективности липазы Thermomyces lanuginosus Lip, выявленные с помощью стратегия комплементарной белковой инженерии Сяо-Цзюнь Ли a, b, Рен-Чао Чжэн a, b, Хун-Е Ма a, b, Цзянь-Фэн Хуан a, b, Ю-Го Чжэн a, b, ∗ ab
Институт Биоинженерия, Технологический университет Чжэцзян, Ханчжоу 310014, Китайский инженерный исследовательский центр биоконверсии и биочистки Министерства образования, Технологический университет Чжэцзян, Ханчжоу 310014, Китай
article
info
История статьи: Поступила 17 мая 2014 г. Поступила в редакции Форма 25 июля 2014 г. Принята 1 августа 2014 г. Доступна онлайн 10 августа 2014 г. Ключевые слова: Липаза Белковая инженерия Полимеразная цепная реакция, подверженная ошибкам Прегабалин Каталитическая эффективность
аннотация Вариант Lip-T (S88T / A99N / V116D) липазы Lip из Thermomyces lanuginosus оказался потенциальным биокатализатором для кинетического разделения этилового эфира 2-карбоксиэтил-3-циано-5-метилгексановой кислоты (CNDE) на производят ценный хиральный промежуточный продукт прегабалина.В этом исследовании был проведен случайный, сайт-направленный и сайт-насыщенный мутагенез для дальнейшего повышения активности Lip-T, и были выявлены ключевые остатки, ответственные за каталитическую эффективность. Мутант S63L / D232A с улучшенной активностью по отношению к CNDE был получен после скрининга приблизительно 2500 клонов из библиотек случайных мутантов. Сайт-направленный мутагенез в сайтах 63 и 232 продемонстрировал, что одноточечные мутанты S63L и D232A проявляют противоположный эффект на активность. S63L продемонстрировал значительное улучшение активности, тогда как D232A проявил небольшой ингибирующий эффект.Затем с помощью мутагенеза с насыщением сайтов был получен мутант S63M с каталитической эффективностью в 4,5 раза выше, чем у Lip-T. Были проанализированы структурные изменения, возникающие в результате мутаций, и обсуждены механизмы, ответственные за усиление активности. Более того, сконструированная липаза катализирует энантиоселективный гидролиз CNDE при очень высокой загрузке субстрата (765 г / л). Поскольку использовалось только 5% (мас. / Об.) Покоящихся клеток, биопроцесс намного более рентабелен, чем процесс Pfizer с использованием 8% (мас. / Об.) Коммерчески доступной липазы Lipolase®.Эти результаты предоставляют не только новое понимание взаимосвязи структура-функция липазы, но и новый надежный биокатализатор для производства прегабалина. © 2014 Elsevier B.V. Все права защищены.
1. Введение Липазы (EC 3.1.1.3) представляют собой ферменты, катализирующие гидролиз и синтез триацилглицерина и других сложных эфиров жирных кислот (Jaeger et al., 1999; Schmid and Verger, 1998). Он является членом семейства складчатых ␣ / -гидролаз (Nardini and Dijkstra, 1999), активный центр которого состоит из каталитической триады Ser-His-Asp / Glu, а остаток Ser находится в консенсусной последовательности Gly-X -Ser-X-Gly (Брэди и др., 1990; Jaeger et al., 1999). Благодаря таким свойствам, как высокая стабильность, широкая специфичность к субстрату, независимость от кофакторов и высокая специфичность (хемо-, регио- и энантиоселективность), липазы играют важную роль в фармацевтике, агрохимии, продуктах питания
∗ Автор для корреспонденции: Институт биоинженерии, Технологический университет Чжэцзян, Ханчжоу 310014, Китай. Тел .: +86571 88320630; факс: +86 571 88320630. Адрес электронной почты: [электронная почта защищена] (Y.-G. Zheng). http: //dx.doi.org / 10.1016 / j.jbiotec.2014.08.004 0168-1656 / © 2014 Elsevier B.V.Все права защищены.
и индустрии моющих средств (Bassegoda et al., 2012; Gupta et al., 2004; Jaeger and Eggert, 2002). По сравнению с липазами растений и животных микробные липазы имеют большее промышленное значение и широко используются для катализирования разнообразных реакций биоконверсии из-за низкой стоимости производства, более высокой стабильности и большей доступности, чем другие источники (Gupta et al., 2004). Однако только несколько липаз доступны для промышленного применения из-за сложности разработки биокатализаторов с высокой производительностью процесса и создания рентабельного расширения масштабов.Разработка новых липаз с высокой производительностью процесса имеет решающее значение для расширения их промышленного применения. Белковая инженерия стала мощным средством для изменения характеристик ферментов (Bornscheuer et al., 2012), хотя некоторые методы, такие как иммобилизация и химическая модификация белков, могут в определенной степени улучшить свойства ферментов (Marciello et al., 2012; Rodrigues и др., 2013). (S) -3- (Аминометил) -5-метилгексановая кислота (Прегабалин) представляет собой липофильное производное 4-аминомасляной кислоты (ГАМК), ключевого ингибирующего нейромедиатора в центральной нервной системе млекопитающих
30
X.-J. Ли и др. / Journal of Biotechnology 188 (2014) 29–35
Система(Mukherjee and Martinez, 2011; Zheng et al., 2013). Прегабалин превратился в лекарство-блокбастер для лечения ряда заболеваний центральной нервной системы (Silverman, 2008). Поскольку (S) -прегабалин значительно более эффективен, чем рацемат и (R) -энантиомер (Belliotti et al., 2005), было разработано несколько химических и хемоэнзиматических способов получения оптически чистого (S) -прегабалина (Chen et al. al., 2011). Наиболее успешная масштабируемая хемоферментная стратегия для (S) -прегабалина включала катализируемое липазой расщепление этилового эфира 2-карбоксиэтил-3-циано-5-метилгексановой кислоты (CNDE) до (3S) -2-карбоксиэтил-3-циано-5-метилгексановой кислоты. (Martinez et al., 2008). До сих пор, за исключением Lipolase®, коммерчески доступной липазы Thermomyces lanuginosus (TLL), поставляемой Novozymes (Fernandez-Lafuente, 2010), в промышленном производстве прегабалина не использовался другой надежный биокатализатор. В нашем предыдущем исследовании еще одна термостабильная липаза Lip от T.lanuginosus был клонирован и гетерологично экспрессирован в Escherichia coli (Li et al., 2014). Хотя Lip проявлял высокую энантиоселективность по отношению к CNDE (E> 200), низкая активность ограничивала его промышленное применение. После мутагенеза с насыщением сайтов в оксианионном отверстии и шарнирной области крышки тройной вариант Lip-T, S88T / A99N / V116D показал 60-кратное улучшение специфической активности для CNDE по сравнению с Lip дикого типа. Чтобы разработать новый биокатализатор с высокой производительностью процесса, совместимый с промышленными предпосылками, в этой работе мы попытались дополнительно повысить его активность в отношении CNDE Lip-T с помощью стратегии комплементарной белковой инженерии, включая подверженную ошибкам ПЦР, сайт-направленный мутагенез. и мутагенез с насыщением сайтов.Были выявлены ключевые остатки, ответственные за повышение каталитической эффективности Lip, и был получен новый надежный биокатализатор для кинетического разрешения CNDE. 2. Материалы и методы 2.1. Ферменты, химические вещества и реагенты ДНК-полимераза Taq и ДНК-полимераза PrimeSTAR HS для подверженной ошибкам ПЦР и сайт-направленного мутагенеза были получены от Takara (Далянь, Китай). Эндонуклеазы рестрикции и ДНК-лигаза Т4 были приобретены в Fermentas (Шэньчжэнь, Китай). Набор для экстракции ДНК из геля и набор для экстракции плазмидной ДНК были от Axygen (Ханчжоу, Китай) и использовались в соответствии со спецификациями производителя.Изопропил--d-тиогалактопиранозид (IPTG) и канамицин были приобретены у Sigma. CNDE был любезно предоставлен Zhejiang Apeloa Medical Technology Co., Ltd. (Цзиньхуа, Китай). Все остальные химические вещества были аналитической чистоты. 2.2. Бактериальные штаммы и плазмиды E. coli BL21 (DE3) и pET-28b (+) использовали в качестве штамма-хозяина и вектора для экспериментов по экспрессии. Плазмида pET28Lip-T, несущая ген, кодирующий тройной мутированный вариант S88T / A99N / V116D липазы Lip из T. lanuginosus, была сконструирована и сконструирована, как описано ранее (Li et al., 2014). 2.3. ПЦР, подверженная ошибкам, и конструирование мутантной библиотеки ПЦР, подверженная ошибкам, была проведена с использованием плазмиды pET28-Lip-T в качестве матрицы, а ген липазы был амплифицирован со стандартным праймером промотора Т7 и праймером терминатора Т7 (таблица дополнительных материалов S1), которые отжигаются за пределами сайты клонирования (NcoI / HindIII). Реакционная смесь для ПЦР, подверженная ошибкам, содержала 1 × буфер Taq без Mg2 +, 0,2 мкМ каждого праймера, 7 мМ MgCl2, 0,2 мМ MnCl2, 0,2 мМ dGTP, 0,2 мМ dATP, 1,0 мМ dCTP, 1,0 мМ dTTP, 50 нг матричной ДНК. и 5 ед. полимеразы Taq в реакционном объеме 100 мкл.Условия ПЦР были следующими: горячий старт при 94 ◦ C в течение 5 минут, затем 30 циклов при 94 ◦ C в течение 30 с, 55 ◦ C в течение 30 с и 72 ◦ C
в течение 60 с, с заключительным этапом продления. при 72 ◦ C в течение 10 мин. Продукты ПЦР проверяли электрофорезом в агарозном геле, а затем очищали с использованием набора для экстракции ДНК из геля. Впоследствии амплифицированный мутированный продукт ПЦР расщепляли NcoI и HindIII и лигировали в плазмиду pET-28b (+), которая ранее была также аналогичным образом расщеплена теми же ферментами.Лигирование выполняли с ДНК-лигазой Т4 при 16 ° C в течение ночи, и полученную смесь для лигирования использовали для трансформации компетентных клеток штамма E. coli BL21 (DE3) и высевали на агаровую среду LB с добавлением 50 мкг / мл канамицина. с использованием стандартных процедур молекулярной биологии, как описано Sambrook and Russell (2001). 2.4. Метод скрининга мутантов с улучшенной активностью Первоначальный скрининг мутантов с улучшенной активностью из случайной библиотеки мутантов проводили в 96-луночных планшетах с использованием CNDE в качестве субстрата в высокопроизводительном скрининге на основе pH, как описано ранее (Li et al., 2014; Moris-Varas et al., 1999). Колонии отбирали в отдельные лунки 96-луночных планшетов, содержащих среду LB с добавлением 50 мкг / мл канамицина. 96-луночные планшеты инкубировали при 37 ° C до тех пор, пока значение OD600 не достигло примерно 0,6, с последующей индукцией 0,1 мМ IPTG и дальнейшим ростом при 28 ° C в течение 10 часов. Клетки собирали центрифугированием (4000 × g, 10 мин) и ресуспендировали в калий-фосфатном буфере (10 мМ, pH 7,2). Аликвоты (20 мкл) супернатанта переносили в отдельные лунки новых микротитровальных планшетов, содержащих 180 мкл раствора субстрата (10 мМ калий-фосфатный буфер, pH 7.2, 0,01% бромтимоловый синий, 100 мМ CNDE) и инкубирование при 30 ◦ C. Клеточный супернатант E. coli BL21 (DE3) / pET28-Lip-T использовали в качестве контроля при скрининге библиотеки случайных мутантов. Скорость изменения цвета реакционной системы с синего на желтый использовали в качестве показателя активности. Для дальнейшего подтверждения отбирали только мутанты с более высоким индексом активности по сравнению с контрольной клеткой. Отобранные мутанты повторно выращивали в масштабе 100 мл и анализировали активность в трех повторностях. Отобранные мутанты секвенировали для подтверждения внесенных мутаций.2.5. Конструирование одноточечных мутантов и библиотек мутагенеза насыщения. Чтобы определить, необходимы ли замены в S63L / D232A для улучшения активности, одноточечные мутанты S63L и D232A индивидуально вводили в Lip-T посредством сайт-направленного мутагенеза. Сайт-направленный мутагенез выполняли с использованием праймеров S63L-F / R и D232A-F / R (таблица дополнительных материалов S1) для определения замены одной аминокислоты в положении 63 и 232 лейцином и аланином, соответственно.Впоследствии для дальнейшего исследования эффектов замен, замененных другими 19 аминокислотами в сайтах S63 и D232, библиотеки мутагенеза с насыщением были созданы в сайтах S63 и D232 с использованием соответствующих прямых и обратных праймеров, перечисленных в таблице дополнительных материалов S1, соответственно. Сайт-направленный мутагенез и сайт-насыщенный мутагенез проводили с использованием протокола, основанного на ПЦР, как описано Chronopoulou и Labrou (2011). Реакции ПЦР проводили с ДНК-полимеразой PrimeSTAR HS с использованием соответствующих праймеров и плазмиды pET28-Lip-T в качестве ДНК-матрицы.Используемая программа ПЦР составляла 2 минуты при 98 ◦ C, затем 25 циклов по 10 секунд при 98 ◦ C, 10 секунд при 60 ◦ C и 6 минут при 72 ◦ C и завершалась 10 минутами при 72 ◦ C. После подтверждения агарозой. После гель-электрофореза 20 мкл продуктов амплификации ПЦР обрабатывали 1 мкл DpnI для удаления матричной плазмидной ДНК, и реакционные смеси трансформировали в компетентные клетки E. coli BL21 (DE3), а затем высевали на агаровую среду LB с добавлением 50 мкл Г мл − 1 канамицина. Первый раунд скрининга мутантов (примерно 500 клонов
X.-J. Ли и др. / Journal of Biotechnology 188 (2014) 29–35
31
на сайт) с улучшенной активностью из библиотек мутагенеза с насыщением сайтов проводили в 96-луночных планшетах, как описано выше. Клеточный супернатант E. coli BL21 (DE3) / pET28-S63L и E. coli BL21 (DE3) / pET28-Lip-T использовали в качестве контроля при скрининге библиотек мутагенеза насыщения, созданных в сайтах S63 и D232, соответственно. 2.6. Экспрессия и очистка липазы дикого типа и мутантной липазы. Отобранные мутанты и Lip-T были дополнительно экспрессированы в E.coli BL21 (DE3). Отобранные трансформанты инокулировали в среду LB в течение ночи при 37 ° C в качестве посевной культуры. Однопроцентные семена переносили в свежую среду LB. Клетки выращивали при 37 ◦ C с канамицином до тех пор, пока оптическая плотность при 600 нм не достигла примерно 0,6. IPTG добавляли до конечной концентрации 0,1 мМ, чтобы вызвать экспрессию, и клетки культивировали при 28 ◦ C в течение следующих 10 часов. Клетки собирали центрифугированием для очистки и анализа активности. Экспрессированные липазы дикого типа и мутантные липазы очищали до гомогенности из бесклеточного экстракта тремя дополнительными методами разделения, включая осаждение сульфатом аммония, хроматографию гидрофобного взаимодействия (колонка с фенилсефарозой 6 FF, 1.6 × 20 см, GE) и ионообменной хроматографии (колонка DEAE-Sepharose FF, 1,6 × 20 см, GE) (Li et al., 2014). Чистоту белка проверяли электрофорезом в 12% додецилсульфат натрия в полиакриламидном геле (SDS-PAGE). Концентрации белка определяли по методу Брэдфорда (1976) с использованием бычьего сывороточного альбумина в качестве стандарта. 2.7. Анализ активности и аналитические методы. Активность липазы определяли путем мониторинга гидролиза CNDE, а энантиомерные составы остаточного эфира и соответствующей кислоты в реакционной системе определяли с помощью газовой хроматографии GC-14C (Shimadzu, Япония), оснащенной детектором FID и хиральная капиллярная колонка Astec CHIRALDEXTM G-TA (30 м × 0.25 мм, толщина пленки 0,25 мкм) с использованием гелия в качестве газа-носителя (Li et al., 2014; Zheng et al., 2013). За одну единицу активности (U) принимали количество фермента, высвобождающего 1 мкмоль 2-карбоксиэтил-3-циано-5-метилгексановой кислоты в минуту. Чтобы определить диастереоселективность вариантов по отношению к CNDE, образцы были дериватизированы триметилсилилдиазометаном (TMSCh3 N2) и метанолом, и диастереомерный избыток (de) был рассчитан в соответствии с методом, описанным ранее (Zheng et al., 2013). 2.8. Кинетический анализ дикого типа и вариантов. Константа Михаэлиса-Ментен (Km) и максимальная скорость реакции (Vmax) были рассчитаны для вариантов и Lip-T из стандартных графиков Микелиса-Ментен. Кинетические параметры (Km и Vmax) определяли при 40 ◦ C и pH 7,5 (100 мМ трис-HCl буфер) при варьировании концентрации субстрата (2–100 мМ) с использованием 5% ацетонитрила. 2.9. Кинетическое разрешение CNDE с помощью сконструированной липазы Для оценки каталитической эффективности сконструированной липазы кинетическое разрешение CNDE на (3S) -2-карбоксиэтил-3-циано5-метилгексановую кислоту было выполнено с использованием S63M или Lip-T для цельных клеток в качестве биокатализатор.Реакционная смесь (1,0 л) содержит 5% клеток E. coli (мас. / Об., 50 г влажных клеток / л), 150 мМ ацетата кальция и 1–3 М CNDE. Полученный раствор инкубировали при 30 ◦ C и перемешивали при 400 об / мин, а pH реакции поддерживали на уровне 7,0 с помощью автотитратора с использованием 5 M раствора NaOH. Образцы были
Рис. 1. Схематическое изображение направленной эволюции липазы Lip с повышенной активностью в отношении CNDE. Было показано сочетание различных методов мутагенеза (подверженная ошибкам ПЦР, сайт-направленный мутагенез и сайт-насыщенный мутагенез) для эволюции липазы Lip.Удельная активность очищенного Lip-T (2,35 Ед / мг) и Lip дикого типа (0,04 Ед / мг) установлена как 1, соответственно. Указаны аминокислотные замены, обнаруженные в этих вариантах.
периодически извлекают из реакционной смеси для определения степени превращения и энантиомерного избытка (ее) с помощью хирального ГХ-анализа. 2.10. Молекулярное моделирование и анализ. Трехмерная модель гомологии Lip-T и мутантов была построена на основе известной структуры липазы T. lanuginosus (PDB 1GT6) (Yapoudjian et al., 2002) с помощью программы MODELER, модуля в Discovery Studio 2.1 (Accelrys Software, Сан-Диего, США). Сгенерированная модель затем была подвергнута оптимизации молекулярной механики с использованием силового поля CHARMM27 и проверена с помощью программы Procheck (Laskowski et al., 1993). Визуализация и графика выполнены с помощью программы PyMOL (http://www.pymol.org). 3. Результаты 3.1. Скрининг мутантной библиотеки на варианты с улучшенной активностью. ПЦР, подверженная ошибкам, была использована для создания случайной мутантной библиотеки Lip-T со средней скоростью мутаций в 2–3 замены оснований на ген липазы, что соответствует максимум трем аминокислотным заменам на липазу. белок.Полученную библиотеку подвергли скринингу на наличие мутантов с улучшенной активностью в формате 96-луночного планшета. Для дальнейшего подтверждения были отобраны только мутанты с более высоким индексом активности по сравнению с Lip-T. Мутант-кандидат, названный 3D5, со значительно улучшенной активностью по отношению к CNDE, был отобран среди примерно 2500 клонов. 3D5 был идентифицирован и дополнительно охарактеризован с использованием очищенного фермента. Секвенирование ДНК аллеля Lip-T из плазмиды в 3D5 показало, что он имеет две аминокислотные замены (S63L и D232A) по сравнению с Lip-T (рис.1). Мутант был очищен, и удельная активность S63L / D232A составила 4,78 ± 0,21 Ед / мг. Чтобы определить, необходимы ли обе замены в S63L / D232A для улучшения активности, был введен сайт-направленный мутагенез для конструирования одноточечных мутантов S63L и D232A, соответственно. Варианты очищали и подтверждали с помощью SDSPAGE (дата не показана). Интересно, что мутанты S63L и D232A показали противоположный эффект на активность. Замена S63L показала значительный эффект улучшения активности (7.21 ± 0,26 Ед / мг), тогда как замена D232A оказывала небольшое ингибирующее действие на активность (1,59 ± 0,15 Ед / мг) (рис. 1). Результаты показали, что оба сайта важны для каталитической способности фермента.
32
X.-J. Ли и др. / Journal of Biotechnology 188 (2014) 29–35
Рис. 2. Анализ диастереоселективности Lip-T и мутантов по CNDE. ГХ-хроматограммы ограниченного химического гидролиза CNDE 100 мМ NaOH (A), ферментативного гидролиза CNDE с помощью Lip-T (B), мутанта S63L / D232A (C), S63L (D) и S63M (E), соответственно.Образцы были испытаны после дериватизации ТМСЧ3 N2 и метанолом.
Для повышения эффективности направленной эволюции библиотеки мутагенеза с насыщением в сайтах S63 и D232 были созданы и подвергнуты скринингу, соответственно. Используя S63L в качестве контроля, был получен и идентифицирован мутант S63M с повышенной каталитической активностью в отношении CNDE (8,68 ± 0,23 Ед / мг) по сравнению с S63L. Однако не было обнаружено дальнейшего улучшения при проведении мутагенеза с насыщением сайтов в положении 232. Результат показал, что исходный аспартат уже мог быть оптимальным в положении 232.Когда липазы дикого типа и все мутантные липазы экспрессировались в хозяине E. coli BL21 (DE3), было обнаружено, что не было разницы в уровнях экспрессии и молекулярных массах белков (данные не показаны). 3.2. Влияние мутации на энантиоселективность. Оптическая чистота стала решающим прилагательным свойством лекарственных соединений в фармацевтической промышленности в последние десятилетия. Следовательно, влияние на энантиоселективность, вызванное мутациями, следует учитывать при разработке ферментов. В данной работе замены практически не повлияли на энантиоселективность и диастереоселективность фермента по отношению к CNDE (рис.2). Величина ее и де-ценность продукта мутантов составляла 98% и 98%, такие же, как у фермента дикого типа, что демонстрирует, что замена не влияет на стереоспецифичность фермента.
3.3. Анализ кинетических параметров Lip-T и вариантов Кинетические параметры (Km, Vmax, kcat и kcat / Km) Lip-T и вариантов по отношению к CNDE были определены и суммированы в таблице 1. Наблюдается небольшое влияние на Km для всех мутантов. с улучшенной активностью, в то время как значительное увеличение kcat, что внесло основной вклад в повышение каталитической эффективности.По сравнению с Lip-T, значения Km всех мутантов, кроме D232A, немного снизились, а значения Vmax мутантов S63L / D232A, S63L и S63M увеличились до некоторой степени. Каталитические константы (kcat) мутантов S63L / D232A, S63L и S63M увеличились в 2,1, 3,2 и 3,7 раза соответственно, а у мутанта D232A снизились до 70%. Каталитическая эффективность мутантов (S63L / D232A, S63L и S63M) для CNDE значительно увеличилась, на что указывают более высокие значения kcat / Km по сравнению с липазой дикого типа.Самый активный мутант, S63M, продемонстрировал 4,5-кратное улучшение каталитической эффективности по сравнению с Lip-T. Между тем, каталитическая эффективность мутантов D232A по отношению к CNDE значительно снизилась, на что указывают более низкие значения kcat / Km, чем у Lip-T. С другой стороны, параметры кинетики мутантов оценивали путем перекрестного сравнения с TLL, очищенным от коммерческой липолазы®, как описано ранее (Peters et al., 1998). Стоит отметить, что каталитическая эффективность мутанта S63M составляла около 4.В 4 раза больше, чем у TLL.
X.-J. Ли и др. / Journal of Biotechnology 188 (2014) 29–35
33
Таблица 1 Кинетические параметры очищенных TLL, Lip и мутантов. Ферменты
км (мМ)
Lip TLLa Lip-T S63L / D232A S63L D232A S63M
36,6 19,7 18,2 15,1 15,1 18,4 15,1
a
± ± 0,2 ± 0,3 ± ± ±
9000 0,6 0,2 0,8 0,2 0,2Vmax (мкмоль мг − 1 мин − 1) 0,074 2,80 2,71 5,62 8,61 1,89 10,01
± ± ± ± ± ± ±
0.005 0,17 0,21 0,29 0,31 0,19 0,30
ккат (мин − 1)
ккат / км (мин − 1 мМ − 1)
Каталожный номер
2,22 88,85 81,3 168,6 258,3 56,7 300,3
0,06 4,51 4,46 11,16 9,109 3,08
Li et al. (2014) Ли и др. (2014) Это исследование Это исследование Это исследование Это исследование Это исследование
TLL, липаза Thermomyces lanuginosus, очищенная от коммерческой Lipolase®.
Рис. 4. Кривые прогресса кинетического разрешения CNDE с использованием мутантных S63M и цельных клеток Lip-T в качестве биокатализатора, соответственно.Реакционная смесь содержит 5% остальных клеток (вес / объем, 50 г сырого веса клеток / л), 150 мМ ацетата кальция и 1–3 М CNDE. Полученный раствор инкубировали при 30 ◦ C и перемешивали при 400 об / мин, а pH реакции поддерживали на уровне 7,0 с помощью автотитратора с использованием 5 M раствора NaOH. Рис. 3. Модель структуры Lip-T и положение сайтов мутагенеза в LipT. Остатки каталитической триады активного центра (Ser151, Asp206 и His263) помечены зеленым цветом с моделями «палочки». Крышка с ␣-спиралью (Val91-Asn97) и две петли вокруг крышки (Gly87-Ser90 и Leu98-Asp101) отмечены пурпурным цветом.Положение сайтов мутагенеза показано красным цветом с моделями палочек. (Для интерпретации ссылки на цвет в легенде этого рисунка читатель может обратиться к веб-версии этой статьи.)
3.4. Структурные наблюдения Чтобы получить некоторое представление об улучшении активности ферментов вариантов S63L / D232A, S63L и S63M, общая трехмерная структура каждого варианта, очень похожая на структуру Lip дикого типа, была построена методами молекулярного моделирования. Относительные положения замещенных аминокислотных остатков были расположены в смоделированной структуре.Остаток S63 лежит на петле 2, которая соединена с цепью 1 и цепью 2. Мутация D232A затрагивает остаток в петле между цепью 8 и спиралью 6. Все замены были локализованы на поверхности и оказались в областях, далеких от связывания субстрата и активного центра (рис. 3). Следовательно, было трудно предсказать эти мутации с помощью рациональных подходов к дизайну. 3.5. Кинетическое разрешение CNDE с высокой загрузкой субстрата. Чтобы проверить каталитическую эффективность мутанта S63M в энантиоселективном гидролизе CNDE, кинетическое разрешение было выполнено с использованием целых клеток в качестве биокатализатора.На рис. 4 показаны временные профили синтеза (3S) -2-карбоксиэтил-3-циано-5-метилгексановой кислоты с помощью Lip-T и S63M. На начальном этапе реакции количество продуктов резко увеличивается. (3S) -2-карбоксиэтил-3-циано-5-метилгексановая кислота была получена в 463 мМ с загрузкой субстрата 1 М (255 г / л) через 7 ч, 936 мМ с 2 М (510 г / л) через
16 ч и 1356 мМ с 3 М (765 г / л) через 24 ч соответственно. Напротив, только 451 мМ продукт был образован с 1,0 M CNDE после 24-часовой биоконверсии целыми клетками Lip-T.Каталитическая эффективность S63M была в 3,31 раза выше, чем у Lip-T при такой же загрузке субстрата (1 M). Кроме того, катализируемое S63M разделение CNDE происходило гладко при загрузке субстрата до 3,0 М. Значение ее продукта для S63M было таким же, как у фермента дикого типа (98%), что дополнительно продемонстрировало, что замена не влияет на стереоспецифичность. фермента. Эти результаты показали, что мутант S63M является надежным биокатализатором для кинетического разрешения CNDE. 4. Обсуждение В последние годы ферменты все чаще применяются в промышленных условиях в качестве дешевой и экологически чистой альтернативы химическим катализаторам (Quin and Schmidt-Dannert, 2011).Однако в большинстве случаев ферменты дикого типа могут не подходить для практического применения из-за таких ограничений, как низкая стабильность, низкая активность и низкая каталитическая эффективность, особенно в промышленных условиях (Wang et al., 2012; Yang et al., 2014). Наиболее часто используемая стратегия заключается в разработке существующих биокатализаторов для совместимости с целевым производственным процессом (Dalby, 2011; Reetz, 2012; Wang et al., 2012). Благодаря быстрому развитию молекулярных методов и биоинформатики было разработано множество стратегий белковой инженерии для модификации диких ферментов с целью улучшения каталитических характеристик.Подверженная ошибкам ПЦР – одна из самых простых и популярных инженерных стратегий. Однако в основном из-за систематической ошибки, в том числе смещения кодонов и смещения амплификации в epPCR, не следует ожидать, что введение конкретных аминокислот действительно будет оптимальным (Liebeton et al., 2000; Neylon, 2004). Тем не менее, комбинация epPCR с мутагенезом с насыщением сайтов до
34
X.-J. Ли и др. / Journal of Biotechnology 188 (2014) 29–35
введение всех остальных 19 аминокислот в важные участки (горячие точки), которые ранее были определены как важные для каталитических характеристик ферментов, может устранить дефект ошибки смещения в epPCR и еще больше повысить производительность фермента.Комплементарная стратегия успешно использовалась для направленной эволюции липаз из Pseudomonas aeruginosa и Yarrowia lipolytica (Bordes et al., 2011; Reetz et al., 2001; Wen et al., 2012). Здесь три стратегии комплементарной белковой инженерии, включая epPCR, сайт-направленный мутагенез и сайт-насыщенный мутагенез, были успешно использованы для дальнейшего повышения активности Lip-T в отношении CNDE. Во-первых, мутант S63L / D232A с улучшенной активностью был подвергнут скринингу из случайной мутантной библиотеки на основе epPCR.Затем одноточечные мутанты S63L и D232A индивидуально вводили в Lip-T посредством сайт-направленного мутагенеза. Наконец, одноточечный мутант S63M с наивысшей активностью был получен из библиотеки мутагенеза с насыщением сайтов. Межфазная активация – уникальный механизм, используемый липазами (Verger, 1997). Активный центр большинства липаз полностью скрыт под коротким амфифильным спиральным сегментом (крышкой), который защищает активный центр от воздействия растворителя. Когда фермент контактирует с поверхностью раздела масло-вода, крышка претерпевает конформационные изменения, отклоняясь от вершины активного центра.В результате активный центр обнажается и обеспечивает доступ субстратам и последующим каталитическим реакциям. Следовательно, движение и конформационное изменение крышки играют роль в ферментативной активации липаз. Фрагмент ␣-спирали от V91 до N97 образует крышку, которая разворачивается двумя петлями (87 GTRS90 и 98 LAAD101), служащими петлями. Примечательно, что D67 образует Н-связи с S90 и N93, что стабилизирует N-концевую часть крышки в открытой конформации (рис. 5A). В соответствии с этим структурным наблюдением, наше предыдущее ферментативное исследование показало, что мутант D67A имел заметное снижение активности (Li et al., 2014). S63 расположен в той же петле, что и D67 (рис. 5A). Интересно, что он расположен поверх гидрофобного ядра, состоящего из остатков P60, V68, L83, F85, W122, V125, I129, F147 и L156 (рис. 5B). Мутация S63 на гидрофобный остаток M или L вызывает новое гидрофобное взаимодействие между ним и находящимся под ним гидрофобным ядром, что, вероятно, вызывает легкое движение петли 2 к гидрофобному ядру. Примечательно, что существует тонко настроенная сеть Н-связей, состоящая из остатков D62, D67, V68, которые блокируют два конца петли 2, и остатков D67, S90, N93, которые блокируют крышку и петлю 2.При этом крышка и петля 2 плотно соединяются (рис. 5А). Следовательно, мутация S63 в M или L также, вероятно, слегка вытягивает N-концевую область крышки наружу, что обеспечивает более легкий доступ субстратов к активному центру и улучшает каталитическую эффективность. Следуя этому обоснованию, мутация S63 в другие гидрофобные остатки, такие как A или V, также может усиливать активность. Однако эти мутации не были выбраны при скрининге библиотеки мутагенеза насыщения, поскольку активность этих мутантов ниже, чем у контроля (S63L).Разумно думать, что A63 и V63 имеют меньшие боковые цепи и, следовательно, более слабое взаимодействие с гидрофобным ядром. С другой стороны, также разумно объяснить, что боковые цепи лейцина, изолейцина, фенилаланина, триптофана, пролина и валина больше, что может занимать больше места и увеличивать стерические препятствия. Для сравнения, метионин имеет удлиненную стержнеобразную боковую цепь. Благодаря правильному взаимодействию с гидрофобным ядром и относительно небольшому пространству, занимаемому боковой цепью метионина, S63M приводил к оптимальному перемещению петли 2 и, таким образом, достигал максимальной активности, чем любые другие мутанты.Наконец, D232A имеет более низкую активность, что может быть связано с предпочтением полярного остатка перед неполярным на поверхности белка. Эффективный гидролиз CNDE является наиболее важным этапом химиоферментативного пути прегабалина. Высокая загрузка субстрата в биокаталитическом процессе является критическим фактором для его промышленного применения. В настоящем исследовании клетки E. coli, несущие новый
Рис. 5. Анализ модели структуры вокруг остатка 63 и иллюстрация разумного механизма для S63M с повышенной активностью.Сеть H-скрепления, состоящая из остатков D62, D67, V68, которые блокируют два конца петли 2 (оранжевая пунктирная рамка), и остатков D67, S90, N93, которые блокируют крышку (синяя пунктирная рамка) и петля 2, плотно соедините крышку и петлю 2 (А). Водородные связи показаны пунктирными линиями. Остаток 63 лежит поверх гидрофобного ядра, состоящего из остатков P60, V68, L83, F85, W122, V125, I129, F147 и L156 (B). Взаимодействие между Met63 (окрашенным в светло-розовый цвет) с гидрофобным ядром вызывает легкое движение петли 2 к гидрофобному ядру (левая стрелка представляет), которое слегка отодвигает N-концевую область крышки (правая стрелка представляет), облегчает доступ субстратов к активному центру и улучшает каталитическую эффективность.(Для интерпретации ссылки на цвет в легенде этого рисунка читатель отсылается к веб-версии этой статьи.)
Разработаннаялипаза была использована для кинетического разрешения CNDE с образованием (3S) -2-карбоксиэтил-3-циано. -5-метилгексановая кислота с улучшенными характеристиками. Даже при очень высокой загрузке субстрата (3 M, 765 г / л) сконструированная липаза могла эффективно гидролизовать CNDE при высокой конверсии. Конверсия 45,2% с КПД 98% была достигнута через 24 часа, что полностью соответствует уровню разрешающей способности CNDE, катализируемой липолазой® (Martinez et al., 2008). Более того, этот процесс намного более рентабелен, так как 5% (мас. / Об.) Влажного
X.-J. Ли и др. / Journal of Biotechnology 188 (2014) 29–35
цельных клеток использовали в качестве биокатализатора вместо 8% (мас. / Об.) Коммерчески доступной липазы Lipolase®. В этом смысле модифицированная липаза будет привлекательной и конкурентоспособной альтернативой Lipolase® для крупномасштабного производства прегабалина. В заключение, три дополнительные стратегии белковой инженерии были успешно использованы для улучшения гидролитической активности Lip-T в отношении CNDE.Был получен мутант S63M с повышенной каталитической эффективностью в 4,5 раза по сравнению с Lip-T и выявлены ключевые остатки, ответственные за повышение каталитической эффективности. Кроме того, мутант S63M использовали для кинетического разрешения CNDE при высокой загрузке субстрата (765 г / л). Эти результаты показали, что сконструированная липаза является многообещающим биокатализатором для кинетического разрешения CNDE. Благодарности. Работа выполнена при финансовой поддержке Национальной программы исследований и разработок высоких технологий Китая (программа 863, 2012AA022201), ключевых научных и технологических программ провинции Чжэцзян (2012C03005-2).Приложение A. Дополнительные данные Дополнительные данные, связанные с этой статьей, можно найти в онлайн-версии по адресу http://dx.doi.org/10.1016/j.jbiotec. 2014.08.004. Ссылки Bassegoda, A., Pastor, FIJ, Diaz, P., 2012. Штамм Rhodococcus sp CR-53 lipR, первый член нового семейства бактериальных липаз (семейство X), демонстрирующий необычную оксианионную дыру Y-типа, похожую на Candida antarctica. клан липазы. Прил. Environ. Microbiol. 78, 1724–1732. Беллиотти, Т.Р., Капирис, Т., Экхато, И.В., Кинсора, Дж.Дж., Филд, М.Дж., Хеффнер, Т.Г., Мельцер, Л.Т., Шварц, Дж. Б., Тейлор, С.П., Торп, А.Дж., Вартаниан, М.Г., Уайз, Л.Д., Чжи-Су, Т., Вебер, М.Л., Вустров, Д.Д., 2005. Взаимосвязь между структурой и активностью прегабалина и аналогов, нацеленных на альфа (2) -дельта-белок. J. Med. Chem. 48, 2294–2307. Bordes, F., Tarquis, L., Nicaud, J.M., Marty, A., 2011. Выделение термостабильного варианта липазы Lip2 из Yarrowia lipolytica путем направленной эволюции и более глубокого понимания задействованных механизмов денатурации.J. Biotechnol. 156, 117–124. Bornscheuer, U.T., Huisman, G.W., Kazlauskas, R.J., Lutz, S., Moore, J.C., Robins, K., 2012. Разработка третьей волны биокатализа. Nature 485, 185–194. Bradford, M., 1976. Быстрый и чувствительный метод количественного определения количества белка в микрограммах, использующий принцип связывания белок-краситель. Анальный. Biochem. 72, 248–252. Брэди, Л., Брзозовски, А.М., Деревенда, З.С., Додсон, Э., Додсон, Г., Толли, С., Туркенбург, Дж. П., Кристиансен, Л., Хугейенсен, Б., Норсков, Л., Thim, L., Menge, U., 1990. Триада сериновой протеазы образует каталитический центр триацилглицеринлипазы. Nature 343, 767–770. Чен, Ю.С., Ли, X., Ченг, Р.Н., Чжао, С.Ю., 2011. Последние разработки в области синтеза прегабалина. Подбородок. J. Org. Chem. 31, 1582–1594. Chronopoulou, E.G., Labrou, N.E., 2011. Мутагенез с насыщением сайтов: мощный инструмент для структурно-ориентированного дизайна библиотек комбинаторных мутаций. В: Dunn, B. (Ed.), Current Protocols in Protein Science, vol. 63. Джон Вили, Нью-Йорк, стр.26.6.1–26.6.10. Далби П.А., 2011. Стратегия и успех направленной эволюции ферментов. Curr. Opin. Struc. Биол. 21, 473–480. Фернандес-Лафуэнте, Р., 2010. Липаза из Thermomyces lanuginosus: использование и перспективы в качестве промышленного биокатализатора. J. Mol. Катал. B: Энзим. 62, 197–212.
35
Гупта Р., Гупта Н., Рати П., 2004. Бактериальные липазы: обзор производства, очистки и биохимических свойств. Прил. Microbiol. Biotechnol. 64, 763–781. Йегер, К.Э., Дейкстра, Б.W., Reetz, M.T., 1999. Бактериальные биокатализаторы: молекулярная биология, трехмерные структуры и биотехнологические применения липаз. Анну. Rev. Microbiol. 53, 315–351. Jaeger, K.E., Eggert, T., 2002. Липазы для биотехнологии. Curr. Opin. Biotechnol. 13, 390–397. Ласковски, Р.А., Макартур, М.В., Мосс, Д.С., Торнтон, Дж. М., 1993. Procheck – программа для проверки стереохимического качества белковых структур. J. Appl. Кристаллогр. 26, 283–291. Ли, X.J., Чжэн, R.C., Ма, H.Y., Zheng, Y.G., 2014.Разработка липазы Lip от Thermomyces lanuginosus: создание нового биокатализатора для эффективного биосинтеза хирального промежуточного соединения прегабалина. Прил. Microbiol. Biotechnol. 98, 2473–2483. Liebeton, K., Zonta, A., Schimossek, K., Nardini, M., Lang, D., Dijkstra, B.W., Reetz, M.T., Jaeger, K.E., 2000. Направленная эволюция энантиоселективной липазы. Chem. Биол. 7, 709–718. Марсьелло, М., Филис, М., Паломо, Дж. М., 2012. Различные стратегии повышения активности липазных катализаторов. Катал. Sci.Technol. 2, 1531–1543. Мартинес, С.А., Ху, С., Дюмон, Ю., Тао, Дж., Келлехер, П., Талли, Л., 2008. Разработка химико-ферментативного процесса производства прегабалина. Орг. Процесс Res. Dev. 12, 392–398. Морис-Варас, Ф., Шах, А., Айкенс, Дж., Надкарни, Н.П., Роззелл, Дж. Д., Демирджян, Д.К., 1999. Визуализация реакций, катализируемых ферментами, с использованием индикаторов pH: быстрый скрининг библиотек гидролаз и оценка энантиоселективность. Биоорг. Med. Chem. 7, 2183–2188. Мукерджи, Х., Мартинес, К.А., 2011. Биокаталитический путь к хиральным предшественникам замещенных–аминокислот. ACS Catal. 1, 1010–1013. Нардини, М., Дейкстра, Б.В., 1999. Альфа / бета-гидролазные складчатые ферменты: семья продолжает расти. Curr. Opin. Struc. Биол. 9, 732–737. Нейлон, С., 2004. Химические и биохимические стратегии рандомизации последовательностей ДНК, кодирующих белок: методы построения библиотек для направленной эволюции. Nucl. Acids Res. 32, 1448–1459. Петерс, Г.Х., Свендсен, А., Лангберг, Х., Винд, Дж., Паткар, С.А., Токсваерд, С., Kinnunen, P.K.J., 1998. Активный серин участвует в стабилизации петли активного центра липазы Humicola lanuginosa. Биохимия 37, 12375–12383. Куин, М. Б., Шмидт-Даннерт, К., 2011. Разработка биокатализаторов: от эволюции к созданию. ACS Catal. 1, 1017–1021. Ритц, М.Т., 2012. Лабораторная эволюция стереоселективных ферментов как средство расширения набора инструментов химиков-органиков. Тетраэдр 68, 7530–7548. Reetz, M.T., Wilensek, S., Zha, D.X., Jaeger, K.E., 2001. Направленная эволюция энантиоселективного фермента посредством комбинаторного множественного кассетного мутагенеза.Энгью. Chem. Int. Эд. 40, 3589–3591. Родригес, Р.С., Ортис, К., Беренгер-Мурсия, А., Торрес, Р., Фернандес-Лафуэнте, Р., 2013. Изменение активности и селективности ферментов путем иммобилизации. Chem. Soc. Ред. 42, 6290–6307. Sambrook, J., Russell, D.W., 2001. Молекулярное клонирование: лабораторное руководство, 3-е изд. Лаборатория Колд-Спринг-Харбор, Нью-Йорк. Шмид, Р.Д., Вергер, Р., 1998. Липазы: межфазные ферменты с привлекательными применениями. Энгью. Chem. Int. Эд. 37, 1609–1633. Сильверман, Р.Б., 2008. От фундаментальной науки до наркотика-блокбастера: открытие Lyrica.Энгью. Chem. Int. Эд. 47, 3500–3504. Вергер, Р., 1997. «Межфазная активация» липаз: факты и артефакты. Trends Biotechnol. 15, 32–38. Ван, М., Си, Т., Чжао, Х.М., 2012. Разработка биокатализатора путем направленной эволюции. Биоресурсы. Technol. 115, 117–125. Вэнь, С., Тан, Т.В., Чжао, Х.М., 2012. Повышение термостабильности липазы Lip2 из Yarrowia lipolytica. J. Biotechnol. 164, 248–253. Ян, Х., Ли, Дж., Шин, Х.-д., Ду, Г., Лю, Л., Чен, Дж., 2014. Молекулярная инженерия промышленных ферментов: последние достижения и перспективы на будущее.Прил. Microbiol. Biotechnol. 98, 23–29. Япоуджян, С., Иванова, М.Г., Бжозовски, А.М., Паткар, С.А., Винд, Дж., Свендсен, А., Вергер, Р., 2002. Связывание липазы Thermomyces (Humicola) lanuginosa со смешанными мицеллами цис-паринарной кислоты. кислота / NaTDC – резонансный перенос энергии флуоресценции и кристаллографические исследования. Евро. J. Biochem. 269, 1613–1621. Zheng, R.C., Wang, T.Z., Fu, D.J., Li, A.P., Li, X.J., Zheng, Y.G., 2013. Биокаталитический синтез хирального промежуточного соединения прегабалина с высокой загрузкой субстрата с помощью недавно выделенной Morgarella morganii ZJB-09203.Прил. Microbiol. Biotechnol. 97, 4839–4847.
3BHE022293R0101 | ABB | ПК D232 A Коммуникационный модуль ввода / вывода
ABB 3BHE022293R0101
- 3BHE022293R0101
- ABB
- ПК D232 A
- Коммуникационный модуль ввода / вывода
4D Controls принадлежит и управляется командой инженеров по автоматизации и электротехнике.С 2005 года мы поставляем промышленным предприятиям мира новое, бывшее в употреблении и избыточное оборудование для управления и автоматизации.
Мы продаем широкий ассортимент запчастей. Например, программируемые логические контроллеры (PLC), распределенные системы управления (DCS), человеко-машинный интерфейс (HMI), приводы с регулируемой скоростью (VSD), источники питания, датчики давления и все, что между ними. Многие из этих продуктов, которые мы храним, устарели, и их трудно найти.
В частности, мы гордимся тем, что поставляем профессиональные, надежные и экономичные альтернативы новым элементам управления и автоматизации от большого количества различных брендов.Такие как Allen Bradley, Siemens, Schneider, GE Fanuc, Pilz, Symax, Yokogawa, Foxboro, Triconnex и многие другие.
Цель нашей компании – предоставить точную замену марки, номера детали, серии или встроенного ПО при необходимости. Мы верим в нашу продукцию. Таким образом, на все наши детали предоставляется 2-летняя гарантия.
Наш собственный ремонтно-испытательный центр, укомплектованный и управляемый квалифицированными специалистами и инженерами по электронике, электрике, КИПиА и автоматизации, может отремонтировать широкий спектр продукции.Кроме того, на все наши ремонтные работы распространяется двухлетняя гарантия. Свяжитесь с нами для получения более подробной информации о наших возможностях ремонта.
У нас есть запасы на складах как в США, так и в Австралии, и мы предлагаем различные варианты перевозки. Как правило, мы отправляем все товары в течение одного рабочего дня после получения оплаты.
* 4D Controls не является авторизованным дистрибьютором, реселлером или представителем какой-либо из представленных продуктов. Использование любых брендов, логотипов, товарных знаков и наименований только в целях идентификации.
3BHE022293R0101 | ABB | PC D232 A Коммуникационный модуль ввода / вывода был последний раз изменен: 23 августа 2021 года администратором
D232a Сух Сагар Тулси Марг, Бани Паркджайпур, 302016 Номер пин-кода
D232a Сух Сагар Тулси Марг, Бани Паркджайпур, 302016
Полное имя: D232a Сух Сагар Тулси Марг, Бани Паркджайпур, 302016
Пин-код: Талук: Джайпур
Район: Шастри Нагар
Район: Джайпур
Штат: Раджастан
Почтовые отделения рядом с D232a Сух Сагар Тулси Марг, Бани Паркджайпур, 302016
Больше похоже на D232a Сух Сагар Тулси Марг, Бани Паркджайпур, 302016
D232a Сух Сагар Тулси Марг, Бани Паркджайпур, 302016 расположен в районе , Джайпур талука, округ Джайпур, в штате Раджастхан, а пин-код – 302016 .
- D238a Бихари Марг, Gulab Residency Flat №305 Бани Паркджайпур, 302016
- D24 Akar Marg Bani Park, Джайпур, 302016
- D232a Сух Сагар Тулси Марг, Бани Паркджайпур, 302016
- D234 Behind Collectory, Banipark Jaipur, 302016
- D24 Be Veer Durga Das Colony Hari Nagar, Shastri Nagarjaipur, 302016
- D237 Bhihari Marg Bani Park, Джайпур, 302016
- D235 Bihari Marg, Bani Parkjaipur, 302016
- D237 Bihari Marg Banipark, Джайпур, 302016
- D234, Bihari Marg, почтовое отделение Banipark, Shastri Nagar, Jaipur, 302016
- D234 Bihari Marg Behaind Collectorate, Bani Prak Jaipur, 302016
- D234 Bihari Marg Behind, Collectorate Banipark Jaipur, 302016
- D238 Iiird Follr, 3 Bihari Marg, Bani Parkjaipur, 302016
- D24 2nd Lane Rpa Road Dwarkapuri, Sanjay Colony Shastri Nagar Jaipur, 302016
- D24 Pink City Apartments, Акар Марг, Бани Парк, Питал Фабрика Джайпур, 302016
- D24 Pink City Apartments Blocka, квартира №401, Акар Марг, Бани Паркджайпур, 302016
- D239 302 Pratap Enclave Biharii Marg, Baniparkjaipur, 302016
- D237 Rao Veer Vikram Singh, Bihari Marg Baniparkjaipur, 302016
- D239 Shop Nob2 Propbal Krishan Balsaria, Pratap Enclav, Bihri Margbanipark Jaipur, 302016
- D238, Shri Gopalam Apartmentg4, Bihari Marg, Shastri Nagar, Джайпур, 302016,
- D238 Srigopalam Apartment Bihari Marg Bani Park, Джайпур, 302016
Карта проезда D232a Sukh Sagar Tulsi Marg, Bani Parkjaipur, 302016
Почтовые отделения в Джайпуре
Звезда Индианаполиса из Индианаполиса, Индиана, 15 ноября 2003 г. · Стр. 8
ЗВЕЗДА ИНДИАНАПОЛИСА WWW.INDYSTAR.COM HamiltonAM st СУББОТА, 15 НОЯБРЯ 2003 г. H5 J $ & amp; a t r i ‘Tiff & amp; ivO’ T ‘kt’ SKikSSS-fc. 96 MERCURY COUGAR AT.V8, 65K3174A1 4995 долларов США 00 TOYOTA COROLLA 33K, AT C478A 7 995 долларов США, fs am j1! “00 KIERCURY VILLAGER SPORT, 7-ПАССАЖИРСКИЙ 3605A s 10 935 00 долларов США FORD EXPLORER XL MA V6C50 1,95 м. LX 11K, CD, ATP6064 s 12,995 01 MAZDA MSLLENIA 39K, КОЖА P6080 15 991 $ 03 MERCURY GR MARQUIS LS 14K, КОЖА P6089 18 995 $ яма W ii T 1 J ti 1 I it f nWii nli.-, tj. 00 LINCOLN NAVIGATOR 54K, 6 ДИСКОВ CD, HTD. СИДЕНЬЯ 3675A 24 495 долл. 04 MAZDA RX8 1 800 миль, 37 954 долл. США P6048A s31 995 См. -US 93 FORD MUSTANG 5 SPD., 77K, V6 P6033A “6995 99 СТАРЫЙ INTRIGUE GS, ЗАГРУЖЕННЫЙ D232A ‘7995 03 FORD FOCX 27K, AT, Z P6000 10 995 долларов США 02 HYUNDAI SONATA 31 K, ЗАГРУЖЕННЫЙ, AT, NICE P601 2B 1 995 долларов США 00 NISSAN QUEST 58K, ENT.PKG., 4000 млрд долларов США 1 2995 долларов США 00 MAZDA MIATA 25K, SE, 6 SPD., ОБЕ ВЕРШИНЫ P6077 1 5700 долларов США CHER JEEP GRAND LAREDO, 4X4 P6020 $ 1 9995 w- 00 BMW 5281 3777A $ 24 995 02 LINCOLN NAVIGATOR LEATHER, SR00F, REAR AC P6043 s31 935 ‘- – – i (U.S. 31. (MertlDi alt iWfffc Mflft, MO., 77lfflS. At OTlilJW 97 MAZDA 626 LX V6, AT C595A 6995 долл. 1 98ISUZUAMIG0 35K, 5-SPD., 4X4 P6017A 7995 долл. США-! 98ACURA3.0CL 4695 $ 01 MERCURY SABLE LS 23K, КОЖА, ЛЮК P6034 $ 1 1995 99 LINCOLN TOWN CAR SIG. СЕРИЯ, 62K 3647A $ 1 2995 -… 4 “n 02 MERCURY VILLAGER SPORT 25K, С НАГРУЗКОЙ, CD P6075 $ 1 5,9953ZB 02 CHEVLAZERA LT 20953 $ EXPEDITION XLT 30K, 8 PASS. P6035 24 995 $ 03 GMC YUKON XL 13K, ЗАГРУЖЕННЫЙ, XL DENAU 4069A ’38, 995 i CS) ErS…, Sfl a UU Kid 00 MERCURY GRAND MARQUIS PW, PL, V8 3702A “7 495 долларов США 97 MERC. GRAND MARQUIS LS 52K, LTHR., FULL PWR. 4058A, 8 995 долларов США WOW 01 MAZDA 626 LX V6, SUNROOF P6077 1 1495 долларов США 02 MAZDA 62 LX AT, 22K P6002 1 995 долларов 03 BUICK ОТЗЫВАЕТ FULL POWER STK P5903 1 2995 долларов 02 JEEP LIBERTY SPORT 3,7 л, 4X4 P6008 16 795 долларов 17K, ЗАГРУЖЕННЫЙ, ОЧЕНЬ КРАСИВЫЙ P6069 s39,995 -1 691th – f Tiiir “” – “: – i -irBriir -” lif – ‘-1 – 9-0 WES., ff jmlBUJM в i. ufo 99 MERCURY COUGAR 57K, 5-SPD., SOOF D1 10A $ 7 995 99 CHRY. ГОРОД & amp; СТРАНА LXI, КОЖА, 4 ДР. P5861A $ 9995 02 MAZDA B3000 X-CAB DUAL SPORT, 19K, AT, tP5998 $ 11, 595 00 FORD MUSTANG 52K, AT, КОЖА, CD P6016 $ 1 1995 5 00 MAZDA MIATA AUTO, 33K P6047A $ 1 2995 02 FORD F-1 50 X -КАБИНА 27K, AT, XLT P6039 $ 17 495 iiaisasa Mi t- 6v. . . . 02 LINCOLN LS V6, КОЖА, ЛЮК P6087 23995 долларов США S3 JAGUAR XJR 52K, SHARP D145A 2S, 995 J WM I rr – -i – i’.IS -1 03 LINCOLN NAVIGATOR 13K, С НАГРУЗКОЙ P5872 40995 долларов США.3Т, В; Fff., Пороги fl-G 01 PQNTIAC SUNFIRE AT, AC 4052A 7995 долларов США 00 JEEP WRANGLER 55K, 5-SPD h4759B 1 0995 долларов США 02 MAZDA PROTEGE 5 5SPD., 17KP6037 1 1895 долларов США 01 MAZDA TRIBUTE 40K, 5-SPD 1,9195 долларов США PRIX GT SROOF, CD, 35K 3715A 14 709 вон 00 LINCOLN CONTINENTAL 39K, СЕРТИФИЦИРОВАННАЯ, С НАГРУЗКОЙ 3534A 17 995 долл. 02 LINCOLN LSE V8, кожа, люк P6086 24 995 долл. 02 CKEVYTAH0ELT 33K, 7-ПАССОВ. ., ЗАГРУЖЕННЫЙ P6071 48 995 долл.
