Y-образный элемент 30-45 для полукруглого конька Mtile кофе
Конек полукруглый тройной Metrotile – доборный элемент, который применяется для скатных кровель. Является важной составляющей крыши Metrotile.
Позволяет красиво оформить верхнюю часть кровли и придать конструкции эстетически завершённый вид. Вместе с этим он обеспечивает циркуляцию воздуха в подкрышном пространстве и принимает на себя основную нагрузку от неблагоприятных внешних факторов.
Полукруглые коньковые элементы укладываются с нахлестом, образуя специальный замок. Коньковые элементы крепятся гвоздями к верхним брускам обрешетки.
Расход – 0,9 штук/погонный метр.
При производстве тройных полукруглых коньков Metrotile используется эксклюзивная запатентованная технология. Композитные элементы отличаются высокой стойкостью к влаге, жаре и холоду. Они обладают высоким классом пожаробезопасности, не выделяют токсинов и вредных веществ.
Средний срок службы изделий составляет 50 лет.
В своей структуре имеет восемь слоев. Основа – это качественный стальной лист, который во избежание коррозии имеет двустороннее покрытие алюмоцинковым сплавом. Надежную защиту черепице обеспечивает покрытие из крошки натурального камня. Акриловый клей является связующим звеном металла и каменного покрытия. Он обеспечивает сохранность каменной крошки на поверхности листа.
Конек является универсальными аксессуаром для любой коллекции композитной черепицы Metrotile. В случае необходимости торцы коньков закрываются заглушками.
Преимущества коньковых элементов для композитной черепицы Metrotile:
– Долговечность за счет стального основания с алюмоцинковым покрытием.
– Не выцветают на солнце благодаря глубокой термальной окраске каменной посыпки.
– Бесшумность при дожде и граде, благодаря защитному слою из гранулята натурального камня.
– Высокая пожаробезопасность. Запатентованный многослойный состав материала обеспечивает наивысший класс сопротивления природным катаклизмам.
– На всю продукцию предоставляется письменная гарантия 30 лет.
Элемент RefBy (Cell_Type complexType) (Visio XML)
Элемент Cell (Раздел Тег действия)
Cell_Type
Определяет одно свойство для тега действий на форме или странице.
Элемент Cell (строка Actions)
Cell_Type
Указывает одно свойство действия, связанного с настраиваемой командой в меню ярлыка или тега действий.
Элемент Cell (строка ArcTo)
Cell_Type
Содержит координаты x, координаты y или лук круговой дуги.
Элемент Cell (Раздел символов)
Cell_Type
Указывает атрибут форматирования для текстового запуска фигуры, например шрифта, цвета, стиля, дела, позиции по отношению к базовому или размеру точки.
Элемент Cell (строка Connection)
Cell_Type
Содержит x- или y-координаты, горизонтальное или вертикальное направление или введите для одной точки подключения на фигуре.
Элемент Cell (строка Controls)
Cell_Type
Содержит свойство для определенной ручки управления, определенной для фигуры.
Элемент Cell (строка Ellipse)
Cell_Type
Содержит x или y-координаты центральной точки эллипса и две точки на эллипсе.
Элемент Cell (строка EllipticalArcTo)
Cell_Type
Содержит x- или y-координаты конечной точки эллиптической дуги, x или y-координаты точек управления на дуге, угол от оси x до основной оси эллипса или соотношение между основными и небольшими осями эллипса.
Элемент Cell (Полевой раздел)
Cell_Type
Отображает функции и формулы, вставленные в текст фигуры с помощью диалогового окна Field.
Элемент Cell (Раздел Заполнения Градиента)
Cell_Type
Содержит цвет, прозрачность и расположение остановки градиента для градиента заполнения.
Элемент Cell (Раздел Геометрия)
Cell_Type
Определяет свойства, определяющих форматирование и поведенческие свойства в отношении линий и дуг, которые составляют раздел Геометрия.
Элемент Cell (строка Hyperlink)
Cell_Type
Содержит сведения для одной гиперссылки, связанной с фигурой. Фигура будет содержать одну строку гиперссылки для каждой гиперссылки.
Элемент Cell (строка InfiniteLine)
Cell_Type
Содержит x или y-координаты двух точек на бесконечной строке.
Элемент Cell (Раздел Layer)
Cell_Type
Указывает одно свойство для слоя или его свойств для страницы.
Элемент Cell (раздел Line Gradient)
Cell_Type
Содержит цвет, прозрачность или положение остановки градиента для градиента строки.
Элемент Cell (строка LineTo)
Cell_Type
Содержит x-или y-координаты конечной вершины сегмента прямой линии.
Элемент Cell (строка MoveTo)
Cell_Type
Содержит x или y-координаты первой вершины фигуры или представляет x- или y-координаты первой вершины после разрыва пути.
Элемент Cell (строка NURBSTo)
Cell_Type
Содержит x- или y-координаты, положение второго до последнего узла, положение последнего веса, положение первого узла, положение первого веса или формулу для неинформированной рациональной B-spline (NURBS).
Элемент Cell (Раздел Абзац)
Cell_Type
Указывает атрибут форматирования абзаца для текста фигуры, например вмятины, интервалы строк, пули или горизонтальное выравнивание абзацев.
Элемент Cell (строка PolyLineTo)
Cell_Type
Содержит x или y-координаты последней точки полилиновой или полилиновой формулы.
Элемент Cell (строка RelCubBezTo)
Cell_Type
Содержит x- или y-координаты конечной точки кривой кубического Bézier относительно ширины и высоты фигуры, x- или y-координаты точки управления в начале ширины и высоты относительной формы кривой или x-или y-координаты точки управления окончанием ширины и высоты кривой относительной формы.
Элемент Cell (строка RelEllipticalArcTo)
Cell_Type
Содержит x- или y-координаты конечной точки эллиптической дуги относительно ширины и высоты фигуры, x- или y-координат пунктов управления на дуге относительно ширины и высоты фигуры, угла от оси x до основной оси эллипса или соотношения между основными и небольшими осями эллипса.
Элемент Cell (RelLineTo Row)Cell
Cell_Type
Содержит x-или y-координаты конечной вершины сегмента прямой линии относительно ширины и высоты фигуры.
Элемент Cell (строка RelMoveTo)
Cell_Type
Содержит x- или y-координаты первой вершины фигуры или x- или y-координаты первой вершины после перерыва в пути, относительно высоты и ширины фигуры.
Элемент Cell (Раздел RelQuadBezTo
Cell_Type
Содержит x или y-координаты конечной точки квадратной кривой Bézier по отношению к ширине и высоте фигуры или x- или y-координатам точки управления ширины и высоты кривой относительной формы.
Элемент Cell (Scratch Section)
Cell_Type
Указывает область работы для ввода и тестирования формул, на которые могут ссылаться другие ячейки.
Элемент Cell (Раздел Фигура данных
Cell_Type
Указывает одно свойство данных фигуры.
Элемент Cell (строка SplineKnot)
Cell_Type
Содержит x или y-координаты для точки управления spline или узла spline.
Элемент Cell (Раздел SplineStart
Cell_Type
Содержит x- или y-координаты для второй точки управления spline, ее второго узла, первого узла, последнего узла или степени spline.
Элемент Cell (Раздел Вкладок)
Cell_Type
Указывает свойство, которое управляет положением остановки или выравниванием стоп-вкладок формы и стиля.
Элемент Cell (Раздел ячейки, определяемой пользователем)
Cell_Type
Одно свойство указанного пользователем фрагмента информации, на которое можно ссылаться другими ячейками и средствами надстройки.
химический элемент Иттрий Yttrium — “Химическая продукция”
Что такое
Иттрий, yttrium, характеристики, свойства
Иттрий — это химический элемент Y элемент побочной подгруппы третьей группы пятого периода периодической системы химических элементов Д. И. Менделеева, с атомным номером 39. Обозначается символом Y (лат. Yttrium). Простое вещество иттрий — металл светло-серого цвета. Существует в двух кристаллических модификациях: α-Y с гексагональной решёткой типа магния, β-Y с кубической объёмноцентрированной решёткой типа α-Fe, температура перехода α↔β 1482 °C.
Иттрий класс химических элементов
Элемент Y — относится к группе, классу хим элементов (…)
Элемент Y свойство химического элемента Иттрий Yttrium
Основные характеристики и свойства элемента Y…, его параметры.
формула химического элемента Иттрий Yttrium
Химическая формула Иттрия:
Атомы Иттрий Yttrium химических элементов
Атомы Yttrium хим. элемента
Yttrium Иттрий ядро строение
Строение ядра химического элемента Yttrium — Y,
История открытия Иттрий Yttrium
Открытие элемента Yttrium — В 1794 году финский химик Юхан (Иоганн) Гадолин (1760—1852) выделил из минерала иттербита оксид элемента, который он назвал иттрием — по названию шведского населённого пункта Иттербю, находящегося на острове Ресарё, входящем в Стокгольмский архипелаг (иттербит был найден здесь в заброшенном карьере). В 1843 году Карл Мосандер доказал, что этот оксид на самом деле является смесью оксидов иттрия, эрбия и тербия и выделил из этой смеси Y2O3.
Металлический иттрий, содержащий примеси эрбия, тербия и других лантаноидов, был получен впервые в 1828 году Фридрихом Велером.
Иттрий Yttrium происхождение названия
Откуда произошло название Yttrium …
Распространённость Иттрий Yttrium
Как любой хим. элемент имеет свою распространенность в природе, Y— химический аналог лантана. Кларк 26 г/т, содержание в морской воде 0,0003 мг/л. Иттрий почти всегда содержится вместе с лантаноидами в минеральном сырье. Несмотря на неограниченный изоморфизм, в группе редких земель в определённых геологических условиях возможна раздельная концентрация редких земель иттриевой и цериевой подгрупп.
Цериевая подгруппа
С щелочными породами и связанными с ними постмагматическими продуктами преимущественное развитие получает цериевая подгруппа, а с постмагматическими продуктами гранитоидов с повышенной щёлочностью — иттриевая. Большинство фторкарбонатов обогащено элементами цериевой подгруппы. Многие тантало-ниобаты содержат иттриевую подгруппу, а титанаты и титано-тантало-ниобаты — цериевую.
Yttrium — получение элемента — соединения иттрия получают из смесей с другими редкоземельными металлами экстракцией и ионным обменом. Металлический иттрий получают восстановлением безводных галогенидов иттрия литием или кальцием c последующей отгонкой примесей.
Физические свойства Иттрий Yttrium
Основные свойства Yttrium
Изотопы Yttrium Иттрий
Наличие и определение изотопов Yttrium
Y свойства изотопов Иттрий Yttrium
…
Химические свойства Иттрий Yttrium
Определение химических свойств Yttrium
Меры предосторожности Иттрий Yttrium
Внимание! Внимательно ознакомьтесь с мерами безопасности при работе с Yttrium
Стоимость Иттрий Yttrium
Рыночная стоимость Y, цена Иттрий Yttrium
Примечания
Список примечаний и ссылок на различные материалы про хим. элемент Y
M22-XC-Y / | : 216407.
Технические характеристики для подтверждения типа конструкции
Номинальный ток для указания потери мощности [In]
0 A
Потеря мощности на полюс, в зависимости от тока [Pvid]
0 W
Потеря мощности оборудования, в зависимости от тока [Pvid]
0 W
Статическая потеря мощности, не зависит от тока [Pvs]
0 W
Способность отдавать потери мощности [Pve]
0 W
Мин. рабочая температура
-25 °C
Макс. рабочая температура
+70 °C
Проверка конструкции IEC/EN 61439
10.2 твёрдость материалов и деталей10.2.2 Коррозионная стойкость
Требования производственного стандарта выполнены.
10.2 твёрдость материалов и деталей10. 2.3.1 Нагревостойкость изоляции
Требования производственного стандарта выполнены.
10.2 твёрдость материалов и деталей10.2.3.2 Сопротивление изоляционных материалов при обычном нагреве
Требования производственного стандарта выполнены.
10.2 твёрдость материалов и деталей10.2.3.3 Сопротивление изоляционных материалов при сильном нагреве
Требования производственного стандарта выполнены.
10.2 твёрдость материалов и деталей10. 2.4 Устойчивость к ультрафиолетовому излучению
По запросу
10.2 твёрдость материалов и деталей10.2.5 Подъём
Не имеет значения, поскольку необходимо оценить всё коммутационное оборудование.
10.2 твёрдость материалов и деталей10.2.6 Испытание на удар
Не имеет значения, поскольку необходимо оценить всё коммутационное оборудование.
10.2 твёрдость материалов и деталей10.2.7 Ярлыки
Требования производственного стандарта выполнены.
10.3 Класс защиты изоляции
Не имеет значения, поскольку необходимо оценить всё коммутационное оборудование.
10.4 Воздушные промежутки и пути утечки тока
Требования производственного стандарта выполнены.
10.5 Защита от удара электрическим током
Не имеет значения, поскольку необходимо оценить всё коммутационное оборудование.
10. 6 Монтаж оборудования
Не имеет значения, поскольку необходимо оценить всё коммутационное оборудование.
10.7 Внутренние электрические цепи и соединения
Находится в сфере ответственности компании, монтирующей распределительные устройства.
10.8 Подключения проводов, введённых снаружи
Находится в сфере ответственности компании, монтирующей распределительные устройства.
10.9 Свойства изоляции10.9.2 Электрическая прочность при рабочей частоте
Находится в сфере ответственности компании, монтирующей распределительные устройства.
10.9 Свойства изоляции10.9.3 Прочность по отношению к импульсному напряжению
Находится в сфере ответственности компании, монтирующей распределительные устройства.
10.9 Свойства изоляции10.9.4 Проверка оболочек кабелей из изолирующего материала
Находится в сфере ответственности компании, монтирующей распределительные устройства.
10.10 Нагрев
Неприемлемо.
10. 11 Стойкость к коротким замыканиям
Находится в сфере ответственности компании, монтирующей распределительные устройства. Соблюдать указания для коммутационных устройств.
10.12 Электромагнитная совместимость
Находится в сфере ответственности компании, монтирующей распределительные устройства. Соблюдать указания для коммутационных устройств.
10.13 Механическая функция
Для устройства требования считаются выполненными, если были соблюдены данные инструкции по монтажу (IL).
Риск-менеджмент — ключевой элемент в цифровой трансформации промышленного предприятия // Risk Management as a Key Element in the Digital Transformation of an Industrial Enterprise
D. Pashchenko S.,
N. Komarov M.,
Д. Пащенко С. and
Н. Комаров М. Additional contact information Н. Комаров М.: ФГУП «ЦНИИ «ЦЕНТР», Москва
Мир новой экономики // The world of new economy, 2021, vol. 15, issue 1, 14-27
Abstract:
The Russian industry is entering digital transformation with some delay, and the implementation of accompanying large-scale changes contains a significant number of specific risks. Creating a promising business model and the technological outline of an industrial enterprise that implements it takes place under conditions of high uncertainty. The article proposes a general approach and elements of a practical risk model for planning and carrying out an industrial enterprise’s digital transformation to reduce the uncertainty of the process in its signifiant aspects. The proposed management solutions help reduce the likelihood of a negative impact of accompanying general and specifi risks, build a formalized model of risk management, and increase transparency in managing relevant practical projects. The article also discusses fiancial reserves usage as a tool for active risk management.
Российская промышленность с некоторым опозданием вступает в цифровую трансформацию, и внедрение сопутствующих масштабных изменений содержит значительное количество специфических рисков. Создание перспективной бизнес-модели и реализующего ее технологического контура промышленного предприятия происходит в условиях высокой неопределенности. В статье предложен общий подход и элементы практической риск-модели для планирования и проведения цифровой трансформации промышленного предприятия. Предложенные управленческие решения позволяют снизить вероятности негативного влияния сопутствующих общих и специфических рисков, построить формализованную модель риск-менеджмента, повысить прозрачность в управлении соответствующими практическими программами прикладных проектов. Также в статье рассматривается использование финансовых резервов как инструмента в активном управлении рисками.
Keywords: risk management; fiancial management; digital transformation; change management; fiancial reserves; риск-менеджмент; финансовое управление; цифровая трансформация; управление изменениями; финансовые резервы (search for similar items in EconPapers) Date: 2021-03-24 References: Add references at CitEc Citations: Track citations by RSS feed
More articles in Мир новой экономики // The world of new economy from Финансовый университет при Правительстве Российской Федерации // Financial University under The Governtment оf The Russian Federation Bibliographic data for series maintained by Алексей Скалабан ().
Yttrium – Информация об элементе, свойства и использование
Расшифровка:
Химия в ее элементе: иттрий
(Promo)
Вы слушаете Химию в ее элементе, представленную вам Chemistry World , журналом Королевского химического общества.
(Конец промо)
Meera Senthilingam
На этой неделе последний из элементов, обнаруженных в маленьком городке Иттерби, и его соединения, кажется, имеют множество применений.
Саймон Коттон
Ю. Это не вопрос. Y – символ элемента иттрия.
Примерно 20 лет назад большинство ученых не слышали о нем, за исключением смутных указаний на его место в периодической таблице, под скандием и над лантаном. Некоторые люди могли просто знать, что это один из четырех химических элементов, названных в честь маленького шведского городка Иттерби, наряду с иттербием, эрбием и тербием.
Затем в 1986 году два ученых, работавшие в IBM в Цюрихе, Георг Беднорц и Карл Мюллер, обнаружили, что оксид лантана, бария, меди стал сверхпроводящим при почти рекордно высокой температуре, 35 градусов выше абсолютного нуля.Другими словами, ниже минус 238 ° C электрическое сопротивление соединения исчезло.
Беднорц и Мюллер получили Нобелевскую премию по физике в 1987 году за это открытие. Побуждение других ученых смахнуть пыль со своих Периодических таблиц и попробовать заменить лантановую часть на другие аналогичные металлы. Два американских профессора, Мо-Куен Ву и Пол Чу, вместе со своими исследовательскими группами в университетах Алабамы и Хьюстона изучали оксид иттрия-бария-меди. Он имеет формулу YBa 2 Cu 3 O 7 и часто сокращенно называют YBCO.Они обнаружили, что он стал сверхпроводящим при 95 градусах ниже абсолютного нуля (-178 ºC).
Может показаться, что разница температур невелика, но это означало, что YBCO можно было поддерживать в сверхпроводящем состоянии, используя жидкий азот, а не гораздо более дорогой жидкий гелий. Это послужило поводом для проведения множества исследований за последние 20 лет. Конечная цель, Святой Грааль, состоит в том, чтобы найти материал, который будет сверхпроводником при комнатной температуре, но его еще никто не получил.
Есть много возможных приложений для YBCO; например, сканеры МРТ можно было бы заставить работать дешевле при более высоких температурах с использованием охлаждающей жидкости жидкого азота.Однако в настоящее время существуют технические проблемы, препятствующие использованию этих коммерческих приложений. Один из них заключается в том, что для того, чтобы иметь сверхпроводимость при 95 К, YBCO должен быть немного дефицитным по кислороду, чтобы иметь чуть меньше семи атомов кислорода на атом иттрия. Точная сумма имеет решающее значение, и ее сложно достичь.
Другие проблемы включают приведение YBCO в правильное состояние; В настоящее время проводится множество исследований по созданию тонких пленок из него и поиску способа превращения его в непрерывный провод, а не просто сборку кристаллов, собранных вместе, которые не могут проводить приличные токи.Следователи изучают возможность размещения YBCO поверх гибких металлических проводов, и исследования по этому поводу продолжаются.
Помимо этого, существует множество повседневных применений соединений иттрия. В своих соединениях иттрий всегда присутствует в виде трех плюс-иона иттрия, что означает, что он бесцветен и не имеет неспаренных электронов; поэтому он не обладает собственными интересными магнитными или спектроскопическими свойствами. Положительной стороной этого является то, что соединения иттрия являются очень хорошими материалами-хозяевами для других лантаноидов.
Наиболее известное применение – красный люминофор в электронно-лучевых трубках, который используется в традиционных цветных телевизорах. Он состоит из оксисульфида иттрия Y2O2S, содержащего небольшое количество ионов трехвалентного европия. Точно так же хозяева иттрия часто используются для размещения ионов тербия, которые представляют собой зеленый люминофор. Такие материалы используются в люминесцентных лампах «холодного белого цвета».
Иттрий-алюминиевый гранат, также известный как YAG, является очень важным синтетическим минералом. Из него делают твердые искусственные бриллианты, которые сверкают, как настоящие.Более того, вводя небольшие количества ионов лантаноидов, можно получить материалы с рядом полезных свойств. Например, введите небольшое количество церия, и вы получите хороший желтый люминофор. Или добавьте 1% неодима к YAG, и вы получите наиболее широко используемый твердотельный лазерный материал. А эрбий дает инфракрасный лазер.
Иттрий также находит применение в топливных элементах для питания автомобилей и автобусов, компьютеров и цифровых телефонов и, возможно, зданий. Небольшое количество оксида иттрия добавляется к оксиду циркония для получения так называемого оксида циркония, стабилизированного оксидом иттрия (также называемого YSZ).Он обладает необычным свойством проводить ионы оксидов, что делает его очень полезным в этих топливных элементах. YSZ также используется для установки лямбда-зондов в выхлопную систему вашего автомобиля. Они контролируют количество кислорода в выхлопных газах и отправляют обратную связь, чтобы обеспечить лучшую топливовоздушную смесь в двигателе.
Итак, это иттрий для вас. Бесцветный, невзрачный, но, несомненно, исполняющий множество важных второстепенных ролей.
Мира Сентилингам
Итак, Оскар за лучшую роль второго плана достается, как вы уже догадались, Иттриуму.Это был Саймон Коттон из Аппингемской школы с множеством ролей и применений иттрия. На следующей неделе у нас есть элемент, который может перенести нас в другое измерение.
Хейли Берч
В 1949 году Милтон Смит опубликовал небольшой художественный труд под названием «Тайна элемента 117». Настоящий элемент 117 еще предстоит открыть – это пустое место в Периодической таблице сразу под галогенами. . Смит 117, однако, был странным материалом, который можно было использовать, чтобы открыть окно в другое измерение.Он назвал это магнитным монопольным веществом – веществом, которое вместо полюсов во множественном числе, как у обычного магнита, имело полюс. Единственное число. Теперь, хотя ни один уважаемый ученый не станет утверждать, что магнитный монополь может открыть межпространственный портал, его существование не выходит за рамки возможного, и, если недавние сообщения и есть что-либо, это может зависеть от в противном случае приземленного металлического элемента, который вы можете найти прячущихся около нижней части таблицы Менделеева – гольмий.
Мира Сентилингам
А Хейли Берч раскроет правду о таких мифических монополях в «Химии в ее элементе» на следующей неделе.А пока я Мира Сентилингам, и спасибо за внимание.
(промо)
(конец промо)
Иттрий | химический элемент | Britannica
Иттрий (Y) , химический элемент, редкоземельный металл группы 3 периодической таблицы.
Иттрий – серебристо-белый, умеренно мягкий, пластичный металл. Он довольно устойчив на воздухе; быстрое окисление начинается примерно при температуре выше 450 ° C (840 ° F), в результате получается Y 2 O 3 .Металл легко реагирует с разбавленными кислотами, за исключением плавиковой кислоты (HF), в которой нерастворимый защитный слой YF 3 , образующийся на поверхности металла, предотвращает дальнейшую реакцию. Иттриевые стружки легко воспламеняются на воздухе, раскаленные добела. Металл является парамагнитным с независимой от температуры магнитной восприимчивостью от 10 до 300 К (от -263 до 27 ° C или от -442 до 80 ° F). Он становится сверхпроводящим при 1,3 К (-271,9 ° C или -457 ° F) при давлении, превышающем 110 килобар.
Британская викторина
118 Названия и символы из таблицы Менделеева
Периодическая таблица Менделеева состоит из 118 элементов.Насколько хорошо вы знаете их символы? В этой викторине вам будут показаны все 118 химических символов, и вам нужно будет выбрать название химического элемента, который представляет каждый из них.
В 1794 году финский химик Йохан Гадолин выделил оксид иттрия, новую землю или оксид металла, из минерала, найденного в Иттерби, Швеция. Иттрий, первый открытый редкоземельный элемент, оказался смесью оксидов, из которой за более чем столетний период образовались девять элементов – иттрий, скандий (атомный номер 21) и тяжелые лантаноиды из тербия ( атомный номер 65) до лютеция (атомный номер 71) – были разделены.Иттрий встречается особенно в тяжелых редкоземельных рудах, из которых наиболее важными являются латеритные глины, гадолинит, эвксенит и ксенотим. В магматических породах земной коры этого элемента больше, чем любого из других редкоземельных элементов, за исключением церия, и в два раза больше, чем у свинца. Иттрий также присутствует в продуктах ядерного деления.
Стабильный иттрий-89 – единственный изотоп природного происхождения. Всего 33 (без ядерных изомеров) радиоактивных изотопов иттрия с массой от 77 до 109 и периодом полураспада от 41 миллисекунды (иттрий-108) до 106.Сообщалось о 63 днях (иттрий-88).
Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту.
Подпишитесь сейчас
В промышленных масштабах иттрий отделяют от других редкоземельных элементов методом жидкость-жидкость или ионообменной экстракцией, а металл получают металлотермическим восстановлением безводного фторида кальцием. Иттрий существует в двух аллотропных (структурных) формах. Α-фаза является плотноупакованной гексагональной с a = 3,6482 Å и c = 5,7318 Å при комнатной температуре.Β-фаза является объемноцентрированной кубической с a = 4,10 Å при 1478 ° C (2692 ° F).
Иттрий и его соединения имеют множество применений. Основные области применения включают хосты для красных люминофоров для люминесцентных ламп, цветных дисплеев и экранов телевизоров, в которых используются электронно-лучевые трубки. В лазерах используется иттрий-алюминиевый гранат (АИГ), легированный другими редкоземельными элементами; железо-иттриевый гранат (ЖИГ) используется для микроволновых фильтров, радаров, средств связи и синтетических драгоценных камней; а стабилизированный оксидом иттрия кубический диоксид циркония используется в датчиках кислорода, конструкционной керамике, термобарьерных покрытиях и синтетических алмазах.В основном иттрий используется в высокотемпературной сверхпроводящей керамике, такой как YBa 2 Cu 3 O 7 , которая имеет температуру сверхпроводящего перехода 93 K (-180 ° C или -292 ° F) для Линии электропередачи и сверхпроводящие магниты. Этот металл используется в качестве легирующей добавки к черным и цветным сплавам для повышения коррозионной стойкости и стойкости к окислению. Соединения иттрия используются в оптических стеклах и в качестве катализаторов.
Иттрий химически ведет себя как типичный редкоземельный элемент со степенью окисления +3.Его ионный радиус близок к радиусам диспрозия и гольмия, что затрудняет отделение от этих элементов. Помимо белого полуторного оксида иттрий образует серию почти белых солей, включая сульфат, трихлорид и карбонат.
Свойства элемента
атомный номер
39
атомный вес
88.
точка плавления
1,522 ° C (2,772,33 ° F)
температура кипения
(6053 ° F)
удельный вес
4.469 (24 ° C или 75 ° F)
степень окисления
+3
электронная конфигурация
[Kr] 4 d 1 5 s 2
иттрия [видео] | @GrrlScientist | Наука
Элемент этой недели – иттрий, известный под атомным символом Y и атомным номером 39. Первоначально его атомным символом был Yt, но где-то в начале 1920-х он был заменен на Y.Этот элемент получил свое странное название от деревни Иттерби в Швеции, которая находится недалеко от того места, где этот элемент был обнаружен. Как вы можете видеть на изображении выше, иттрий похож на многие другие элементарные металлы; это серебристо-серый цвет. Он также мягкий, блестящий и очень кристаллический.
Иттрий – это редкоземельный металл, который никогда не встречается в чистом виде в дикой природе. Несмотря на то, что это «редкоземельный металл», он встречается на Земле в 400 раз чаще, чем серебро. Это также довольно часто встречается на Луне.Однако большая часть иттрия в Солнечной системе – это «звездное вещество»; созданы в красных звездах-гигантах.
Этот элемент имеет много общего с лантаноидами и часто встречается в ассоциации с ними, и поэтому часто группируется с ними. Как и все элементы группы III, иттрий имеет нечетный атомный номер, поэтому он имеет только один стабильный изотоп природного происхождения, 89 Y.
Иттрий используется в различных предметах, с которыми вы можете соприкасаться. Чтобы назвать несколько, он используется на электродах высокоэффективных свечей зажигания и в качестве датчика кислорода в автомобилях, он используется для придания красного цвета телевизорам с ЭЛТ (особенно важно, если ваша команда носит красный цвет) и используется для укрепить фианиты – «поддельные бриллианты» – в украшениях.
Биологи в толпе будут разочарованы, узнав, что иттрий не играет известной биологической роли, хотя различные формы этого элемента могут необратимо повредить ваши легкие или печень при вдыхании.
Иттрий действительно применяется в медицине, особенно при лечении рака. Радиоактивный изотоп иттрия-90, период полураспада которого составляет 64 часа, используется для целенаправленного воздействия на различные лимфомы и лейкемии, клетки рака яичников, толстой кишки, поджелудочной железы и костей путем связывания этого изотопа с моноклональными антителами, которые затем связываются непосредственно в раковые клетки.Затем интенсивное излучение распадающегося иттрия-90 убивает связанные клетки.
В этом видео мы посетим шведскую деревню Иттерби зимой, а летом мы также посетим светлую сторону Луны, чтобы рассказать историю иттрия:
Посетите канал периодических видео на YouTube [ссылка на видео]
.. .. .. .. .. .. .. .. .. .. .. ..
Видеожурналист Брэди Харан – человек с фотоаппаратом, а Ноттингемский университет – это место, где живут химики.Вы можете следить за Брэди в твиттере @periodicvideos и за Ноттингемским университетом в твиттере @UniNottingham
Вы уже встречали эти элементы:
Стронций: Sr , атомный номер 38 Рубидий: Rr , атомный номер 37 Криптон: Kr , атомный номер 36 Бром: Br , атомный номер 35227 Селен : Se , атомный номер 34 Мышьяк: As , атомный номер 33 Германий: Ge , атомный номер 32 Галлий: Ga , атомный номер 31 Цинк: Zn , атомный номер 30 Медь: Cu , атомный номер 29 Никель: Ni , атомный номер 28 Кобальт: Co , атомный номер 27 Железо: Fe , атомный номер 26 Марганец: Mn , атомный номер 25 Хром: Cr , атомный номер 24 Ванадий: V , атомарный номер 23 Титан: Ti , атомный номер 22 Скандий: Sc , атомный номер 21 Кальций: Ca , атомный номер 20 Калий: K , атомный номер 19 Аргон: Ar , атомный номер 18 Хлор: Cl , атомный номер 17 Сера: S , атомный номер 16 Фосфор: P , атомный номер 15 Кремний: Si , атомный номер 14 Алюминий: Al , атомный номер 13 Магний: Mg , атомный номер 12 Натрий: Na , атомный номер 11 Неон : Ne , атомный номер 10 Фтор: F , атомный номер 9 Кислород: O , атомный номер 8 Азот: 9 0007 N , атомный номер 7 Углерод: C , атомный номер 6 Бор: B , атомный номер 5 Бериллий: Be , атомный номер 4 Литий: Li , атомный номер 3 Гелий: He , атомный номер 2 Водород: H , атомный номер 1
Вот замечательная интерактивная Периодическая таблица элементов, с которой на самом деле действительно интересно играть с !
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно.Ниже приведены наиболее частые причины:
В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.
Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
Дата на вашем компьютере в прошлом.Если часы вашего компьютера показывают дату до 1 января 1970 г.,
браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.
Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу.Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Измерение и интерпретация экспрессии мобильных элементов
1.
Chénais, B., Caruso, A., Hiard, S. & Casse, N. Влияние мобильных элементов на геномы эукариот: от увеличения размера генома до генетической адаптации к стрессовой среде . Ген 509 , 7–15 (2012).
PubMed
Google ученый
2.
Лиш, Д. Насколько важны транспозоны для эволюции растений? Nat. Преподобный Жене. 14 , 49–61 (2013).
CAS
PubMed
Google ученый
3.
Фолкнер, Дж. Дж. И Гарсиа-Перес, Дж. Л. L1 Мозаицизм у млекопитающих: масштабы, эффекты и эволюция. Trends Genet. 33 , 802–816 (2017).
CAS
PubMed
Google ученый
4.
Чыонг, Э. Б., Элде, Н. К. и Фешотт, К. Регулирующая деятельность сменных элементов: от конфликтов к выгодам. Nat. Преподобный Жене. 18 , 71–86 (2017).
CAS
PubMed
Google ученый
5.
Пайер, Л. М. и Бернс, К. Х. Мобильные элементы в генетических заболеваниях человека. Nat. Преподобный Жене. 20 , 760–772 (2019).
CAS
PubMed
Google ученый
6.
Там, О. Х., Остров, Л. В., Гейл Хэммелл, М. Заболевания нервной системы: активность ретротранспозона при нейродегенеративных заболеваниях. Моб. ДНК 10 , 32 (2019).
PubMed
PubMed Central
Google ученый
7.
Сотеро-Кайо, К.Г., Платт, Р. Н. II, Сух, А. и Рэй, Д. А. Эволюция и разнообразие мобильных элементов в геномах позвоночных. Genome Biol. Evol. 9 , 161–177 (2017).
CAS
PubMed
PubMed Central
Google ученый
8.
Чо, Дж. И Пашковски, Дж. Регулирование развития корней риса ретротранспозоном, действующим как губка микроРНК. eLife 6 , 796 (2017).
Google ученый
9.
Brattås, P. L. et al. TRIM28 контролирует регуляторную сеть генов, основанную на эндогенных ретровирусах в клетках-предшественниках нейронов человека. Cell Rep. 18 , 1–11 (2017).
PubMed
Google ученый
10.
Петри Р. и др. Мобильные элементы LINE-2 являются источником функциональных микроРНК человека и сайтов-мишеней. PLoS Genet. 15 , e1008036 (2019).
CAS
PubMed
PubMed Central
Google ученый
11.
Кашкуш К., Фельдман М. и Леви А. А. Активация транскрипции ретротранспозонов изменяет экспрессию соседних генов в пшенице. Nat. Genet. 33 , 102–106 (2003).
CAS
PubMed
Google ученый
12.
Percharde, M. et al. Партнерство LINE1-нуклеолин регулирует раннее развитие и идентичность ESC. Ячейка 174 , 391–405.e19 (2018).
CAS
PubMed
PubMed Central
Google ученый
13.
Conte, C., Dastugue, B. & Vaury, C. Конкуренция промоторов как механизм транскрипционной интерференции, опосредованной ретротранспозонами. EMBO J. 21 , 3908–3916 (2002).
CAS
PubMed
PubMed Central
Google ученый
14.
Jachowicz, J. W. et al. Активация LINE-1 после оплодотворения регулирует глобальную доступность хроматина у ранних эмбрионов мыши. Nat. Genet. 49 , 1502–1510 (2017).
CAS
PubMed
Google ученый
15.
Стетсон, Д. Б., Ко, Дж. С., Хайдманн, Т. и Меджитов, Р. Trex1 предотвращает внутриклеточную инициацию аутоиммунитета. Cell 134 , 587–598 (2008).
CAS
PubMed
PubMed Central
Google ученый
16.
Аравин А.А. и др. Двухцепочечная РНК-опосредованное подавление тандемных повторов генома и мобильных элементов в геноме D.зародышевой линии melanogaster. Curr. Биол. 11 , 1017–1027 (2001).
CAS
PubMed
Google ученый
17.
De Cecco, M. et al. L1 управляет IFN в стареющих клетках и способствует возрастному воспалению. Nature 566 , 73–78 (2019).
PubMed
PubMed Central
Google ученый
18.
Goic, B. et al. РНК-опосредованная интерференция и обратная транскрипция контролируют персистентность РНК-вирусов в модели насекомых Drosophila. Nat. Иммунол. 14 , 396–403 (2013).
CAS
PubMed
Google ученый
19.
Буржуа, Ю. и Буассино, С. О динамике популяции мусора: обзор популяционной геномики мобильных элементов. Гены 10 , 419–423 (2019).
CAS
Google ученый
20.
Хан, Х., Смит, А. и Буассино, С.Молекулярная эволюция и темп амплификации ретротранспозонов LINE-1 человека с момента происхождения приматов. Genome Res. 16 , 78–87 (2006).
CAS
PubMed
PubMed Central
Google ученый
21.
Хуанг, К. Р. Л., Бернс, К. Х. и Боке, Дж. Д. Активная транспозиция в геномах. Annu. Преподобный Жене. 46 , 651–675 (2012).
CAS
PubMed
PubMed Central
Google ученый
22.
Миллс, Р. Э., Беннет, Э. А., Искоу, Р. К. и Девайн, С. Е. Какие мобильные элементы активны в геноме человека? Trends Genet. 23 , 183–191 (2007).
CAS
PubMed
Google ученый
23.
Brouha, B. et al. Горячие L1 составляют основную часть ретротранспозиции в человеческой популяции. Proc. Natl Acad. Sci. США 100 , 5280–5285 (2003).
CAS
PubMed
Google ученый
24.
Beck, C. R. et al. Активность ретротранспозиции LINE-1 в геномах человека. Cell 141 , 1159–1170 (2010).
CAS
PubMed
PubMed Central
Google ученый
25.
Tubio, J. M. C. et al. Обширная трансдукция неповторяющейся ДНК, опосредованная ретротранспозицией L1 в геномах рака. Наука 345 , 1251343 (2014).
PubMed
PubMed Central
Google ученый
26.
Gardner, E. J. et al. Инструмент локатора мобильных элементов (MELT): обнаружение и биология мобильных элементов в масштабе популяции. Genome Res. 27 , 1916–1929 (2017).
CAS
PubMed
PubMed Central
Google ученый
27.
Rodriguez-Martin, B. et al. Пан-рак анализ полных геномов выявляет перестройки драйверов, вызванные ретротранспозицией LINE-1. Nat. Genet. 52 , 306–319 (2020). Tubio et al. (2014), Гарднер и др. (2017) и Родригес-Мартин и др. (2020) идентифицируют элементы-предшественники L1, активные у человека, на основе полногеномного секвенирования с использованием 3′-трансдукций и внутренних SNP в последовательностях L1.
CAS
PubMed
PubMed Central
Google ученый
28.
Дейнингер П. Л., Батцер М. А., Хатчисон К. А. и Эдджелл М. Х. Мастер-гены в повторяющейся амплификации ДНК млекопитающих. Trends Genet. 8 , 307–311 (1992).
CAS
PubMed
Google ученый
29.
Jacobs, F. M. J. et al. Эволюционная гонка вооружений между генами цинковых пальцев KRAB ZNF91 / 93 и ретротранспозонами SVA / L1. Природа 516 , 242–245 (2014).
CAS
PubMed
PubMed Central
Google ученый
30.
Имбо, М., Хеллебоид, П.-Й.& Trono, D. Белки «цинковые пальцы» KRAB вносят свой вклад в эволюцию регуляторных сетей генов. Nature 543 , 550–554 (2017).
CAS
PubMed
Google ученый
31.
Sanchez-Luque, F. J. et al. LINE-1 уклонение от эпигенетической репрессии у людей. Мол. Ячейка 75 , 590–604 (2019).
CAS
PubMed
Google ученый
32.
Буассино, С., Энтезам, А., Янг, Л., Мансон, П. Дж. И Фурано, А. В. История внедрения активного семейства ретротранспозонов L1 у человека. Genome Res. 14 , 1221–1231 (2004).
CAS
PubMed
PubMed Central
Google ученый
33.
Scott, E.C. et al. Горячий ретротранспозон L1 уклоняется от соматической репрессии и вызывает колоректальный рак человека. Genome Res. 26 , 745–755 (2016). Это исследование повторно секвенировало все нереференсные элементы L1 в случае рака толстой кишки, чтобы идентифицировать внутренние диагностические SNP, а затем, какие элементы экспрессируются в образце .
CAS
PubMed
PubMed Central
Google ученый
34.
Chalopin, D., Naville, M., Plard, F., Galiana, D. & Volff, J.-N. Сравнительный анализ мобильных элементов подчеркивает разнообразие и эволюцию мобилом у позвоночных. Genome Biol. Evol. 7 , 567–580 (2015).
CAS
PubMed
PubMed Central
Google ученый
35.
Quadrana, L. et al. Arabidopsis thaliana mobilome и его влияние на видовой уровень. eLife 5 , e15716 (2016).
PubMed
PubMed Central
Google ученый
36.
McCullers, T. J. & Steiniger, M.Переносные элементы у дрозофилы. Моб. Genet. Элем. 7 , 1–18 (2017).
CAS
Google ученый
37.
Витте С. и Пано О. Ретротранспозоны LTR и размер генома цветковых растений: появление модели увеличения / уменьшения. Cytogenet. Genome Res. 110 , 91–107 (2005).
CAS
Google ученый
38.
Хокинс, Дж.С., Пру, С. Р., Рапп, Р. А. и Вендель, Дж. Ф. Быстрая потеря ДНК как противовес расширению генома через пролиферацию ретротранспозонов в растениях. Proc. Natl Acad. Sci. США 106 , 17811–17816 (2009).
CAS
Google ученый
39.
Капуста А., Сух А. и Фешотт С. Динамика эволюции размера генома у птиц и млекопитающих. Proc. Natl Acad. Sci. США 114 , E1460 – E1469 (2017).
CAS
Google ученый
40.
Гернер-Потвин, П. и Бурк, Г. Вычислительные инструменты для разоблачения мобильных элементов. Nat. Преподобный Жене. 19 , 688–704 (2018).
CAS
PubMed
Google ученый
41.
Vendrell-Mir, P. et al. Тест средств обнаружения вставки транспозона с использованием реальных данных. Моб. ДНК 10 , 53 (2019).
CAS
PubMed
PubMed Central
Google ученый
42.
О’Нил, К., Брокс, Д. и Хэммелл, М.Г. Мобильная геномика: инструменты и методы борьбы с транспозонами. Philos. Пер. R. Soc. Лондон. B. Biol. Sci. 375 , 201 (2020).
PubMed
PubMed Central
Google ученый
43.
Sudmant, P.H. et al. Интегрированная карта структурных вариаций в 2 504 геномах человека. Природа 526 , 75–81 (2015).
CAS
PubMed
PubMed Central
Google ученый
44.
Ewing, A. D. & Kazazian, H. H. Высокопроизводительное секвенирование выявляет обширные вариации в человеческом содержании L1 в индивидуальных геномах человека. Genome Res. 20 , 1262–1270 (2010).
CAS
PubMed
PubMed Central
Google ученый
45.
Максакова И.А. и др. Ретровирусные элементы и их хозяева: инсерционный мутагенез в зародышевой линии мыши. PLoS Genet. 2 , e2 (2006).
PubMed
PubMed Central
Google ученый
46.
Zhang, Y., Maksakova, I. A., Gagnier, L., van de Lagemaat, L. N. & Mager, D. L. Полногеномные оценки выявляют чрезвычайно высокие уровни полиморфизма двух активных семейств эндогенных ретровирусных элементов мыши. PLoS Genet. 4 , e1000007 (2008).
PubMed
PubMed Central
Google ученый
47.
Nellåker, C. et al. Геномный ландшафт сформирован путем отбора на мобильных элементах 18 линий мышей. Genome Biol. 13 , R45 (2012).
PubMed
PubMed Central
Google ученый
48.
Ричардсон, С.R. et al. Наследственная ретротранспозиция L1 в первичной зародышевой линии и раннем эмбрионе мыши. Genome Res. 27 , 1395–1405 (2017).
CAS
PubMed
PubMed Central
Google ученый
49.
Carpentier, M.-C. и другие. Ретротранспозиционный ландшафт азиатского риса выявлен по 3000 геномам. Nat. Commun. 10 , 24 (2019).
CAS
PubMed
PubMed Central
Google ученый
50.
Feusier, J. et al. Родословная оценка скорости ретротранспозиции мобильных элементов человека. Genome Res. 29 , 1567–1577 (2019).
CAS
PubMed
PubMed Central
Google ученый
51.
Rech, G.E. et al. Реакция на стресс, поведение и развитие формируются мутациями, вызванными мобильными элементами у дрозофилы. PLoS Genet. 15 , e1007900 (2019).
CAS
PubMed
PubMed Central
Google ученый
52.
Гонсалес, Дж., Карасов, Т. Л., Мессер, П. В. и Петров, Д. А. Полногеномные паттерны адаптации к умеренной среде, связанные с мобильными элементами у дрозофилы. PLoS Genet. 6 , e1000905 (2010).
PubMed
PubMed Central
Google ученый
53.
Payer, L. M. et al. Структурные варианты, вызванные вставками Alu, связаны с риском многих заболеваний человека. Proc.Natl Acad. Sci. США 114 , E3984 – E3992 (2017).
CAS
PubMed
Google ученый
54.
Казазян, Х. Х. Мл. И Моран, Дж. В. Мобильная ДНК в здоровье и болезнях. N. Engl. J. Med. 377 , 361–370 (2017).
CAS
PubMed
PubMed Central
Google ученый
55.
Seleme, M. D. C. et al. Обширные индивидуальные вариации в способности ретротранспозиции L1 вносят вклад в генетическое разнообразие человека. Proc. Natl Acad. Sci. США 103 , 6611–6616 (2006). Seleme et al. (2006) и Sanchez-Luque et al. (2019) показывают, что данный локус L1 может проявлять вариации внутренней последовательности, приводящие к различиям в активности ретротранспозиции между людьми.
CAS
PubMed
Google ученый
56.
Swergold, G.D. Идентификация, характеристика и клеточная специфичность промотора LINE-1 человека. Мол. Клетка. Биол. 10 , 6718–6729 (1990).
CAS
PubMed
PubMed Central
Google ученый
57.
Томпсон, П. Дж., Макфарлан, Т. С. и Лоринц, М. С. Длинные концевые повторы: от паразитарных элементов до строительных блоков репертуара регуляции транскрипции. Мол. Ячейка 62 , 766–776 (2016).
CAS
PubMed
PubMed Central
Google ученый
58.
Мигелл А. Дж., Маркхэм А. Ф. и Робинсон П. А. Последовательности Alu. FEBS Lett. 417 , 1–5 (1997).
CAS
PubMed
Google ученый
59.
Hancks, D. C., Ewing, A. D., Chen, J. E., Tokunaga, K. & Kazazian, H.H. Захват экзонов, опосредованный человеческим ретротранспозоном SVA. Genome Res. 19 , 1983–1991 (2009).
CAS
PubMed
PubMed Central
Google ученый
60.
Хонигман А., Бар-Шира А., Силберберг Х. и Панет А. Получение однородной 3’-концевой РНК вируса лейкемии мышей. J. Virol. 53 , 330–334 (1985).
CAS
PubMed
PubMed Central
Google ученый
61.
Домброски, Б. А., Матиас, С. Л., Нантакумар, Э., Скотт, А. Ф. и Казазиан, Х. Х. Изоляция активного человеческого подвижного элемента. Science 254 , 1805–1808 (1991).
CAS
PubMed
Google ученый
62.
Conti, A. et al. Идентификация локусов Alu, транскрибируемых РНК-полимеразой III, путем компьютерного скрининга данных RNA-Seq. Nucleic Acids Res. 43 , 817–835 (2014).
PubMed
PubMed Central
Google ученый
63.
Холмс, С.Э., Домброски, Б.А., Кребс, К.М., Бем, К.Д., Казазян, Х.H. Новый ретротранспортированный элемент L1 человека из локуса LRE2 на хромосоме 1q дает химерную вставку. Nat. Genet. 7 , 143–148 (1994).
CAS
PubMed
Google ученый
64.
Моран, Дж. В., ДеБерардинис, Р. Дж. И Казазиан, Х. Х. Перетасовка экзонов с помощью ретротранспозиции L1. Наука 283 , 1530–1534 (1999).
CAS
PubMed
Google ученый
65.
McKerrow, W. & Fenyö, D. L1EM: инструмент для точной количественной оценки локус-специфической РНК LINE-1. Биоинформатика 544 , 115 (2019).
Google ученый
66.
Пикерал, О. К., Макаловски, В., Богуски, М. С. и Боке, Дж. Д. Частая трансдукция геномной ДНК человека, вызванная ретротранспозицией LINE-1. Genome Res. 10 , 411–415 (2000).
CAS
PubMed
PubMed Central
Google ученый
67.
Goodier, J. L., Ostertag, E. M. и Kazazian, H.H. Трансдукция 3’-фланкирующих последовательностей является обычным явлением при ретротранспозиции L1. Хум. Мол. Genet. 9 , 653–657 (2000).
CAS
PubMed
Google ученый
68.
Lander, E. S. et al. Начальная последовательность и анализ человеческого генома. Nature 409 , 860–921 (2001).
CAS
Google ученый
69.
Evrony, G.D. et al. Анализ секвенирования одного нейрона ретротранспозиции L1 и соматической мутации в головном мозге человека. Cell 151 , 483–496 (2012).
CAS
PubMed
PubMed Central
Google ученый
70.
Damert, A. et al. Группы 5’-трансдуцирующих ретротранспозонов SVA эффективно распространяются по геному человека. Genome Res. 19 , 1992–2008 (2009).
CAS
PubMed
PubMed Central
Google ученый
71.
Eickbush, D. G. & Eickbush, T.H. Ретротранспозоны R2 кодируют саморасщепляющийся рибозим для процессинга из котранскрипта рРНК. Мол. Клетка. Биол. 30 , 3142–3150 (2010).
CAS
PubMed
PubMed Central
Google ученый
72.
Перепелица-Белансио, В. и Дейнингер, П. Усечение РНК преждевременным полиаденилированием ослабляет активность мобильных элементов человека. Nat. Genet. 35 , 363–366 (2003).
CAS
PubMed
Google ученый
73.
Schrom, E.-M., Moschall, R., Schuch, A. & Bodem, J. Регулирование ретровирусного полиаденилирования. Adv. Virus Res. 85 , 1–24 (2013).
CAS
PubMed
Google ученый
74.
Белансио, В. П., Хеджес, Д. Дж. И Дейнингер, П. Сплайсинг РНК LINE-1 и влияние на экспрессию генов млекопитающих. Nucleic Acids Res. 34 , 1512–1521 (2006).
CAS
PubMed
PubMed Central
Google ученый
75.
Teixeira, F. K. et al. ПиРНК-опосредованная регуляция альтернативного сплайсинга транспозонов в соме и зародышевой линии. Природа 552 , 268–272 (2017).
CAS
PubMed
PubMed Central
Google ученый
76.
Кинес, К. Дж., Соколовски, М., ДеХаро, Д. Л., Кристиан, К. М. и Белансио, В. П. Потенциал нестабильности генома, связанный с ретротранспозиционно-некомпетентными локусами L1. Nucleic Acids Res. 42 , 10488–10502 (2014).
CAS
PubMed
PubMed Central
Google ученый
77.
Saha, A. et al. Транс-доминантная форма Gag ограничивает ретротранспозицию Ty1 и обеспечивает контроль количества копий. J. Virol. 89 , 3922–3938 (2015).
CAS
PubMed
PubMed Central
Google ученый
78.
Speek, M. Антисмысловой промотор человеческого ретротранспозона L1 управляет транскрипцией соседних клеточных генов. Мол. Клетка. Биол. 21 , 1973–1985 (2001).
CAS
PubMed
PubMed Central
Google ученый
79.
Cruickshanks, H. A. & Tufarelli, C. Выделение специфичных для рака химерных транскриптов, индуцированных гипометилированием антисмыслового промотора LINE-1. Genomics 94 , 397–406 (2009).
CAS
PubMed
Google ученый
80.
Weber, B., Kimhi, S., Howard, G., Eden, A. & Lyko, F. Деметилирование антисмыслового промотора LINE-1 в локусе cMet нарушает передачу сигналов Met за счет индукции незаконной транскрипции . Онкоген 29 , 5775–5784 (2010).
CAS
PubMed
Google ученый
81.
Li, J. et al. Антисмысловой промотор в открытой рамке считывания-1 ретротранспозона L1 мыши инициирует экспрессию различных слитых транскриптов и ограничивает ретротранспозицию. Nucleic Acids Res. 42 , 4546–4562 (2014).
CAS
PubMed
PubMed Central
Google ученый
82.
Denli, A. M. et al. ORF0, специфичный для приматов, способствует разнообразию, опосредованному ретротранспозоном. Ячейка 163 , 583–593 (2015). Это первое исследование, в котором используются данные масс-спектрометрии в крупном масштабе для идентификации неизвестных химерных белков ТЕ .
CAS
PubMed
Google ученый
83.
Руссо Дж., Харрингтон А. В. и Стейнигер М. Антисмысловая транскрипция ретротранспозонов у дрозофилы: происхождение эндогенных предшественников малых интерферирующих РНК. Генетика 202 , 107–121 (2016).
CAS
PubMed
Google ученый
84.
Harrington, A. W. & Steiniger, M. Биоинформатический анализ смысловой и антисмысловой экспрессии терминальных транспозонов с инвертированными повторами в соматических клетках дрозофилы. FLY 10 , 1–10 (2016).
PubMed
PubMed Central
Google ученый
85.
Zingler, N. et al. Анализ 5′-соединений ретротранспозонов LINE-1 и Alu человека предлагает альтернативную модель для 5′-конца прикрепления, требующего микрогомологического соединения концов. Genome Res. 15 , 780–789 (2005).
CAS
PubMed
PubMed Central
Google ученый
86.
Suzuki, J. et al. Генетические доказательства того, что негомологичный путь репарации концевых соединений участвует в ретротранспозиции LINE. PLoS Genet. 5 , e1000461 (2009).
PubMed
PubMed Central
Google ученый
87.
Larson, P. A. et al. Сплайсинговое образование интегрированного ретротранспонированного элемента (SpIRE) в геноме человека. PLoS Biol. 16 , e2003067 (2018).
PubMed
PubMed Central
Google ученый
88.
Penzkofer, T. et al.L1Base 2 – больше ретротранспозиционно-активных LINE-1, больше геномов млекопитающих. Nucleic Acids Res. 45 , D68 – D73 (2017).
CAS
PubMed
Google ученый
89.
Wirth, T., Glöggler, K., Baumruker, T., Schmidt, M. & Horak, I. Семейство средне повторяющихся последовательностей ДНК в геноме мыши со структурными особенностями одиночных ретровирусных длинных концевых повторов. Proc. Natl Acad. Sci. USA 80 , 3327–3330 (1983).
CAS
PubMed
Google ученый
90.
Магер, Д. Л. и Гудчайлд, Н. Л. Гомологичная рекомбинация между LTR элемента, подобного ретровирусу человека, вызывает делецию 5 т.п.н. у двух братьев и сестер. Am. J. Hum. Genet. 45 , 848–854 (1989).
CAS
PubMed
PubMed Central
Google ученый
91.
Vitte, C. & Panaud, O.Образование соло-LTR посредством неравной гомологичной рекомбинации уравновешивает амплификации ретротранспозонов LTR у риса Oryza sativa L. Mol. Биол. Evol. 20 , 528–540 (2003).
CAS
PubMed
Google ученый
92.
Cossu, R.M. et al. LTR Ретротранспозоны демонстрируют низкие уровни неравномерной рекомбинации и высокие скорости внутриэлементной конверсии генов в геномах крупных растений. Genome Biol. Evol. 9 , 3449–3462 (2017).
CAS
PubMed
PubMed Central
Google ученый
93.
Реболло, Р., Фаривар, С. и Магер, Д. Л. C-GATE – каталог генов, на которые влияют мобильные элементы. Моб. ДНК 3 , 9 (2012).
CAS
PubMed
PubMed Central
Google ученый
94.
Келли Д. и Ринн Дж.Мобильные элементы выявляют специфичный для стволовых клеток класс длинных некодирующих РНК. Genome Biol. 13 , R107 (2012).
PubMed
PubMed Central
Google ученый
95.
Kapusta, A. et al. Мобильные элементы вносят основной вклад в происхождение, диверсификацию и регуляцию длинных некодирующих РНК позвоночных. PLoS Genet. 9 , e1003470 (2013). Kapusta et al. (2013) и Kelley and Rinn (2012) обнаружили, что большая часть lncRNA происходит от TE у позвоночных.
CAS
PubMed
PubMed Central
Google ученый
96.
Lu, X. et al. Ретровирус HERVH представляет собой длинную некодирующую РНК, необходимую для идентификации эмбриональных стволовых клеток человека. Nat. Struct. Мол. Биол. 21 , 423–425 (2014).
CAS
PubMed
Google ученый
97.
Wang, J. et al. Специфическая для приматов эндогенная транскрипция, управляемая ретровирусом, определяет наивно-подобные стволовые клетки. Природа 516 , 405–409 (2014).
CAS
PubMed
Google ученый
98.
Исвак, З., Ван, Дж., Сингх, М., Магер, Д. Л. и Херст, Л. Д. Плюрипотентность и эндогенный ретровирус HERVH: конфликт или интуитивная интуиция? BioEssays 38 , 109–117 (2015).
Google ученый
99.
Deininger, P. et al. Комплексный подход к экспрессии локусов L1. Nucleic Acids Res. 45 , e31 (2017).
PubMed
Google ученый
100.
Navarro, F. C. P. et al. TeXP: Деконволюция эффектов всеобъемлющей и автономной транскрипции мобильных элементов. PLoS Comput. Биол. 15 , e1007293 (2019).
CAS
PubMed
PubMed Central
Google ученый
101.
Дженсен Т.Х., Жакье А. и Либри Д. Работа с повсеместной транскрипцией. Мол. Ячейка 52 , 473–484 (2013).
CAS
PubMed
Google ученый
102.
Ли, Х., Чжан, З. и Краузе, Х. М. Длинные некодирующие РНК и повторяющиеся элементы: мусор или интимные партнеры в эволюции? Trends Genet. 35 , 892–902 (2019).
CAS
PubMed
Google ученый
103.
Ким Т.-К., Хемберг М. и Грей Дж. М. Энхансерные РНК: класс длинных некодирующих РНК, синтезируемых энхансерами. Cold Spring Harb. Перспектива. Биол. 7 , a018622 (2015).
PubMed
PubMed Central
Google ученый
104.
Wassenegger, M., Heimes, S., Riedel, L. & Sänger, H.L. РНК-направленное метилирование de novo геномных последовательностей в растениях. Cell 76 , 567–576 (1994).
CAS
PubMed
Google ученый
105.
Fire, A. et al. Сильная и специфическая генетическая интерференция двухцепочечной РНК у Caenorhabditis elegans. Nature 391 , 806–811 (1998).
CAS
PubMed
PubMed Central
Google ученый
106.
Yang, N. & Kazazian, H.H. Ретротранспозиция L1 подавляется эндогенно кодируемыми малыми интерферирующими РНК в культивируемых клетках человека. Nat. Struct. Мол. Биол. 13 , 763–771 (2006).
CAS
PubMed
Google ученый
107.
Slotkin, R.K. et al. Эпигенетическое репрограммирование и подавление малых РНК мобильных элементов в пыльце. Cell 136 , 1451–1454 (2009).
Google ученый
108.
Heras, S. R. et al. Микропроцессор контролирует активность ретротранспозонов млекопитающих. Nat. Struct. Мол. Биол. 20 , 1173–1181 (2013).
CAS
PubMed
Google ученый
109.
Куэрда-Гил, Д. и Слоткин, Р. К. Неканоническое РНК-направленное метилирование ДНК. Nat. Растения 2 , 567–568 (2016).
Google ученый
110.
van de Lagemaat, L. N., Medstrand, P. & Mager, D. L. Множественные эффекты регулируют паттерны выживания эндогенных ретровирусов в интронах генов человека. Genome Biol. 7 , R86 (2006).
PubMed
PubMed Central
Google ученый
111.
Berrens, R.V. et al. Механизм репрессии на основе эндозиРНК противодействует активации транспозонов во время глобального деметилирования ДНК в эмбриональных стволовых клетках. Стволовая клетка 21 , 694–703.e7 (2017).
Roulois, D. et al. Агенты, деметилирующие ДНК, нацелены на клетки колоректального рака, индуцируя вирусную мимикрию эндогенными транскриптами. Ячейка 162 , 961–973 (2015).
CAS
PubMed
PubMed Central
Google ученый
114.
Chiappinelli, K. B. et al. Ингибирование метилирования ДНК вызывает ответ интерферона при раке через дцРНК, включая эндогенные ретровирусы. Cell 162 , 974–986 (2015).
CAS
PubMed
PubMed Central
Google ученый
115.
Skowronski, J. & Singer, M.F. Экспрессия цитоплазматического транскрипта LINE-1 регулируется в линии клеток тератокарциномы человека. Proc. Natl Acad. Sci. США 82 , 6050–6054 (1985).
CAS
PubMed
Google ученый
116.
Белансио, В. П., Рой-Энгель, А. М., Почампалли, Р. Р. и Дейнингер, П. Соматическая экспрессия элементов LINE-1 в тканях человека. Nucleic Acids Res. 38 , 3909–3922 (2010). Вместе с Deininger et al.(2017), эта работа показывает, что большая часть РНК L1, обнаруживаемая в соматических клетках, не является РНК единичной длины, а представляет собой усеченную РНК L1 или происходит из ко-транскрипции или всепроникающей транскрипции .
CAS
PubMed
PubMed Central
Google ученый
117.
Морильон, А., Бенар, Л., Спрингер, М. и Лесаж, П. Дифференциальные эффекты хроматина и Gcn4 на 50-кратный диапазон экспрессии среди отдельных дрожжевых ретротранспозонов Ty1. Мол. Клетка. Биол. 22 , 2078–2088 (2002).
CAS
PubMed
PubMed Central
Google ученый
118.
Слоткин Р. К. и Мартиенсен Р. Мобильные элементы и эпигенетическая регуляция генома. Nat. Преподобный Жене. 8 , 272–285 (2007).
CAS
PubMed
Google ученый
119.
Писарро, Дж.Г. и Кристофари Г. Посттранскрипционный контроль ретротранспозиции LINE-1 клеточными факторами хозяина в соматических клетках. Фронт. Cell Dev. Биол. 4 , 14 (2016).
PubMed
PubMed Central
Google ученый
120.
Гудье, Дж. Л. Ограничение ретротранспозонов: обзор. Моб. ДНК 7 , 344 (2016).
Google ученый
121.
Schorn, A.J., Gutbrod, M.J., LeBlanc, C. & Martienssen, R. Контроль LTR-ретротранспозона с помощью малых РНК, полученных из тРНК. Cell 170 , 61–71.e11 (2017).
CAS
PubMed
PubMed Central
Google ученый
122.
Hohjoh, H. & Singer, M. F. Цитоплазматические рибонуклеопротеиновые комплексы, содержащие человеческий белок LINE-1 и РНК. EMBO J. 15 , 630–639 (1996).
CAS
PubMed
PubMed Central
Google ученый
123.
Biczysko, W., Pienkowski, M., Solter, D. & Koprowski, H. Вирусные частицы в ранних эмбрионах мыши. J. Natl Cancer Inst. 51 , 1041–1050 (1973).
CAS
PubMed
Google ученый
124.
Kulpa, D. A. & Moran, J. V. Образование частиц рибонуклеопротеина необходимо, но не достаточно для ретротранспозиции LINE-1. Хум. Мол. Genet. 14 , 3237–3248 (2005).
CAS
PubMed
Google ученый
125.
Grow, E. J. et al. Внутренняя реактивация ретровирусов в преимплантационных эмбрионах и плюрипотентных клетках человека. Природа 522 , 221–225 (2015).
CAS
PubMed
PubMed Central
Google ученый
126.
Seifarth, W. et al. Комплексный анализ транскрипционной активности эндогенного ретровируса человека в тканях человека с помощью специфичного для ретровируса микроматрицы. J. Virol. 79 , 341–352 (2005).
CAS
PubMed
PubMed Central
Google ученый
127.
Picault, N. et al. Идентификация активного ретротранспозона LTR у риса. Завод. J. 58 , 754–765 (2009).
CAS
PubMed
Google ученый
128.
Horard, B. et al. Глобальный анализ метилирования ДНК и транскрипции повторяющихся последовательностей человека. Эпигенетика 4 , 339–350 (2009).
CAS
PubMed
Google ученый
129.
Reichmann, J. et al. Анализ микроматрицы молчания ретротранспозона LTR идентифицирует Hdac1 как регулятор экспрессии ретротранспозона в эмбриональных стволовых клетках мыши. PLoS Comput. Биол. 8 , e1002486 (2012).
CAS
PubMed
PubMed Central
Google ученый
130.
Гнанаккан, В.P. et al. TE-array – высокопроизводительный инструмент для изучения транскрипции транспозонов. BMC Genomics 14 , 869 (2013).
PubMed
PubMed Central
Google ученый
131.
Faulkner, G.J. et al. Стратегия спасения для множественных отображений меток с короткой последовательностью уточняет исследования транскрипционной активности с помощью CAGE. Genomics 91 , 281–288 (2008).
CAS
PubMed
Google ученый
132.
Chung, N. et al. Транскриптомный анализ соматических тканей, прилегающих к опухоли, выявляет гены, коэкспрессированные с мобильными элементами. Моб. ДНК 10 , 15 (2019). Chung et al. (2019) и McKerrow and Fenyö (2019) предлагают стратегии, основанные на алгоритме EM, для различения и количественной оценки типов транскриптов TE .
Google ученый
133.
Секстон, К. Э. и Хан, М. В. Картирование парных концов мобильных элементов в геноме человека. Моб. ДНК 10 , 29 (2019).
PubMed
PubMed Central
Google ученый
134.
Teissandier, A., Servant, N., Barillot, E. & Bourc’his, D. Инструменты и передовые методы анализа ретротранспозонов с использованием данных высокопроизводительного секвенирования. Моб. ДНК 10 , 52 (2019).
PubMed
PubMed Central
Google ученый
135.
Бао, В., Кодзима, К. К. и Кохани, О. Обновление Repbase, база данных повторяющихся элементов в геномах эукариот. Моб. ДНК 6 , 11 (2015).
PubMed
PubMed Central
Google ученый
136.
Lerat, E., Fablet, M., Modolo, L., Lopez-Maestre, H. & Vieira, C. TEtools облегчает анализ выражений больших данных для мобильных элементов и выявляет антагонизм между их активностью и активностью. генов пиРНК. Nucleic Acids Res. 45 , 1–12 (2017).
Google ученый
137.
Romero-Soriano, V. et al. Неправильная регуляция мобильных элементов связана с расхождением между родительскими путями piRNA у гибридов Drosophila. Genome Biol. Evol. 9 , 1450–1470 (2017).
CAS
PubMed
PubMed Central
Google ученый
138.
Zeng, Z. et al. Полногеномное метилирование ДНК и транскриптомные профили в стратегиях образа жизни и бесполом развитии лесного грибного патогена Heterobasidion parviporum. Эпигенетика 14 , 16–40 (2019).
PubMed
PubMed Central
Google ученый
139.
Song, H. et al. Быстрая эволюция пути piRNA и его транспозонов-мишеней у японской камбалы (Paralichthys olivaceus). Комп.Biochem. Physiol. Часть. D Genomics Proteom. 31 , 100609 (2019).
CAS
Google ученый
140.
Langmead, B. & Salzberg, S. L. Быстрое выравнивание по пробелам и чтению с Bowtie 2. Nat. Методы 9 , 357–359 (2012).
CAS
PubMed
PubMed Central
Google ученый
141.
Li, H. & Durbin, R. Быстрое и точное согласование коротких считываний с помощью преобразования Барроуза-Уиллера. Биоинформатика 25 , 1754–1760 (2009).
CAS
PubMed
PubMed Central
Google ученый
142.
Трапнелл, К., Пахтер, Л. и Зальцберг, С. Л. TopHat: обнаружение сплайсинговых соединений с помощью RNA-Seq. Биоинформатика 25 , 1105–1111 (2009).
CAS
PubMed
PubMed Central
Google ученый
143.
Dobin, A. et al. STAR – сверхбыстрый универсальный выравниватель RNA-seq. Биоинформатика 29 , 15–21 (2013).
CAS
PubMed
PubMed Central
Google ученый
144.
Criscione, S. W., Zhang, Y., Thompson, W., Sedivy, J. M. & Neretti, N. Транскрипционный ландшафт повторяющихся элементов в нормальных и раковых клетках человека. BMC Genomics 15 , 583 (2014).
PubMed
PubMed Central
Google ученый
145.
Yang, W. R., Ardeljan, D., Pacyna, C. N., Payer, L. M. и Burns, K. H. SQuIRE обнаруживает локус-специфическую регуляцию экспрессии вкрапленных повторов. Nucleic Acids Res. 47 , e27 (2019).
CAS
PubMed
PubMed Central
Google ученый
146.
Вальдебенито-Матурана, Б. и Риади, Г. TEcandidates: предсказание геномного происхождения экспрессируемых мобильных элементов с использованием данных РНК-seq. Биоинформатика 34 , 3915–3916 (2018).
CAS
PubMed
Google ученый
147.
Li, B. & Dewey, C. N. RSEM: точное количественное определение транскриптов на основе данных RNA-Seq с референсным геномом или без него. BMC Bioinforma. 12 , 323 (2011).
CAS
Google ученый
148.
Джин, Ю., Там, О. Х., Паниагуа, Э. и Хэммелл, М. TEtranscripts: пакет для включения мобильных элементов в анализ дифференциальной экспрессии наборов данных RNA-seq. Биоинформатика 31 , 3593–3599 (2015). TEtranscripts – первое приложение EM-алгоритма для анализа TE RNA-seq и один из самых популярных программных пакетов, посвященных этой задаче, с момента его выпуска .
CAS
PubMed
PubMed Central
Google ученый
149.
Bendall, M. L. et al. Телескоп: характеристика ретротранскриптома путем точной оценки выражения транспонируемого элемента. PLoS Comput. Биол. 15 , e1006453 (2019).
CAS
PubMed
PubMed Central
Google ученый
150.
Брей, Н. Л., Пиментел, Х., Мельстед, П. и Пахтер, Л. Почти оптимальная вероятностная количественная оценка последовательности РНК. Nat. Biotechnol. 34 , 525–527 (2016).
CAS
PubMed
Google ученый
151.
Патро, Р., Duggal, G., Love, M. I., Irizarry, R. A. & Kingsford, C. Salmon обеспечивает быструю и достоверную количественную оценку экспрессии транскрипта. Nat. Методы 14 , 417–419 (2017).
CAS
PubMed
PubMed Central
Google ученый
152.
Jeong, H.-H., Yalamanchili, H.K., Guo, C., Shulman, J.M. & Liu, Z. Сверхбыстрый и масштабируемый конвейер количественного анализа для мобильных элементов из данных секвенирования следующего поколения. Pac. Symp. Биокомпьют. 23 , 168–179 (2018).
PubMed
Google ученый
153.
Kong, Y. et al. Экспрессия мобильных элементов в опухолях связана с иммунной инфильтрацией и повышенной антигенностью. Nat. Commun. 10 , 5228 (2019).
PubMed
PubMed Central
Google ученый
154.
Philippe, C.и другие. Активация отдельных экземпляров ретротранспозона L1 ограничена пермиссивными локусами, зависимыми от клеточного типа. eLife 5 , 166 (2016). Это исследование предлагает первую стратегию профилирования экспрессии эталонных и нереференсных элементов L1 на уровне локуса путем интеграции целевого повторного секвенирования элементов L1 (секвенирование ATLAS), данных RNA-seq и данных ChIP-seq .
Google ученый
155.
Ewing, A. D. Обнаружение мобильных элементов из данных последовательности всего генома. Моб. ДНК 6 , 24 (2015).
PubMed
PubMed Central
Google ученый
156.
Мир А.А., Филипп С. и Кристофари Г. euL1db: Европейская база данных вставок ретротранспозона L1HS у людей. Nucleic Acids Res. 43 , D43 – D47 (2015).
CAS
PubMed
PubMed Central
Google ученый
157.
Tokuyama, M. et al. Анализ ERVmap показывает транскрипцию эндогенных ретровирусов человека по всему геному. Proc. Natl Acad. Sci. США 115 , 12565–12572 (2018).
CAS
PubMed
PubMed Central
Google ученый
158.
Ансалони, Ф., Скарпато, М., Ди Скьяви, Э., Густинчич, С. и Санжес, Р. Исследовательский анализ экспрессии мобильных элементов в ранних эмбрионах C. elegans. BMC Bioinforma. 20 , 484 (2019).
CAS
Google ученый
159.
Каул Т., Моралес М. Е., Сартор А. О., Белансио В. П. и Дейнингер П. Сравнительный анализ экспрессии локусов L1 с использованием различных препаратов RNA-Seq. Моб. ДНК 11 , 860 (2020).
Google ученый
160.
Faulkner, G.J. et al. Регулируемый транскриптом ретротранспозона клеток млекопитающих. Nat. Genet. 41 , 563–571 (2009). Эта статья предлагает первое общегеномное описание транскрипции TE в нескольких тканях с использованием данных CAGE из проекта PHANTOM .
CAS
PubMed
PubMed Central
Google ученый
161.
Brocks, D. et al. Ингибиторы DNMT и HDAC индуцируют скрытые стартовые сайты транскрипции, кодируемые длинными концевыми повторами. Nat. Genet. 49 , 1052–1060 (2017). Brocks et al. (2017), Roulois et al. (2015) и Chiappinelli et al. (2015) раскрывают механизмы, с помощью которых реактивация ТЕ лекарствами, нацеленными на эпигенетические пути, может убивать раковые клетки.
CAS
PubMed
PubMed Central
Google ученый
162.
Batut, P., Dobin, A., Plessy, C., Carninci, P. & Gingeras, T. R. Профилирование промотора с высокой точностью показывает широко распространенное использование альтернативных промоторов и управляемую транспозонами экспрессию генов развития. Genome Res. 23 , 169–180 (2013).
CAS
PubMed
PubMed Central
Google ученый
163.
Rangwala, S.H., Zhang, L. & Kazazian, H.H. Многие элементы LINE1 вносят вклад в транскриптом соматических клеток человека. Genome Biol. 10 , R100 (2009 г.).
PubMed
PubMed Central
Google ученый
164.
Macia, A. et al. Эпигенетический контроль экспрессии ретротранспозона в эмбриональных стволовых клетках человека. Мол. Клетка. Биол. 31 , 300–316 (2011).
CAS
PubMed
Google ученый
165.
Lock, F. E. et al. Отчетливая изоформа FABP7, выявленная путем скрининга генов, активируемых ретроэлементами, в диффузной крупноклеточной B-клеточной лимфоме. Proc. Natl Acad. Sci. США 111 , E3534 – E3543 (2014).
CAS
PubMed
Google ученый
166.
Морган, Х. Д., Сазерленд, Х. Г., Мартин, Д. И. и Уайтлоу, Е. Эпигенетическое наследование в локусе агути у мышей. Nat. Genet. 23 , 314–318 (1999).
CAS
PubMed
Google ученый
167.
Уилан, С. Дж., Айзава, Ю., Хан, Дж. С. и Боке, Дж. Д. Разрушение генов: новая парадигма эволюции генов, опосредованных ретротранспозоном человека. Genome Res. 15 , 1073–1078 (2005).
CAS
PubMed
PubMed Central
Google ученый
168.
Shen, S. et al. Широкое распространение и регуляторное влияние экзонов Alu на гены человека. Proc. Natl Acad. Sci. США 108 , 2837–2842 (2011).
CAS
PubMed
Google ученый
169.
Butelli, E. et al. Ретротранспозоны контролируют специфическое для фруктов, зависимое от холода накопление антоцианов в кровяных апельсинах. Завод. Ячейка 24 , 1242–1255 (2012).
CAS
PubMed
PubMed Central
Google ученый
170.
Ong-Abdullah, M. et al. Потеря метилирования транспозона кармы лежит в основе скрытого сомаклонального варианта масличной пальмы. Природа 525 , 533–537 (2015).
CAS
PubMed
PubMed Central
Google ученый
171.
Barau, J. et al. ДНК-метилтрансфераза DNMT3C защищает мужские половые клетки от транспозонной активности. Наука 354 , 909–912 (2016).
CAS
PubMed
Google ученый
172.
Attig, J. et al. Экспансия ретроэлемента LTR транскриптома рака человека и иммунопептидома, выявленная сборкой транскриптов de novo. Genome Res. 29 , 1578–1590 (2019).
CAS
PubMed
PubMed Central
Google ученый
173.
Jang, H. S. et al. Мобильные элементы приводят к широко распространенной экспрессии онкогенов при раке человека. Nat. Genet. 51 , 611–617 (2019). Attig et al. (2019) и Jang et al. (2019) предоставляет систематический обзор опухолеспецифических транскриптов и антигенов, полученных из TE .
CAS
PubMed
PubMed Central
Google ученый
174.
Нигуман, П., Редик, К., Мятлик, К. и Спик, М.Многие гены человека транскрибируются с антисмыслового промотора ретротранспозона L1. Genomics 79 , 628–634 (2002).
CAS
PubMed
Google ученый
175.
Peaston, A. E. et al. Ретротранспозоны регулируют гены-хозяева в ооцитах мышей и доимплантационных эмбрионах. Dev. Ячейка 7 , 597–606 (2004).
CAS
PubMed
Google ученый
176.
Липатов, М., Ленков, К., Петров, Д. А., Бергман, К. М. Нехватка транскриптов химерных генов-транспонируемых элементов в геноме Drosophila melanogaster. BMC Biol. 3 , 24 (2005).
PubMed
PubMed Central
Google ученый
177.
Ha, H.-S. и другие. Идентификация и характеристика химерных транскриптов, опосредованных мобильными элементами, из баз данных Refseq и EST свиней. Гены.Геном. 34 , 409–414 (2012).
CAS
Google ученый
178.
Criscione, S. W. et al. Полногеномная характеристика транскриптов, управляемых антисмысловым промотором L1 человека. BMC Genomics 17 , 463 (2016).
PubMed
PubMed Central
Google ученый
179.
Пинсон, М.-Э., Погорельник, Р., Корт, Ф., Арно, П. и Вор-Баррьер, К.CLIFinder: идентификация химерных транскриптов LINE-1 в данных RNA-seq. Биоинформатика 34 , 688–690 (2017).
Google ученый
180.
Бабаян А. и др. LIONS: набор для анализа для обнаружения и количественной оценки транскрипции, инициированной мобильным элементом, с RNA-seq. Биоинформатика 35 , 3839–3841 (2019).
CAS
PubMed
Google ученый
181.
Wang, T. et al. Новая аналитическая стратегия для идентификации транскриптов слияния между повторяющимися элементами и экзонами, кодирующими белок, с использованием RNA-Seq. PLoS One 11 , e0159028 (2016).
PubMed
PubMed Central
Google ученый
182.
Ларроса, Р., Арройо, М., Баутиста, Р., Лопес-Родригес, К. М. и Кларос, М. Г. NearTrans может идентифицировать коррелированные изменения экспрессии между ретротранспозонами и окружающими генами при раке человека. Bioinforma. Биомед. Англ. 10813 , 373–382 (2018).
CAS
Google ученый
183.
Karakülah, G., Arslan, N., Yandin, C. & Suner, A. TEffectR: пакет R для изучения потенциальных эффектов мобильных элементов на экспрессию генов с помощью модели линейной регрессии. PeerJ 7 , e8192 (2019).
PubMed
PubMed Central
Google ученый
184.
Decker, C.J. et al. dsRNA-Seq: идентификация вирусной инфекции путем очистки и секвенирования dsRNA. Вирусы 11 , 943 (2019).
CAS
PubMed Central
Google ученый
185.
Castel, S. E. и Martienssen, R. A. РНК-интерференция в ядре: роли малых РНК в транскрипции, эпигенетике и за ее пределами. Nat. Преподобный Жене. 14 , 100–112 (2013).
CAS
PubMed
PubMed Central
Google ученый
186.
Джонсон, Н. Р., Йео, Дж. М., Корух, К. и Акстелл, М. Дж. Улучшенное размещение множественных малых РНК. G3 6 , 2103–2111 (2016).
CAS
PubMed
Google ученый
187.
Бусиос, А., Гаут, Б. С. и Дарзентас, Н. Соображения и сложности отображения высокопроизводительных данных малой РНК на мобильные элементы. Моб. ДНК 8 , 3 (2017).
PubMed
PubMed Central
Google ученый
188.
Гаррисон, Э. и др. Набор инструментов графа вариаций улучшает отображение считываемых данных, представляя генетические вариации в ссылке. Nat. Biotechnol. 36 , 875–879 (2018).
CAS
PubMed
PubMed Central
Google ученый
189.
Rakocevic, G. et al. Быстрый и точный геномный анализ с использованием геномных графиков. Nat. Genet. 51 , 354–362 (2019).
CAS
PubMed
Google ученый
190.
Ким, Д., Пагги, Дж. М., Парк, К., Беннет, С. и Зальцберг, С. Л. Выравнивание генома на основе графа и генотипирование с HISAT2 и HISAT-генотипом. Nat. Biotechnol. 37 , 907–915 (2019).
CAS
PubMed
PubMed Central
Google ученый
191.
Шерман Р. М. и Зальцберг С. Л. Пангеномика в эпоху генома человека. Nat. Преподобный Жене. 21 , 243–254 (2020).
CAS
PubMed
Google ученый
192.
Maringer, K. et al. Протеомика на основе транскриптомики для характеристики активных мобильных элементов и аннотации генома у Aedes aegypti. BMC Genomics 18 , 101 (2017).
PubMed
PubMed Central
Google ученый
193.
Дэвидсон, А. Д., Мэтьюз, Д. А. и Маринджер, К. Техника протеомики открывает новые горизонты в исследованиях мобиломе. Моб. Genet. Элем. 7 , 1–9 (2017).
Google ученый
194.
Ardeljan, D. et al. Экспрессия LINE-1 ORF2p практически незаметна при раке человека. Моб. ДНК 11 , 1–19 (2019).
PubMed
PubMed Central
Google ученый
195.
Brocks, D., Chomsky, E., Mukamel, Z., Lifshitz, A. & Tanay, A. Анализ отдельных клеток показывает динамику транскрипции мобильных элементов после эпигенетической депрепрессии. BioRxiv https://doi.org/10.1101/462853 (2019).
196.
Шахид С. и Слоткин Р. К. Текущая революция в биологии мобильных элементов, вызванная долгим чтением. Curr. Opin. Genet. Dev. 54 , 49–56 (2020).
CAS
Google ученый
197.
Jiang, F. et al. Длинночитываемое прямое секвенирование РНК с помощью 5’-захвата показывает влияние Piwi на широко распространенную экзонизацию мобильных элементов у саранчовых. RNA Biol. 16 , 950–959 (2019). Это исследование обеспечивает первое использование прямой RNA-seq с помощью технологии Oxford Nanopore для характеристики воздействия транскрипции TE на транскриптом немодельного организма .
PubMed
PubMed Central
Google ученый
198.
Debladis, E., Llauro, C., Carpentier, M.-C., Mirouze, M. & Panaud, O. Обнаружение активных мобильных элементов в Arabidopsis thaliana с использованием технологии секвенирования Oxford Nanopore. BMC Genomics 18 , 537 (2017).
PubMed
PubMed Central
Google ученый
199.
Zhou W. et al. Идентификация и характеристика скрытых специфичных для человека вставок LINE-1 с использованием технологии долгого чтения. Nucleic Acids Res. 409 , 860 (2019).
Google ученый
200.
Wu, T. P. et al. Метилирование ДНК по N6-аденину в эмбриональных стволовых клетках млекопитающих. Природа 532 , 329–333 (2016).
CAS
PubMed
PubMed Central
Google ученый
201.
Simpson, J. T. et al. Обнаружение метилирования цитозина ДНК с помощью секвенирования нанопор. Nat. Методы 14 , 407–410 (2017).
CAS
PubMed
Google ученый
202.
Liu, Q. et al. Обнаружение модификаций оснований ДНК глубокой рекуррентной нейронной сетью на данных секвенирования Oxford Nanopore. Nat. Commun. 10 , 2449 (2019).
PubMed
PubMed Central
Google ученый
203.
Лю, К., Георгиева, Д. К., Эгли, Д. и Ван, К. NanoMod: вычислительный инструмент для обнаружения модификаций ДНК с использованием данных долгочитаемого секвенирования Nanopore. BMC Genomics 20 , 78 (2019).
CAS
PubMed
PubMed Central
Google ученый
204.
Куттер, К., Джерн, П. и Сух, А. Устранение пробелов в исследованиях мобильных элементов с помощью одномолекулярных и одноклеточных технологий. Моб. ДНК 9 , 34 (2018).
CAS
PubMed Central
Google ученый
205.
Солк, Дж. Дж., Шмитт, М. В. и Лоеб, Л. А. Повышение точности секвенирования следующего поколения для обнаружения редких и субклональных мутаций. Nat. Преподобный Жене. 19 , 269–285 (2018).
CAS
PubMed
PubMed Central
Google ученый
206.
Слоткин Р.К. Довод о том, чтобы не скрывать повторяющуюся ДНК. Моб. ДНК 9 , 15 (2018).
PubMed
PubMed Central
Google ученый
207.
Финнеган Д. Дж. Эукариотические мобильные элементы и эволюция генома. Trends Genet. 5 , 103–107 (1989).
CAS
PubMed
Google ученый
208.
Wicker, T. et al. Единая система классификации эукариотических мобильных элементов. Nat. Преподобный Жене. 8 , 973–982 (2007).
CAS
PubMed
Google ученый
209.
Пьегу, Б., Бире, С., Аренсбургер, П. и Биго, Ю. Обзор систем классификации мобильных элементов – призыв к фундаментальному обновлению для решения проблемы их разнообразия и сложности. Мол. Филогенет. Evol. 86 , 90–109 (2015).
PubMed
Google ученый
210.
Курсио, М. Дж. И Дербишир, К. М. Возможности транспозиции: от мю к кенгуру. Nat. Rev. Mol. Cell Biol. 4 , 865–877 (2003).
CAS
PubMed
Google ученый
211.
Hubley, R. et al. База данных повторяющихся семейств ДНК Dfam. Nucleic Acids Res. 44 , D81 – D89 (2016).
CAS
PubMed
Google ученый
212.
Amselem, J. et al. RepetDB: унифицированный ресурс для ссылок на переносимые элементы. Моб. ДНК 10 , 6–8 (2019).
Fadloun, A. et al. Сигнатуры хроматина и профили ретротранспозонов у эмбрионов мышей выявляют регуляцию LINE-1 с помощью РНК. Nat. Struct. Мол. Биол. 20 , 332–338 (2013).
CAS
PubMed
Google ученый
215.
Дерриен, Т.и другие. Быстрые вычисления и приложения сопоставимости генома. PLoS One 7 , e30377 (2012).
CAS
PubMed
PubMed Central
Google ученый
216.
Каримзаде, М., Эрнст, К., Кундаже, А. и Хоффман, М. М. Умап и Бисмап: количественная оценка сопоставимости генома и метилома. Nucleic Acids Res. 46 , e120 (2018).
PubMed
PubMed Central
Google ученый
Механизмы реактивного элемента Y на очистке суперсплава К4169 при вакуумной индукционной плавке
[1]
A.Iturbe, E. Giraud, E. Hormaetxe, A. Garay, G. Germain, K. Ostolaza, P.J. Arrazola, Механическое определение характеристик и моделирование поведения материала Inconel 718 для оценки процесса обработки, Mater. Sci. Англ. А , 682 (2017), стр. 441.
Статья
Google ученый
[2]
D.H. Ping, Y.F. Гу, С.Ю. Цуй и Х. Харада, Сегрегация на границах зерен в суперсплаве на основе Ni-Fe (сплав 718), Mater. Sci. Англ. А , 456 (2007), №1-2, стр. 99.
Статья
Google ученый
[3]
Г.А. Zickler, R. Schnitzer, R. Radis, R. Hochfellner, R. Schweins, M. Stockinger и H. Leitner, Микроструктура и механические свойства суперсплава ATI Allvac® 718Plus ™, Mater. Sci. Англ. А , 523 (2009), № 1-2, с. 295.
Статья
Google ученый
[4]
D.K. Дас, В. Сингх и С.В. Джоши, Высокотемпературное окислительное поведение направленно отвержденного суперсплава на основе никеля CM-247LC, Mater. Sci. Technol. , 19 (2013), №6, с. 695.
Статья
Google ученый
[5]
E.C. Caldwell, F.J. Fela, and G.E. Фукс, Сегрегация элементов в монокристаллических суперсплавах на основе никеля с высоким содержанием тугоплавких материалов, JOM , 56 (2004), № 9, с. 44.
Статья
Google ученый
[6]
A.Дж. Бранд, К. Кархаузен и Р. Копп, Моделирование микроструктуры сплава на основе никеля Incone * 718 при производстве дисков турбин, Mater. Sci. Technol. , 12 (1996), № 11, с. 963.
Статья
Google ученый
[7]
T.M. Поллок и С. Тин, Суперсплавы на основе никеля для перспективных газотурбинных двигателей: химия, микроструктура и свойства, J. Propul. Мощность , 22 (2006), № 2, с. 361.
Статья
Google ученый
[8]
В.Сидоров В. Ригин, П. Мин, Ю.И. Фоломейкин, Удаление примеси серы из расплавов сложного никеля в вакууме, . Металл. , 2015 (2015), № 11, с. 910.
Статья
Google ученый
[9]
Х. Наффах-Моосави, Развитие микроструктуры и прогнозирование литейных свойств в недавно разработанных современных суперсплавах на никелевой основе третьего поколения, Int. Дж. Майнер. Металл. Матер. , 23 (2016), №5, с. 548.
Статья
Google ученый
[10]
Л.Ван, Ю. Ван, Ю. Лю, X. Сун, X.D. Люй, Б.Дж. Чжан, Укрупнение γ ‘и γ ″ фаз в суперсплаве Gh5169 при обработке электрическим полем, Int. Дж. Майнер. Металл. Матер. , 20 (2013), № 9, с. 861.
Статья
Google ученый
[11]
W.X. Ю. и Дж. П. Ниу, Раскисление и денитрогенизация при рафинировании суперсплава на основе никеля методом VIM, New Technol. Новый процесс , 2002, № 3, с. 32.
Google ученый
[12]
С.Ф. Миллер, Г. Симмонс, Р.П. Вей, Механизм роста трещин, вызванных кислородом, в инконеле 718, Scripta Mater. , 44 (2001), № 10, с. 2405.
Статья
Google ученый
[13]
К. Садананда, П. Шагинян, Влияние окружающей среды на характер роста трещин ползучести для некоторых конструкционных сплавов, Mater. Sci. Англ. , 43 (1980), № 2, с. 159.
Статья
Google ученый
[14]
С.Сариоглу, К. Стиннер, Дж.Р. Блачеер, Н. Биркс, Ф.С. Петтит, Г. Мейер, Дж. Л. Смиалек, Контроль содержания серы в монокристаллических суперсплавах на основе никеля и его влияние на стойкость к циклическому окислению, Superalloys , 1996, с. 71.
Google ученый
[15]
T.M. Симпсон и А. Прайс, Улучшение окисления обработанных суперсплавов с низким содержанием серы, Суперсплавы , 2000, стр. 387.
Google ученый
[16]
Дж.X. Dong, X.B. Лю, Б. Тан, Ю. Ху, З.С. Сюй, Х.С. Се, Влияние S на механические свойства и микроструктуру сплава Inconel 718, Acta Metall. Грех. , 32 (1996), № 3, с. 241.
Google ученый
[17]
L.V. Раманатан, Роль редкоземельных элементов в высокотемпературном окислительном поведении сплавов FeCr, NiCr и NiCrAl, Corros. Sci. , 35 (1993), № 5-8, стр. 871.
Статья
Google ученый
[18]
Н.Nayan, · Govind, · C.N. Сайкришна, К. Рамая, С.К. Бхаумик, К. Наир и М. Миттал, Вакуумная индукционная плавка NiTi сплавов с памятью формы в графитовом тигле, Mater. Sci. Англ. А , 465 (2007), № 1-2, с. 44.
Статья
Google ученый
[19]
X.H. Ченг, Л. Фань, Л. Ли, К.Ф. Ду, Д.Х. Ван, Влияние легирования алюминия и иттрия на высокотемпературное окислительное поведение сплава Ni-11Fe-10Cu, J. Rare Earths , 34 (2016), No.11, стр. 1139.
Статья
Google ученый
[20]
X.L. Ли, С. Он, X.T. Чжоу, Ю. Цзоу, З.Дж. Ли, А.Г. Ли и X.H. Ю., Влияние редкоземельного иттрия на микроструктуру и свойства суперсплава на никелевой основе Ni-16Mo-7Cr-4Fe, Mater. Charact. , 95 (2014), с. 171.
Статья
Google ученый
[21]
P.J. Zhou, J.J. Ю., Х.Ф. Сунь, Х.Р. Гуань, Х.M. He и Z.Q. Ху, Влияние Y на свойство разрушения под напряжением суперсплава на основе Ni, Mater. Sci. Англ. А , 551 (2012), стр. 236.
Статья
Google ученый
[22]
L.G. Сонг, С.С. Ли, Ю. Чжэн, Ю.Ф. Хан, Влияние иттрия на стойкость к высокотемпературному окислению направленно отверждаемого суперсплава, J. Rare Earths , 22 (2004), № 6, с. 794.
Google ученый
[23]
H.Б. Бай, Х. Р. Чжан, Дж. Ф. Венг, Б. Конг и Х. Чжан, Поведение при очистке обрезков Gh5169 в атмосфере аргона во время вакуумной индукционной плавки, Mater. Res. Инновации , 18 (2014), № S4, с. 357.
Google ученый
[24]
H.R. Zhang, X.X. Тан, К. Чжоу, Х. Чжан и С.В. Чжан, Сравнение направленной кристаллизации сплавов γ-TiAl в обычном Al 2 O 3 и новом Y 2 O 3 Al с покрытием O 3 тиглей, Дж.Евро. Ceram. Soc. , 33 (2013), № 5, с. 925.
Статья
Google ученый
[25]
S.J. Ли, Ю. Ху, Х.С. Мэй, X.S. Се, Ю. Он и Х. Чжан, Обессеривание суперсплава на основе никеля GH690, J. Iron Steel Res. , 15 (2003), № 7, с. 317.
Google ученый
[26]
L.H. Zhao, X.M. Чжэн и Дж. Fei, Поверхностные свойства твердоосновных катализаторов на основе оксидов редкоземельных элементов.Характеристика поверхностно-активных центров калализаторов оксидов редкоземельных элементов, Chin. J. Catal. , 17 (1996), № 3, с. 227.
Google ученый
[27]
К. Сан, Р.Ф. Хуанг, J.T. Го и Z.Q. Ху, Распределение серы в литом суперсплаве на основе никеля K24 и его влияние на механические свойства, High Temp. Technol. , 6 (1988), № 3, с. 145.
Статья
Google ученый
Элемент.getBoundingClientRect () – веб-API | MDN
Метод Element.getBoundingClientRect () возвращает DOMRect объект, предоставляющий информацию о размере элемента и его
положение относительно области просмотра.
domRect = element.getBoundingClientRect ();
Значение
Возвращаемое значение – объект DOMRect , который является наименьшим прямоугольником.
который содержит весь элемент, включая его отступы и ширину границы.В слева , вверху , справа , внизу , x , y , ширина и высота свойств
описать положение и размер всего прямоугольника в пикселях. Свойства кроме ширина и высота относительно левого верхнего угла области просмотра.
Свойства width и height DOMRect объект, возвращаемый методом, включает заполнение , и border-width , а не только ширина / высота содержимого.В стандартной коробочной модели
это будет равно свойству width или height свойства
element + padding + border-width . Но
if box-sizing: border-box – это
для элемента, это будет прямо равно его ширине или высота .
Возвращаемое значение можно рассматривать как объединение прямоугольников, возвращаемых функцией getClientRects () для элемента, то есть CSS
рамки, связанные с элементом.
Пустые рамки полностью игнорируются. Если все рамки элемента пусты,
затем возвращается прямоугольник с шириной и высотой нуля
и где вверху и слева – это верхний левый угол рамки
для первого блока CSS (в порядке содержимого) для элемента.
Объем прокрутки области просмотра (или любой другой
scrollable element) учитывается при вычислении ограничивающего прямоугольника.Этот
означает, что граничные края прямоугольника ( верхний , правый , снизу , слева ) меняют свои значения каждый раз при прокрутке
изменения позиции (поскольку их значения относятся к области просмотра, а не абсолютны).
Если вам нужен ограничивающий прямоугольник относительно левого верхнего угла документа,
просто добавьте текущую позицию прокрутки вверху и слева properties (их можно получить с помощью окна window.scrollX и window.scrollY ), чтобы получить ограничивающий прямоугольник, не зависящий от
текущая позиция прокрутки.
Кроссбраузерный откат
Скрипты, требующие высокой кроссбраузерности, могут использовать window.pageXOffset и window.pageYOffset вместо window.scrollX и window.scrollY. Скриптов без доступа к
эти свойства могут использовать такой код:
пусть elem = document.querySelector ('div');
let rect = elem.getBoundingClientRect ();
for (var key in rect) {
if (typeof rect [ключ]! == 'функция') {
пусть para = document.createElement ('p');
para.textContent = `$ {ключ}: $ {rect [ключ]}`;
document.body.appendChild (параграф);
}
}
Обратите внимание, как ширина / высота равна его ширина / высота + набивка .
Также обратите внимание, как значения x / оставили , y / верхний , правый и нижний равны
на абсолютное расстояние от соответствующего края области просмотра до этой стороны
элемент, в каждом случае.
Прокрутка
Этот пример демонстрирует, как ограничивающий прямоугольник клиента изменяется при прокрутке документа.
div # example {
ширина: 400 пикселей;
высота: 200 пикселей;
отступ: 20 пикселей;
маржа: 50 пикселей автоматически;
фон: фиолетовый;
}
body {padding-bottom: 1000 пикселей; }
р {маржа: 0; }
function update () {
const контейнер = документ.getElementById («элементы управления»);
const elem = document.getElementById ("пример");
const rect = elem.getBoundingClientRect ();
container.innerHTML = '';
for (введите rect) {
if (typeof rect [ключ]! == 'функция') {
пусть para = document.createElement ('p');
para.textContent = `$ {ключ}: $ {rect [ключ]}`;
container.appendChild (параграф);
}
}
}
document.addEventListener ('прокрутка', обновление);
Обновить();
Банкноты
Возвращенный объект DOMRect можно изменить в современных браузерах.Это было
неверно для более старых версий, которые фактически возвращали DOMRectReadOnly .
В IE и Edge невозможно добавить недостающие свойства к возвращаемому ClientRect ,
объект предотвращает засыпку x и y .
Из-за проблем совместимости (см. Ниже) безопаснее всего полагаться только на свойства слева , вверху , справа и внизу .

/periodic-table--illustration-738787291-59888b4822fa3a00109a466d.jpg) Запатентованный многослойный состав материала обеспечивает наивысший класс сопротивления природным катаклизмам.
Запатентованный многослойный состав материала обеспечивает наивысший класс сопротивления природным катаклизмам.







 элемент Y
элемент Y рабочая температура
рабочая температура 2.3.1 Нагревостойкость изоляции
2.3.1 Нагревостойкость изоляции 2.4 Устойчивость к ультрафиолетовому излучению
2.4 Устойчивость к ультрафиолетовому излучению
 6 Монтаж оборудования
6 Монтаж оборудования
 11 Стойкость к коротким замыканиям
11 Стойкость к коротким замыканиям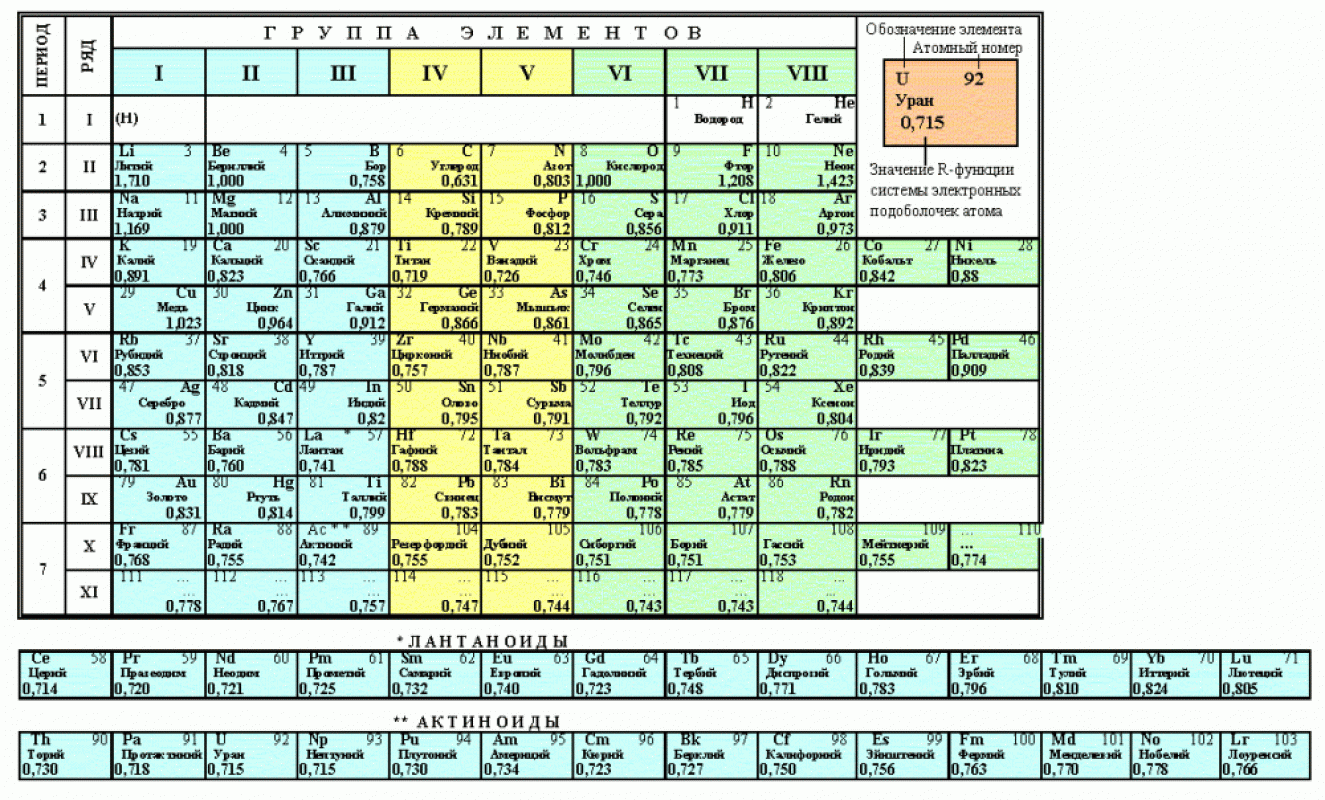 Pashchenko S.,
N. Komarov M.,
Д. Пащенко С. and
Н. Комаров М.
Pashchenko S.,
N. Komarov M.,
Д. Пащенко С. and
Н. Комаров М.
 The article also discusses fiancial reserves usage as a tool for active risk management.
The article also discusses fiancial reserves usage as a tool for active risk management. fa.ru/jour/article/viewFile/304/294.pdf (application/pdf)
fa.ru/jour/article/viewFile/304/294.pdf (application/pdf)

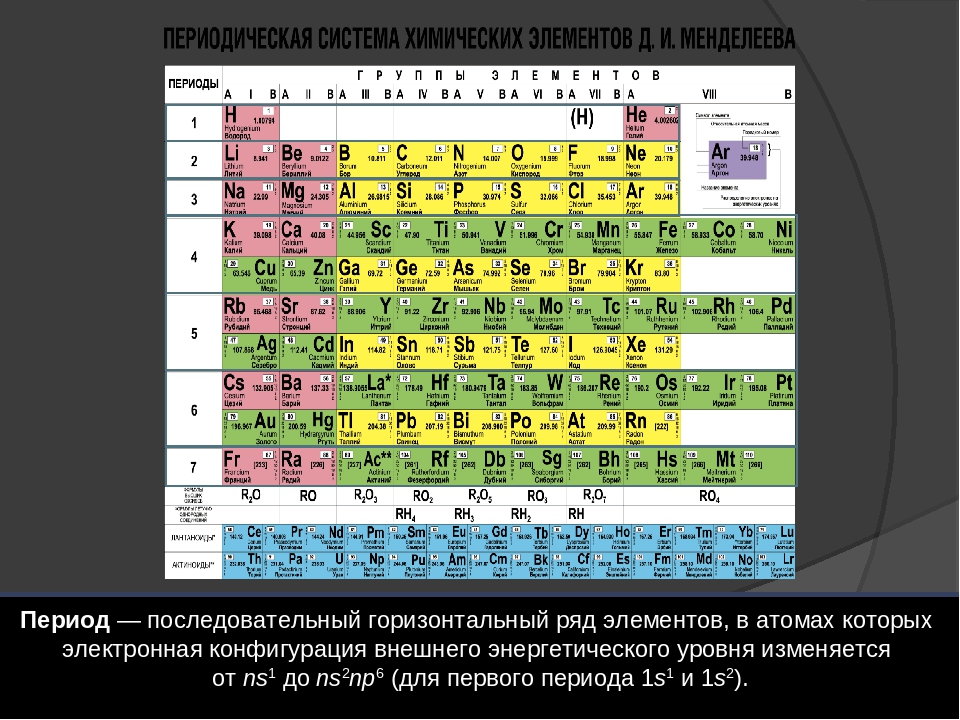 Побуждение других ученых смахнуть пыль со своих Периодических таблиц и попробовать заменить лантановую часть на другие аналогичные металлы. Два американских профессора, Мо-Куен Ву и Пол Чу, вместе со своими исследовательскими группами в университетах Алабамы и Хьюстона изучали оксид иттрия-бария-меди. Он имеет формулу YBa 2 Cu 3 O 7 и часто сокращенно называют YBCO.Они обнаружили, что он стал сверхпроводящим при 95 градусах ниже абсолютного нуля (-178 ºC).
Побуждение других ученых смахнуть пыль со своих Периодических таблиц и попробовать заменить лантановую часть на другие аналогичные металлы. Два американских профессора, Мо-Куен Ву и Пол Чу, вместе со своими исследовательскими группами в университетах Алабамы и Хьюстона изучали оксид иттрия-бария-меди. Он имеет формулу YBa 2 Cu 3 O 7 и часто сокращенно называют YBCO.Они обнаружили, что он стал сверхпроводящим при 95 градусах ниже абсолютного нуля (-178 ºC).