
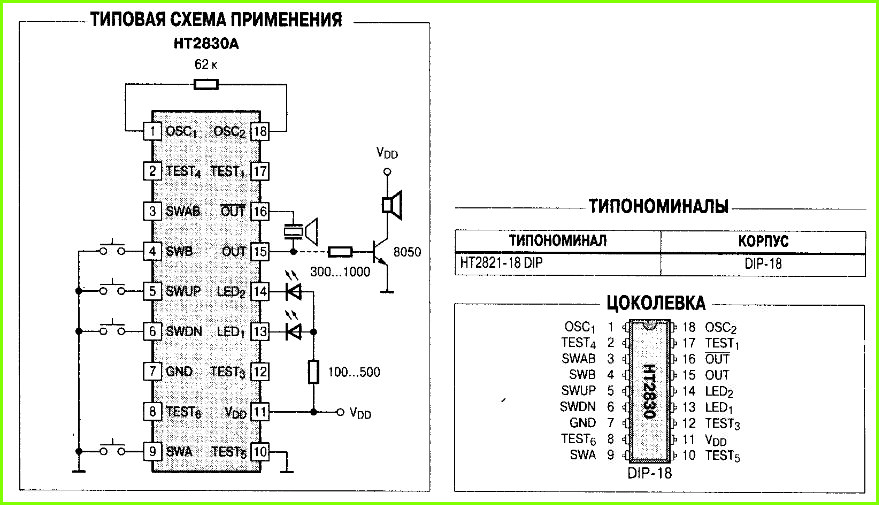
ᐅ Zanussi ZACS-09 HT/N отзывы — 13 честных отзыва покупателей о кондиционере Zanussi ZACS-09 HT/N
Самые выгодные предложения по Zanussi ZACS-09 HT/N
Шуркович Шурк, 11.09.2018
Достоинства:
Тихий, производительный, автоматические вертикальные шторки, много режимов работы, ночной режим действительно позволяет тихо засыпать, соотношение цена-качество-надежность, обогрев до -7, гарантия 3 года.
Недостатки:
Пульт с множеством неактивных кнопок, отсутствие крепления для пульта на стену.
Комментарий:
Порекомендовали сами установщики, вполне достойный аппарат. Рекомендую.
Павел Баренцев, 04.
Недостатки:
нет
Комментарий:
в комнате 22 м2 кондиционер хорошо справляется со своими функциями, работая как на обогрев в холодное время года, так и на охлаждение в жару. Его работа не слышна благодаря шумоизоляции внешнего блока и специальному тихому режиму. Антибактериальный фильтр делает воздух чище и избавляет его от вирусов и аллергенов. В комплекции есть очень удобный и понятный пульт ДУ.
Елена Смирнова, 14.08.2018
Достоинства:
Охлаждает и нагревает быстро. Фильтр очищает воздух от вредных примесей и пыли.
Недостатки:
Много кнопок на пульте, сложно разобраться.
Комментарий:
Кондиционер в любом режиме поддерживает установленную мною температуру, с точностью до 1 гр. Если происходит сбой в электросети, происходит перезапуск системы, с моими настройками. Жалюзи широкоугольные, направление меняю по своему усмотрению, но есть и автоматическое качание, поток воздуха распределяется по вертикали
Анатолий Чкалов, 06.08.2018
Достоинства:
качественная работа.
Недостатки:
нет
Комментарий:
Нравится точность установки температуры до половины градуса. Рабочая температура внешнего блока позволяет использовать систему в межсезонье. А помещение нагревается до нужной температуры за пол часа. Иногда управляю с помощью мобильного приложения, когда не могу найти пульт или забыл заранее поставить таймер.
sf Макар, 14.07.2018
Достоинства:
Симпатичный дизайн, производительный
Недостатки:
нет подсветки в пульте и крепления пульта на стену, датчик температуры только во внутреннем блоке
Комментарий:
Ионизатора в нем нет. В комплекте есть 2 кахетиновых предфильтра, но только на одном есть нера-фильтр, хотя гнезда есть на обоих. В весьма симпатичном пульте нет подсветки. Когда дует ветер (а ветра у нас дуют сильные) во время работы кондея вода в дренажном коллекторе внутреннего блока начинает начинает забавно булькать – сильный подпор воздуха мешает корректному дренажу. Это были минусы. Все минусы! Теперь о плюсах: качественное охлаждение, на комнату в 24 кв. м с высотой потолков 2,7 м мощи предостаточно. Внешний блок не вибрирует, внутренний – тихо шелестит.
Роберт Критченков, 11.06.2018
Достоинства:
Функция запоминания настроек, теплый пуск, дизайн, таймер.
Недостатки:
Пока не обнаружил
Комментарий:
Аппарат установил на даче. Очень быстро делает нужную температуру в помещении, часто использую в режиме автоматической поддержки температуры для экономии электроэнергии. Установлена рабочая система самодиагностики неисправностей и дезодорирующий фильтр. У пульта управления простой интерфейс. Удобная регулировка направления воздушного потока.
Установлена рабочая система самодиагностики неисправностей и дезодорирующий фильтр. У пульта управления простой интерфейс. Удобная регулировка направления воздушного потока.
Aleksej Lisnyakov, 03.06.2018
Достоинства:
светодиодный дисплей на внутреннем блоке.
Недостатки:
недостатков нет
Комментарий:
Мне кондиционер нравится, работает довольно тихо. Пульт пришлось изучить, но там всё понятно. Можно включить автоматический режим. Я выбирал устройство с жалюзи: здесь есть горизонтальные и вертикальные. Всё регулируется с пульта.
Геннадий Платошкин, 04.05.2018
Достоинства:
Цена, функции.
Недостатки:
нет
Комментарий:
Приобретал систему для охлаждения однокомнатной квартиры летом прошлого года, температура изменяется во всем помещении примерно за полчаса. Запускаю в автоматическом режиме, все устраивает. Зимой использовал, естественно, обогрев. Тоже работает ощутимо (очень мощно если на полную). Сделана из хорошего пластика, нет запахов неприятных.
Виталий Афанасьев, 19.04.2018
Достоинства:
современный дизайн, индикация температуры и режимов. Можно регулировать воздушный поток.
Недостатки:
нет
Комментарий:
Установил в офисе на 26 кв.м., включается и выключается по таймеру на заданных параметрах, не нужно заново настраивать. Не портит интерьер, индикация не раздражает глаз. В жару пользуемся быстрым режимом, охлаждает моментально.
В жару пользуемся быстрым режимом, охлаждает моментально.
Марк Чеканов, 14.04.2018
Достоинства:
Большое количество регулировок, точность поддержания температуры, экономичность
Недостатки:
Бывают сбои с настройками
Комментарий:
Пока больше использовал для вентиляции, чем охлаждения или обогрева, но нормально работают все режимы. Дует хорошо, чувствуется метров с трех, даже особенно и охлаждать не надо, максимум что включал пока – вторую мощность. Проверил как температуру держит – действительно, за пределы одного градуса в сторону не выходит. Правда, есть и минус – настройки порой шалят и могут сбиться.
Сергей Бутылин, 13.
Достоинства:
много полезных функций, начиная от фильтрации, дезодорации воздуха до разморозки. Эффективен для помещения до 20 кв. метров. Охлаждение и нагрев четкий, при работе в режиме нагрева включается горячий старт, для исключения выброса холодного воздуха.
Давид Житомирский, 04.09.2013
Достоинства:
Быстрое достижение и четкое поддержание температуры
Недостатки:
нет
Комментарий:
Кондёр работает в гостиной на протяжении всего лета практически без выключения. На максимальной скорости нагнетает 20 гр. минут за 15. На минимальной – дует потихоньку, даже спать не мешает.
Сергей Флёров, 23.07.2013
Достоинства:
Таймер, самодиагностика.
Недостатки:
Нет
Комментарий:
Поставил на даче, отлично холодит и вентилирует в жару. Энергии много не берет, в ненастье обогревает весь дом, ибо можно включать когд легкий морозец (до минус семи) До нужной температуры разгоняется быстро. ПДУ работает четко, через него выставляю все настройки.
HTN – это… Что такое HTN?
HTN — can refer to:* Hypertension a medical condition * Hierarchical task network a planning formalism in artificial intelligence * High Twisted Nematic A type of liquid crystal display with a viewing range from 100 110°. This type of display is not… … Wikipedia
This type of display is not… … Wikipedia
HTN — puede referirse a: Hierarchical Task Network un algorítmo de planificación automática. En medicina es HiperTensióN. High Twisted Nematic Un tipo de liquid crystal display que permite un ranzo azimutal de visión de 100 110°. Hughes Television… … Wikipedia Español
HTN — Hantaan [ like virus]; histatin; hypertension; hypertensive nephropathy … Medical dictionary
HTN — Host Trade Number Sequential number attributed by the host in order to identify a trade. It is attributed per instrument and per ( calendar) day. Chicago Mercantile Exchange Glossary … Financial and business terms
HTN — abbr. med. hypertension – гипертензия, гипертония … Dictionary of English abbreviation
HTN — abbr. Heterodyne … Dictionary of abbreviations
HTN — • Hantaan [ like virus]; • histatin; • hypertension; • hypertensive nephropathy … Dictionary of medical acronyms & abbreviations
Hierarchical task network — In artificial intelligence, the hierarchical task network, or HTN, is an approach to automated planning in which the dependency among actions can be given in the form of networks. Planning problems are specified in the hierarchical task network… … Wikipedia
Planning problems are specified in the hierarchical task network… … Wikipedia
Red jerárquica de tareas — La Red jerárquica de tareas (denominada también del inglés como: Hierarchical task network o abreviadamente HTN) es un algoritmo de planificación automática que crea un plan por descomposición de tareas en subtareas hasta lograr primitivas que… … Wikipedia Español
List of the busiest airports in the People’s Republic of China — Beijing Capital International Airport, the busiest airport by passenger traffic in the People s Republic of China and the second busiest airport in the world. China s busiest airports are a series of lists ranking the one hundred busiest airports … Wikipedia
Zytel — is a trademark owned by DuPont and used for a number of different high strength, abrasion and impact resistant thermoplastic polyamide formulations of the family more commonly known as nylon, often with varying degrees of fiberglass, from 13% to… … Wikipedia
|
Компрессор: Тип: герметичный Марка: Tecumseh Модель: CAE9470Z Мощность: 5/8 ЛС |
Конденсатор Шаг ребер: 2,1 мм Площадь поверхности: 4,9 м2 Количество вентиляторов: 1, диаметр 350 мм Мощность двигателей: 65 Вт Производительность: 2120 м3/ч |
|
Воздухоохладитель: Тип оттайки: ТЭН Медные трубки с алюминиевыми ребрами Шаг ребер: 4,2/8,4 мм Площадь поверхности: 4,8 м2 Количество вентиляторов 1 диаметром 254 мм Мощность двигателей 1х70 Вт Производительность: 810 м3/ч Длина струи: 6 м |
Электрические кабели: Внешний 3 x 1,5 мм2 Силовой межблочный 4 x 1,5 мм2 Пульта дистанционного управления 2 x 1,0 мм2
На датчики 4 x 0,75 экран. |
|
Вес и габариты:
Компрессорно-конденсаторная часть
Масса нетто 66 кг, масса брутто: 70 кг
Габариты упаковки: 505x1215x685 (h) мм
Воздухоохладитель
Масса нетто 9 кг, масса брутто: 22 кг Габариты упаковки: 545x690x360 (h) мм |
 экр
экр
Артериальная гипертензия
Целью исследования была оценка возможной ассоциации межвизитной вариабельности артериального давления (АД) и риска неблагоприятных исходов у больных артериальной гипертензией (АГ) после эпизода острого коронарного синдрома.
Материалы и методы. В статье проанализированы данные о 1456 больных (средний возраст 65,6 ± 12,17 года, 875 (60,1 %) мужчин), выписанных из стационара после эпизода острого коронарного синдрома и наблюдавшихся в течение 1 года в 4 сосудистых центрах Москвы, Астрахани, Казани и Краснодара в 2014–2017 годах. При выписке больных и на 25-й, 90-й, 180-й и 360-й день после выписки регистрировали АД, частоту сердечных сокращений и неблагоприятные события. О межвизитной вариабельности АД судили по коэффициенту VIM (variation independent of mean).
В статье проанализированы данные о 1456 больных (средний возраст 65,6 ± 12,17 года, 875 (60,1 %) мужчин), выписанных из стационара после эпизода острого коронарного синдрома и наблюдавшихся в течение 1 года в 4 сосудистых центрах Москвы, Астрахани, Казани и Краснодара в 2014–2017 годах. При выписке больных и на 25-й, 90-й, 180-й и 360-й день после выписки регистрировали АД, частоту сердечных сокращений и неблагоприятные события. О межвизитной вариабельности АД судили по коэффициенту VIM (variation independent of mean).
Результаты. Вариабельность систолического АД за период наблюдения составила 7,81 ± 0,226 мм рт. ст., вариабельность диастолического АД — 9,89 ± 0,577 мм рт. ст. За время наблюдения в группе было зарегистрировано 110 случаев смерти от любой причины, 63 коронарные смерти, 130 повторных нефатальных коронарных событий, 33 ишемических инсульта. Снижение вариабельности АД было ассоциировано с приемом дигидропиридиновых антагонистов кальция (10,21 ± 6,45 и 7,99 ± 4,70 мм рт. ст., р = 0,024) и тиазидных диуретиков (10,34 ± 6,59 и 7,63 ± 9,63 мм рт. ст., р = 0,049). Многофакторный анализ показал, что высокая долгосрочная вариабельность АД является более значимым фактором, ассоциированным с уровнем общей смертности, чем исходная тяжесть АГ и даже факт достижения целевого АД. Риск ишемического инсульта у больных АГ был ассоциирован с такими факторами, как наличие фибрилляции предсердий, сердечной недостаточности, анамнез инсульта и высокая межвизитная вариабельность АД.
ст., р = 0,024) и тиазидных диуретиков (10,34 ± 6,59 и 7,63 ± 9,63 мм рт. ст., р = 0,049). Многофакторный анализ показал, что высокая долгосрочная вариабельность АД является более значимым фактором, ассоциированным с уровнем общей смертности, чем исходная тяжесть АГ и даже факт достижения целевого АД. Риск ишемического инсульта у больных АГ был ассоциирован с такими факторами, как наличие фибрилляции предсердий, сердечной недостаточности, анамнез инсульта и высокая межвизитная вариабельность АД.
Заключение. Межвизитная вариабельность АД является важной характеристикой контроля за АД и ассоциирована с риском смерти от любых причин и инсульта у больных, перенесших обострение ишемической болезни сердца.
Инструмент для обжима, Обжимной инструмент для контактов, 0.5 мм.кв, 6 мм.кв, Обжим с выемкой
Обжимной инструмент;
Банковский перевод: счет на оплату формируется после оформления заказа или отправки заявки в произвольной форме на электронную почту info@euro-avtomatika. ru. Специалист свяжется с вами для уточнения деталей.
ru. Специалист свяжется с вами для уточнения деталей.
Самовывоз с нашего склада:
По адресу: Московская область, Люберецкий район, п. Томилино, мкр. Птицефабрика, стр. лит. А, офис 109. Мы есть на Яндекс.Карты.
Доставка до двери
Осуществляется курьерской службой или транспортной компанией (на Ваш выбор).
Мы работаем с ведущими транспортными компаниями и доставляем заказы во все регионы России и Казахстана.
Доставка до терминала
Транспортной компании в Москва – БЕСПЛАТНО.
| Australia | |
| Alfred Hospital | |
| Мельбурн, Австралия | |
| Австрия | |
| Венская больница общего профиля | |
| Вена, Австрия | |
| Бельгия | |
| Cliniques Universitaires Saint Luc | |
| Брюссель, Бельгия | |
| Франция | |
| Européen Georges Pompidou | |
| Париж, Франция | |
| Германия | |
| Herzzentrum Bad Krozingen | |
| Бад-Кроцинген, Германия | |
| Universität zu Köln | |
| Кельн, Германия | |
| Universitaetsklinikum Dusseldorf | |
| Дюссельдорф, Германия | |
| Эрлангенский университет в Нюрнберге | |
| Эрланген, Германия | |
| Universitaetsklinikum Essen | |
| Эссен, Германия | |
| Кардио-сосудистый центр Франкфурт | |
| Франкфурт, Германия | |
| Universitatskliniken des Saarlandes | |
| Хомбург, Германия | |
| Лейпцигский университет – Герццентрум | |
| Лейпциг, Германия | |
| Universitatsklinikum Schleswig-Holstein, Luebeck Campus | |
| Лейбек, Германия | |
| Латвия | |
| Клиническая университетская больница имени Паула Страдиня | |
| Рига, Латвия | |
| Польша | |
| Samodzielna Pracownia Hemodynamiczna | |
| Варшава, Польша | |
| Испания | |
| Больница 12 октября | |
| Мадрид, Испания | |
| Швейцария | |
| Universitaatsspital Zurich | |
| Цюрих, Швейцария | |
| Великобритания | |
| Больница Кент и Кентербери | |
| Кентербери, Великобритания | |
| Университет Глазго | |
| Глазго, Великобритания | |
| Бартс и Лондонская школа стоматологии и медицины | |
| Лондон, Соединенное Королевство | |
| Cardiology, PC | |
| Бирмингем, Алабама, США, 35211 | |
| Университет Алабамы в Бирмингеме | |
| Бирмингем, Алабама, США, 35294 | |
| Клиника Скриппса / Скриппс Грин Госпиталь | |
| Ла-Хойя, Калифорния, США, | |
| Kaiser Permanente – Медицинский центр Лос-Анджелеса | |
| Лос-Анджелес, Калифорния, США, | |
| Cedars Sinai Medical Center | |
| Лос-Анджелес, Калифорния, США, | |
St. Больница Иосифа Больница Иосифа | |
| Оранж, Калифорния, США, | |
| Кардиологический центр Сан-Диего | |
| Сан-Диего, Калифорния, США, | |
| Стэнфордский университет | |
| Стэнфорд, Калифорния, США, 94305 | |
| Больница Университета Колорадо | |
| Аврора, Колорадо, США, 80045 | |
| Больничный центр MedStar Washington | |
Вашингтон, Д. C., Округ Колумбия, США, 20010 C., Округ Колумбия, США, 20010 | |
| Больница Ховардского университета | |
| Вашингтон, округ Колумбия, округ Колумбия, США, 20060 | |
| Шандс / Университет Флориды | |
| Гейнсвилл, Флорида, США, 32610 | |
| Mayo Clinic | |
| Джексонвилл, Флорида, США, 32224 | |
| Университет Майами | |
| Майами, Флорида, США, 33136 | |
| Баптистский институт сердца и сосудов, Baptist Health Systems | |
| Майами, Флорида, США, 33176 | |
| Больница общего профиля Тампа | |
| Тампа, Флорида, США, 33606 | |
| Университетская больница Эмори Мидтаун | |
| Атланта, Джорджия, США, 30308 | |
| Пьемонтский институт сердца | |
| Атланта, Джорджия, США, 30309 | |
| Северо-Западный университет | |
| Чикаго, Иллинойс, США, 60611 | |
| Чикагский университет | |
| Чикаго, Иллинойс, США, 60637 | |
| Midwest Heart Foundation | |
| Окбрук Террас, Иллинойс, США, 60181 | |
| Prairie Heart Institute | |
| Спрингфилд, Иллинойс, США, 62701 | |
| Iowa Heart Center Research | |
| Де-Мойн, Айова, США, 50266 | |
| Университет Кентукки | |
| Лексингтон, Кентукки, США, 40536 | |
| Ochsner Medical Center | |
| Новый Орлеан, Луизиана, США, 70121 | |
| Университет Мэриленда, Балтимор | |
| Балтимор, Мэриленд, США, 21201 | |
| Бригам и женская больница | |
| Бостон, Массачусетс, США, 02115 | |
| VA Boston Healthcare System | |
| Бостон, Массачусетс, США, 02132 | |
| Клиника Лахи | |
| Берлингтон, Массачусетс, США, 01805 | |
| UMass Memorial Medical Center | |
| Вустер, Массачусетс, США, 01655 | |
| Мичиганский университет | |
| Энн-Арбор, Мичиган, США, 48109 | |
| Больница Харперского университета – Детройтский медицинский центр | |
| Детройт, Мичиган, США, 48201 | |
| Госпиталь Святого Джозефа Мерси Окленд | |
| Понтиак, Мичиган, США, 48341 | |
| Больница Провиденс | |
| Саутфилд, Мичиган, США, 48075 | |
Michigan Heart, St. Система здравоохранения Джозефа Мерси Система здравоохранения Джозефа Мерси | |
| Ипсиланти, Мичиган, США, 48197 | |
| Abbott Northwestern / Minneapolis Heart Institute Foundation | |
| Миннеаполис, Миннесота, США, 55407 | |
| Клиника Святой Марии Майо | |
| Рочестер, Миннесота, США, 55905 | |
| Hattiesburg Clinic, P.A | |
| Хаттисберг, Миссисипи, США, 39401 | |
| Больница Святого Луки в Канзас-Сити / Mid America Heart Institute | |
| Канзас-Сити, штат Миссури, США, 64111 | |
| Медицинский факультет Вашингтонского университета | |
| Сент-Луис, штат Миссури, США, 63110 | |
| Центр сердца и легких Деборы | |
| Browns Mills, Нью-Джерси, США, 08015 | |
| Медицинский центр Морристауна | |
| Морристаун, Нью-Джерси, США, 07962 | |
| Медицинский центр Университета Джерси Шор | |
| Neptune City, Нью-Джерси, США, 07753 | |
| SUNY Downstate Medical Center | |
| Бруклин, Нью-Йорк, США, 11203 | |
| Медицинский центр Нью-Йоркского университета | |
| Нью-Йорк, Нью-Йорк, США, 10015 | |
| Weill Cornell Medical College | |
| Нью-Йорк, Нью-Йорк, США, 10021 | |
| Медицинский центр Маунт-Синай – Манхэттен | |
| Нью-Йорк, Нью-Йорк, США, 10029 | |
| Медицинский центр Колумбийского университета | |
| Нью-Йорк, Нью-Йорк, США, 10032 | |
St. Госпиталь Фрэнсиса Госпиталь Фрэнсиса | |
| Рослин, Нью-Йорк, США, 11576 | |
| Университетская больница и медицинский центр Стоуни-Брук | |
| Стоуни-Брук, Нью-Йорк, США, 11794 | |
| Университет сердца и сосудов Северной Каролины | |
| Чапел-Хилл, Северная Каролина, США, 27599 | |
| Carolinas Healthcare (SHVI) – Клинические исследования | |
| Шарлотт, Северная Каролина, США, 28203 | |
| Система здравоохранения Университета Дьюка | |
| Дарем, Северная Каролина, США, 27710 | |
| Wake Forest Baptist Health | |
| Уинстон-Салем, Северная Каролина, США, 27157 | |
| Центр исследований и образования Линднера в больнице Христа | |
| Цинциннати, Огайо, США, 45219 | |
| Университетские больницы Case Medical Center | |
| Кливленд, Огайо, США, 44106 | |
| Система MetroHealth | |
| Кливленд, Огайо, США, 44109 | |
| Cleveland Clinic Foundation | |
| Кливленд, Огайо, США, 44195 | |
| Государственный университет Огайо | |
| Колумбус, Огайо, США, 43210 | |
| Ohio Health Research Institute | |
| Колумбус, Огайо, США, 43214-3907 | |
| Geisinger Medical Center | |
| Данвилл, Пенсильвания, США, 17822 | |
| Ланкастерская больница общего профиля | |
| Ланкастер, Пенсильвания, США, 17604 | |
| Университетские больницы Томаса Джефферсона, Inc | |
| Филадельфия, Пенсильвания, США, 19107 | |
| Система здравоохранения Пенсильванского университета | |
| Филадельфия, Пенсильвания, США, 19130 | |
| UPMC | |
| Питтсбург, Пенсильвания, США, 15213 | |
| Медицинский центр Ланкенау | |
| Виннвуд, Пенсильвания, США, 19096 | |
| Исследовательский центр сосудистых заболеваний при больнице Род-Айленда | |
| Провиденс, Род-Айленд, США, 02903 | |
| Медицинский университет Южной Каролины | |
| Чарлстон, Южная Каролина, США, 29425 | |
Stern Cardiovascular Foundation, Inc. | |
| Мемфис, Теннесси, США, 38138 | |
| Медицинский центр Вандербильта | |
| Нашвилл, Теннесси, США, 37232-7235 | |
| Austin Heart PLLC / Heart Hospital of Austin | |
| Остин, Техас, США, 78756 | |
| VA North Texas Health Care System | |
| Даллас, Техас, США, 75216 | |
| Центр сердечно-сосудистых исследований Солтеро – Больница сердца и сосудов Бейлор Джека и Джейн Гамильтон | |
| Даллас, Техас, США, 75226 | |
| Методистская больница | |
| Хьюстон, Техас, США, 77030 | |
| Больница сердца Бейлор Плано | |
| Плано, Техас, США, 75093 | |
| Fletcher Allen Health Care | |
| Берлингтон, Вермонт, США, 05401 | |
| Университет Вирджинии | |
| Шарлоттсвилль, Вирджиния, США, 22908 | |
| Университет Содружества Вирджинии | |
| Ричмонд, Вирджиния, США, 23298 | |
Aurora St. Медицинский центр Люка Медицинский центр Люка | |
| Милуоки, Висконсин, США, 53215 | |
REDUCE HTN: REINFORCE: Рандомизированное контролируемое испытание биполярной радиочастотной денервации почек для лечения гипертензии
Цели: Целью этого исследования было изучить биполярную радиочастотную денервацию почек у пациентов с артериальной гипертензией, не получавших лекарства на исходном уровне.
Задний план: Эффект снижения артериального давления от почечной денервации трудно выделить в клинических исследованиях.
Методы: REDUCE HTN: REINFORCE (Денервация почек с использованием системы почечной денервации Vessix для лечения гипертонии) было рандомизированным мнимо контролируемым многоцентровым испытанием. Пациентам с офисным систолическим артериальным давлением (САД) от 150 до 180 мм рт.ст. и средним суточным амбулаторным САД от 135 до 170 мм рт.ст. после вымывания лекарства проводилась биполярная радиочастотная денервация почек или имитационная процедура. Планируемым результатом было 8-недельное изменение суточного амбулаторного САД. Набор был прекращен из-за очевидной бесполезности до того, как была набрана достаточная выборка для мощных сравнений эффективности. Оценка безопасности включала смерть от всех причин, почечную недостаточность, тяжелую гипотензию или обморок, гипертонический криз и стеноз почечной артерии.
Пациентам с офисным систолическим артериальным давлением (САД) от 150 до 180 мм рт.ст. и средним суточным амбулаторным САД от 135 до 170 мм рт.ст. после вымывания лекарства проводилась биполярная радиочастотная денервация почек или имитационная процедура. Планируемым результатом было 8-недельное изменение суточного амбулаторного САД. Набор был прекращен из-за очевидной бесполезности до того, как была набрана достаточная выборка для мощных сравнений эффективности. Оценка безопасности включала смерть от всех причин, почечную недостаточность, тяжелую гипотензию или обморок, гипертонический криз и стеноз почечной артерии.
Результаты: Исходное 24-часовое артериальное давление составляло 148,3 ± 10,9 / 85,7 ± 9,1 мм рт. Ст. Для группы денервации (n = 34, средний возраст 58,5 ± 10,1 года, 47% женщин) и 149,1 ± 7,2 / 86,4 ± 9,8 мм рт. (n = 17, средний возраст 58,2 ± 9,8 года, 24% женщин). Через 8 недель среднее 24-часовое снижение САД для почечной денервации и контрольной группы составило -5,3 мм рт.ст. (95% доверительный интервал [ДИ]: -8.От 8 до -1,8 мм рт. Ст.) И -8,5 мм рт. Ст. (95% ДИ: от -13,3 до -3,8 мм рт. Ст.), Соответственно (разница 3,3 мм рт. Затем могут быть добавлены антигипертензивные препараты. К 6 месяцам снижение САД было больше в группе денервации, давая межгрупповые различия -7,2 мм рт. Ст. (95% ДИ: от -15,2 до 0,8 мм рт. Ст .; p = 0,08), -9,7 мм рт. От -17,7 до -1,7 мм рт.ст .; p = 0,02) и -11,4 мм рт. Ст. (95% ДИ: от -19,2 до -3,7 мм рт.Через 12 месяцев у 1 пациента (группа почечной денервации) возникли позывы к гипертонической болезни, требующие немедленного лечения, и у 1 пациента наблюдалось прогрессирование стеноза почечной артерии.
Через 8 недель среднее 24-часовое снижение САД для почечной денервации и контрольной группы составило -5,3 мм рт.ст. (95% доверительный интервал [ДИ]: -8.От 8 до -1,8 мм рт. Ст.) И -8,5 мм рт. Ст. (95% ДИ: от -13,3 до -3,8 мм рт. Ст.), Соответственно (разница 3,3 мм рт. Затем могут быть добавлены антигипертензивные препараты. К 6 месяцам снижение САД было больше в группе денервации, давая межгрупповые различия -7,2 мм рт. Ст. (95% ДИ: от -15,2 до 0,8 мм рт. Ст .; p = 0,08), -9,7 мм рт. От -17,7 до -1,7 мм рт.ст .; p = 0,02) и -11,4 мм рт. Ст. (95% ДИ: от -19,2 до -3,7 мм рт.Через 12 месяцев у 1 пациента (группа почечной денервации) возникли позывы к гипертонической болезни, требующие немедленного лечения, и у 1 пациента наблюдалось прогрессирование стеноза почечной артерии.
Выводы: Будущие исследования радиочастотной денервации почек должны предвидеть отсроченные эффекты лечения. (Денервация почек с использованием системы почечной денервации Vessix для лечения гипертонии [REDUCE HTN: REINFORCE]; NCT023
(Денервация почек с использованием системы почечной денервации Vessix для лечения гипертонии [REDUCE HTN: REINFORCE]; NCT023
).
Ключевые слова: гипотензивные средства; катетерная абляция; гипертония; почечная артерия.
Симпатическая денервация почек у пациентов с терапевтически резистентной гипертензией (The Symplicity HTN-2 Trial): рандомизированное контролируемое исследование
Задний план: Активация почечных симпатических нервов является ключом к патогенезу гипертонической болезни.Мы стремились оценить эффективность и безопасность катетерной денервации почек для снижения артериального давления у пациентов с терапевтически резистентной артериальной гипертензией.
Методы: В этом многоцентровом проспективном рандомизированном исследовании пациенты с исходным систолическим артериальным давлением 160 мм рт. Отношение к одному для проведения денервации почек при предыдущем лечении или для продолжения только предыдущего лечения (контрольная группа) в 24 участвующих центрах.Рандомизация проводилась с запечатанными конвертами. Анализаторы данных не маскировались для назначения лечения. Первичной конечной точкой эффективности было изменение измерения систолического артериального давления в офисе через 6 месяцев. Первичный анализ включал всех пациентов, оставшихся под наблюдением через 6 месяцев. Это испытание зарегистрировано на сайте ClinicalTrials.gov, номер NCT00888433.
Отношение к одному для проведения денервации почек при предыдущем лечении или для продолжения только предыдущего лечения (контрольная группа) в 24 участвующих центрах.Рандомизация проводилась с запечатанными конвертами. Анализаторы данных не маскировались для назначения лечения. Первичной конечной точкой эффективности было изменение измерения систолического артериального давления в офисе через 6 месяцев. Первичный анализ включал всех пациентов, оставшихся под наблюдением через 6 месяцев. Это испытание зарегистрировано на сайте ClinicalTrials.gov, номер NCT00888433.
Выводы: 106 (56%) из 190 пациентов, прошедших скрининг на соответствие критериям, были случайным образом распределены в группы почечной денервации (n = 52) или контрольные (n = 54) в период с 9 июня 2009 г. по 15 января 2010 г.49 (94%) из 52 пациентов, перенесших денервацию почек, и 51 (94%) из 54 пациентов контрольной группы были оценены на предмет первичной конечной точки через 6 месяцев. Офисные измерения артериального давления в группе почечной денервации снизились на 32/12 мм рт. Ст. (Стандартное отклонение 23/11, исходный уровень 178/96 мм рт. (изменение на 1/0 мм рт. ст. [21/10], исходный уровень 178/97 мм рт. ст., p = 0,77 систолическое и p = 0,83 диастолическое). Межгрупповые различия артериального давления через 6 месяцев составили 33/11 мм рт. Ст. (P <0,00001).Через 6 месяцев у 41 (84%) из 49 пациентов, перенесших почечную денервацию, наблюдалось снижение систолического артериального давления на 10 мм рт.ст. или более по сравнению с 18 (35%) из 51 контрольной группы (p <0,00001). Мы не отметили серьезных осложнений, связанных с процедурой или устройством, а частота нежелательных явлений не различалась между группами; у одного пациента, у которого была денервация почек, возможно прогрессирование лежащего в основе атеросклеротического поражения, но лечения не потребовалось.
Офисные измерения артериального давления в группе почечной денервации снизились на 32/12 мм рт. Ст. (Стандартное отклонение 23/11, исходный уровень 178/96 мм рт. (изменение на 1/0 мм рт. ст. [21/10], исходный уровень 178/97 мм рт. ст., p = 0,77 систолическое и p = 0,83 диастолическое). Межгрупповые различия артериального давления через 6 месяцев составили 33/11 мм рт. Ст. (P <0,00001).Через 6 месяцев у 41 (84%) из 49 пациентов, перенесших почечную денервацию, наблюдалось снижение систолического артериального давления на 10 мм рт.ст. или более по сравнению с 18 (35%) из 51 контрольной группы (p <0,00001). Мы не отметили серьезных осложнений, связанных с процедурой или устройством, а частота нежелательных явлений не различалась между группами; у одного пациента, у которого была денервация почек, возможно прогрессирование лежащего в основе атеросклеротического поражения, но лечения не потребовалось.
Интерпретация: Катетерная денервация почек может безопасно использоваться для значительного снижения артериального давления у резистентных к лечению пациентов с гипертонией.
Финансирование: Ардиан.
Роль 5-HT рецепторов при депрессии | Молекулярный мозг
Смит К. Психическое здоровье: мир депрессии. Природа. 2014; 515 (7526): 181.
PubMed Статья CAS Google Scholar
Мюррей С.Дж., Лопес А.Д.Политика здравоохранения, основанная на фактических данных – уроки исследования глобального бремени болезней. Наука. 1996. 274 (5288): 740–3.
CAS PubMed Статья Google Scholar
Gorman JM. Коморбидная депрессия и расстройства тревожного спектра. Подавить тревогу. 1996. 4 (4): 160–8.
PubMed Статья Google Scholar
Сэмюэлс Б.А., Леонардо Э.Д., Гадиент Р. , Уильямс А., Чжоу Дж., Дэвид Д.Д. и др.Моделирование устойчивой к лечению депрессии. Нейрофармакология. 2011. 61 (3): 408–13.
, Уильямс А., Чжоу Дж., Дэвид Д.Д. и др.Моделирование устойчивой к лечению депрессии. Нейрофармакология. 2011. 61 (3): 408–13.
CAS PubMed PubMed Central Статья Google Scholar
Loomer HP, Saunders JC, Kline NS. Клиническая и фармакодинамическая оценка ипрониазида как психотропного средства. Psychiatr Res Rep Am Psychiatr Assoc. 1957; 8: 129–41.
CAS PubMed Google Scholar
Tatsumi M, Groshan K, Blakely RD, Richelson E. Фармакологический профиль антидепрессантов и родственных соединений в переносчиках моноаминов человека. Eur J Pharmacol. 1997. 340 (2–3): 249–58.
CAS PubMed Статья Google Scholar
Гловински Дж., Аксельрод Дж. Ингибирование поглощения тритированного норадреналина в интактном мозге крысы имипрамином и структурно родственными соединениями. Природа. 1964; 204: 1318–9.
1964; 204: 1318–9.
CAS PubMed Статья Google Scholar
Росс С.Б., Реньи А.Л. Подавление захвата меченного тритием 5-гидрокситриптамина тканью мозга. Eur J Pharmacol. 1969; 7 (3): 270–7.
CAS PubMed Статья Google Scholar
Feighner JP, Boyer WF. Селективные ингибиторы обратного захвата серотонина, 2-е издание: достижения в фундаментальных исследованиях и клинической практике. Чичестер: Уайли; 1996.
Google Scholar
Stahl SM. Механизм действия селективных ингибиторов обратного захвата серотонина. Рецепторы и пути серотонина опосредуют терапевтические эффекты и побочные эффекты. J влияет на Disord. 1998. 51 (3): 215–35.
CAS PubMed Статья Google Scholar
Коэн Э. CDC: Антидепрессанты, назначаемые чаще всего в США: CNN; 2007 [http://www. cnn.com/2007/HEALTH/07/09/antidepressants/]. По состоянию на 1 мая 2017 г.
cnn.com/2007/HEALTH/07/09/antidepressants/]. По состоянию на 1 мая 2017 г.
Massart R, Mongeau R, Lanfumey L.Помимо моноаминергической гипотезы: нейропластичность и эпигенетические изменения в модели депрессии трансгенных мышей. Philos Trans R Soc Lond B Biol Sci. 2012. 367 (1601): 2485–94.
CAS PubMed PubMed Central Статья Google Scholar
Слэттери Д.А., Хадсон А.Л., Натт DJ. Приглашенный обзор: эволюция антидепрессивных механизмов. Fundam Clin Pharmacol. 2004. 18 (1): 1–21.
CAS PubMed Статья Google Scholar
Akiskal HS, McKinney WT Jr. Депрессивные расстройства: к единой гипотезе. Наука. 1973; 182 (4107): 20–9.
CAS PubMed Статья Google Scholar
Баумейстер А.А., Хокинс М.Ф., Узелак С.М. Миф о депрессии, вызванной резерпином: роль в историческом развитии гипотезы моноаминов. J Hist Neurosci. 2003. 12 (2): 207–20.
J Hist Neurosci. 2003. 12 (2): 207–20.
PubMed Статья Google Scholar
Leyton M, Young SN, Benkelfat C. Рецидив депрессии после быстрого истощения триптофана. Ланцет. 1997. 349 (9068): 1840–1.
CAS PubMed Статья Google Scholar
Delgado PL. Исследования истощения моноаминов: последствия для синдрома отмены антидепрессантов. J Clin Psychiatry. 2006; 67 (Дополнение 4): 22–6.
CAS PubMed Google Scholar
Якобсен Дж. П., Медведев И. О., Карон МГ. Теория депрессии с дефицитом 5-HT: перспективы с натуралистической модели дефицита 5-HT, триптофангидроксилазы 2Arg439His, нокаутирующей мышь. Philos Trans R Soc Lond B Biol Sci. 2012. 367 (1601): 2444–59.
CAS PubMed PubMed Central Статья Google Scholar

Траскман Л., Асберг М., Бертилссон Л., Шостранд Л. Метаболиты моноаминов в спинномозговой жидкости и суицидальном поведении.Arch Gen Psychiatry. 1981. 38 (6): 631–6.
CAS PubMed Статья Google Scholar
Placidi GP, Oquendo MA, Malone KM, Huang YY, Ellis SP, Mann JJ. Агрессивность, попытки суицида и депрессия: взаимосвязь с уровнями метаболитов моноаминов спинномозговой жидкости. Биол Психиатрия. 2001. 50 (10): 783–91.
CAS PubMed Статья Google Scholar
Асберг М. Нейротрансмиттеры и суицидальное поведение. Данные исследований спинномозговой жидкости. Ann N Y Acad Sci. 1997; 836: 158–81.
CAS PubMed Статья Google Scholar
Раш А.Дж., Триведи М.Х., Вишневски С.Р., Ниренберг А.А., Стюарт Дж.В., Уорден Д. и др. Острые и отдаленные результаты у амбулаторных пациентов с депрессией, требующие одного или нескольких этапов лечения: отчет STAR * D. Am J Psychiatry. 2006. 163 (11): 1905–17.
Am J Psychiatry. 2006. 163 (11): 1905–17.
PubMed Статья Google Scholar
Надзиратель Д., Раш А.Дж., Триведи М.Х., Фава М., Вишневски С.Р. Результаты проекта STAR * D: всесторонний обзор результатов. Curr Psychiatry Rep. 2007; 9 (6): 449–59.
PubMed Статья Google Scholar
Сарате К.А. младший, Сингх Дж. Б., Карлсон П. Дж., Брутше Н. Э., Амели Р., Лакенбо Д. А. и др. Рандомизированное исследование антагониста N-метил-D-аспартата при резистентной к лечению большой депрессии.Arch Gen Psychiatry. 2006. 63 (8): 856–64.
CAS PubMed Статья Google Scholar
Берман Р.М., Каппиелло А., Ананд А., Орен Д.А., Хенингер Г.Р., Чарни Д.С. и др. Антидепрессивные эффекты кетамина у пациентов с депрессией. Биол Психиатрия. 2000. 47 (4): 351–4.
CAS PubMed Статья Google Scholar
Фурей М.Л., Древец WC. Антидепрессивная эффективность антимускаринового препарата скополамина: рандомизированное плацебо-контролируемое клиническое исследование.Arch Gen Psychiatry. 2006. 63 (10): 1121–9.
CAS PubMed PubMed Central Статья Google Scholar
Карлезон В.А. младший, Бегин С., Нолл А.Т., Коэн Б.М. Каппа-опиоидные лиганды в исследовании и лечении расстройств настроения. Pharmacol Ther. 2009. 123 (3): 334–43.
CAS PubMed PubMed Central Статья Google Scholar
Lutz PE, Kieffer BL.Опиоидные рецепторы: различные роли в расстройствах настроения. Trends Neurosci. 2013. 36 (3): 195–206.
CAS PubMed Статья Google Scholar
Samuels BA, Nautiyal KM, Kruegel AC, Levinstein MR, Magalong VM, Gassaway MM, et al. Поведенческие эффекты антидепрессанта тианептина требуют наличия мю-опиоидного рецептора. Нейропсихофармакология. 2017. Epub опережает печать.
Нейропсихофармакология. 2017. Epub опережает печать.
Каспи А., Сагден К., Моффит Т. Е., Тейлор А., Крейг И. В., Харрингтон Н. и др.Влияние жизненного стресса на депрессию: сдерживание полиморфизмом гена 5-HTT. Наука. 2003. 301 (5631): 386–9.
CAS PubMed Статья Google Scholar
Lesch KP, Bengel D, Heils A, Sabol SZ, Greenberg BD, Petri S, et al. Ассоциация признаков, связанных с тревогой, с полиморфизмом в регуляторной области гена переносчика серотонина. Наука. 1996. 274 (5292): 1527–31.
CAS PubMed Статья Google Scholar
Bosker FJ, Hartman CA, Nolte IM, Prins BP, Terpstra P, Posthuma D, et al. Плохая репликация генов-кандидатов на большое депрессивное расстройство с использованием данных ассоциации по всему геному. Мол Психиатрия. 2011; 16 (5): 516–32.
CAS PubMed Статья Google Scholar

Холмс А., Мерфи Д.Л., Кроули Дж. Аномальные поведенческие фенотипы мышей с нокаутом переносчика серотонина: параллели с тревожностью и депрессией у человека. Биол Психиатрия.2003. 54 (10): 953–9.
CAS PubMed Статья Google Scholar
Лира А., Чжоу М., Кастанон Н., Ансорге М.С., Гордон Дж. А., Фрэнсис Дж. Х. и др. Изменение поведения, связанного с депрессией, и функциональные изменения в ядре спинного шва у мышей с дефицитом переносчика серотонина. Биол Психиатрия. 2003. 54 (10): 960–71.
CAS PubMed Статья Google Scholar
Beck SG, Choi KC, List TJ. Сравнение 5-гидрокситриптамин1A-опосредованной гиперполяризации в пирамидных клетках гиппокампа CA1 и CA3. J Pharmacol Exp Ther. 1992. 263 (1): 350–9.
CAS PubMed Google Scholar
Хамон М., Ланфумей Л., эль-Местикави С. , Бони С., Микель М.К., Боланос Ф. и др. Основные особенности центральных рецепторов 5-HT1. Нейропсихофармакология. 1990. 3 (5–6): 349–60.
, Бони С., Микель М.К., Боланос Ф. и др. Основные особенности центральных рецепторов 5-HT1. Нейропсихофармакология. 1990. 3 (5–6): 349–60.
CAS PubMed Google Scholar
Riad M, Garcia S, Watkins KC, Jodoin N, Doucet E, Langlois X и др. Соматодендритная локализация 5-HT1A и претерминальная аксональная локализация рецепторов 5-HT1B серотонина в головном мозге взрослых крыс. J Comp Neurol. 2000; 417 (2): 181–94.
CAS PubMed Статья Google Scholar
Ричардсон-Джонс Дж. У., Крейдж С. П., Гьярд Б. П., Стивен А., Мецгер К. Л., Кунг Х. Ф. и др. Уровни ауторецепторов 5-HT1A определяют уязвимость к стрессу и реакцию на антидепрессанты.Нейрон. 2010. 65 (1): 40–52.
CAS PubMed PubMed Central Статья Google Scholar
Хэннон Дж., Хойер Д. Молекулярная биология 5-HT рецепторов. Behav Brain Res. 2008. 195 (1): 198–213.
Behav Brain Res. 2008. 195 (1): 198–213.
CAS PubMed Статья Google Scholar
Блиер П., Пинейро Дж., Эль-Мансари М., Бержерон Р., де Монтиньи С. Роль соматодендритных ауторецепторов 5-HT в модулировании нейротрансмиссии 5-HT.Ann N Y Acad Sci. 1998; 861: 204–16.
CAS PubMed Статья Google Scholar
Ричардсон-Джонс JW, Craige CP, Nguyen TH, Kung HF, Gardier AM, Dranovsky A, et al. Ауторецепторы серотонина-1A необходимы и достаточны для нормального формирования цепей, лежащих в основе врожденного беспокойства. J Neurosci. 2011. 31 (16): 6008–18.
CAS PubMed PubMed Central Статья Google Scholar
Gross C, Zhuang X, Stark K, Ramboz S, Oosting R, Kirby L и др. Рецептор серотонина1A действует во время развития, чтобы установить у взрослого человека нормальное тревожное поведение. Природа. 2002. 416 (6879): 396–400.
Природа. 2002. 416 (6879): 396–400.
CAS PubMed Статья Google Scholar
Рэмбоз С., Остинг Р., Амара Д.А., Кунг Х.Ф., Блиер П., Мендельсон М. и др. Нокаут серотонинового рецептора 1A: животная модель тревожного расстройства. Proc Natl Acad Sci U S A.1998. 95 (24): 14476–81.
CAS PubMed PubMed Central Статья Google Scholar
Wu S, Comings DE. Распространенный полиморфизм C-1018G в гене рецептора 5-HT1A человека. Psychiatr Genet. 1999. 9 (2): 105–6.
CAS PubMed Статья Google Scholar
Lemonde S, Turecki G, Bakish D, Du L, Hrdina PD, Bown CD, et al. Нарушение репрессии полиморфизма гена рецептора 5-гидрокситриптамина 1A, связанное с большой депрессией и самоубийством.J Neurosci. 2003. 23 (25): 8788–99.
CAS PubMed Google Scholar

Альберт П.Р., Lemonde S. 5-HT1A рецепторы, репрессия генов и депрессия: вина по ассоциации. Невролог. 2004. 10 (6): 575–93.
CAS PubMed Статья Google Scholar
Czesak M, Le Francois B., Millar AM, Deria M, Daigle M, Visvader JE, et al. Повышенная экспрессия ауторецептора серотонина-1A (5-HT1A) и снижение уровней серотонина шва у мышей с деформированным эпидермальным ауторегуляторным фактором-1 (Deaf-1) с нокаутом гена.J Biol Chem. 2012. 287 (9): 6615–27.
CAS PubMed PubMed Central Статья Google Scholar
Парси Р.В., Гастингс Р.С., Окендо М.А., Хуанг Ю.Й., Симпсон Н., Арцемент Дж. И др. Снижение связывающего потенциала переносчика серотонина в мозге человека во время эпизодов большой депрессии. Am J Psychiatry. 2006. 163 (1): 52–8.
PubMed Статья Google Scholar

Parsey RV, Oquendo MA, Ogden RT, Olvet DM, Simpson N, Huang YY и др. Изменение связывания серотонина 1A при большой депрессии: исследование позитронно-эмиссионной томографии [карбонил-C-11] WAY100635. Биол Психиатрия. 2006. 59 (2): 106–13.
CAS PubMed Статья Google Scholar
Хамон М., Блиер П. Моноаминовая нейросхема при депрессии и стратегии новых методов лечения. Prog Neuro-Psychopharmacol Biol Psychiatry. 2013; 45: 54–63.
CAS Статья Google Scholar
Lanfumey L, Hamon M. 5-HT1 рецепторы. Препарат Curr действует на нервные расстройства в ЦНС. 2004; 3 (1): 1–10.
CAS PubMed Статья Google Scholar
Сэмюэлс Б.А., Хен Р. Кормление мышей с подавлением новизны. В: Гулд Т.Д., редактор. Фенотипы, связанные с настроением и тревогой, у мышей: характеристика с использованием поведенческих тестов, том II. Нейрометоды. Тотова: Humana Press; 2011. с. 107–21.
Нейрометоды. Тотова: Humana Press; 2011. с. 107–21.
Google Scholar
Гарсия-Гарсия А.Л., Ньюман-Танкреди А., Леонардо Э.Д. 5-HT (1A) [скорректированные] рецепторы в настроении и тревоге: недавнее понимание функции ауторецепторов и гетерорецепторов. Психофармакология. 2014. 231 (4): 623–36.
CAS PubMed Статья Google Scholar
Сэмюэлс Б.А., Анакер С., Ху А., Левинштейн М.Р., Пикенхаген А., Цетсенис Т. и др.Рецепторы 5-HT1A на зрелых клетках гранул зубчатой извилины имеют решающее значение для антидепрессивного ответа. Nat Neurosci. 2015; 18 (11): 1606–16.
CAS PubMed PubMed Central Статья Google Scholar
Танака К.Ф., Самуэльс Б.А., Хен Р. Экспрессия рецептора серотонина вдоль дорсально-вентральной оси гиппокампа мыши. Philos Trans R Soc Lond B Biol Sci. 2012. 367 (1601): 2395–401.
367 (1601): 2395–401.
CAS PubMed PubMed Central Статья Google Scholar
Santarelli L, Saxe M, Gross C, Surget A, Battaglia F, Dulawa S и др. Необходимость нейрогенеза гиппокампа для поведенческих эффектов антидепрессантов. Наука. 2003. 301 (5634): 805–9.
CAS PubMed Статья Google Scholar
Мальберг Дж. Э., Эйш А. Дж., Нестлер Э. Дж., Думан Р. С.. Хроническое лечение антидепрессантами увеличивает нейрогенез в гиппокампе взрослых крыс. J Neurosci. 2000. 20 (24): 9104–10.
CAS PubMed Google Scholar
McAskill R, Mir S, Taylor D. Увеличение пиндолола антидепрессивной терапии. Br J Psychiatry. 1998. 173: 203–8.
CAS PubMed Статья Google Scholar
Newman-Tancredi A, Kleven MS. Сравнительная фармакология антипсихотических средств, обладающих комбинированными свойствами дофамина D2 и серотонинового 5-HT1A рецептора. Психофармакология. 2011; 216 (4): 451–73.
Психофармакология. 2011; 216 (4): 451–73.
CAS PubMed Статья Google Scholar
Neumaier JF, Edwards E, Plotsky PM. Регулирование 5-HT (1B) мРНК в двух моделях животных с измененной реактивностью на стресс. Биол Психиатрия. 2002. 51 (11): 902–8.
CAS PubMed Статья Google Scholar
Ruf BM, Bhagwagar Z. Рецептор 5-HT1B: новая мишень для патофизиологии депрессии. Curr Drug Targets. 2009. 10 (11): 1118–38.
CAS PubMed Статья Google Scholar
Chenu F, David DJ, Leroux-Nicollet I, Le Maitre E, Gardier AM, Bourin M. Активация гетерорецептора серотонина1B вызывает у мышей антидепрессантный эффект с изменением серотонинергической системы. J Psychiatry Neurosci. 2008. 33 (6): 541–50.
PubMed PubMed Central Google Scholar

Дин С., Ли Л., Чжоу ФМ. Устойчивое пресинаптическое подавление серотониновых 5-HT (1B) рецепторов стриатонигрального выхода и его сенсибилизация при хроническом лечении флуоксетином.J Neurophysiol. 2015; 113 (9): 3397–409.
CAS PubMed PubMed Central Статья Google Scholar
Gardier AM, Trillat AC, Malagie I, David D, Hascoet M, Colombel MC, et al. 5-HT1B рецепторы серотонина и антидепрессивные эффекты селективных ингибиторов обратного захвата серотонина. C R Acad Sci III. 2001. 324 (5): 433–41.
CAS PubMed Статья Google Scholar
Лю Ю., Келли М.А., Секстон Т.Дж., Ноймайер Дж.Ф. Ауторецепторы 5-HT1B по-разному модулируют выражение условного страха в зависимости от схемы. Неврология. 2015; 298: 436–47.
CAS PubMed PubMed Central Статья Google Scholar

Наутиял К.М., Тричлер Л., Ахмари С.Е., Дэвид Д.Дж., Гардиер А.М., Хен Р. Недостаток ауторецепторов серотонина 1B приводит к снижению тревожности и поведения, связанного с депрессией.Нейропсихофармакология. 2016; 41 (12): 2941–50.
CAS PubMed Статья Google Scholar
McDevitt RA, Hiroi R, Mackenzie SM, Robin NC, Cohn A, Kim JJ, et al. Ауторецепторы серотонина 1B, берущие начало в ядре каудального дорсального шва, снижают проявление страха и депрессивного поведения. Биол Психиатрия. 2011; 69 (8): 780–7.
CAS PubMed PubMed Central Статья Google Scholar
Neumaier JF, Petty F, Kramer GL, Szot P, Hamblin MW. Приобретенная беспомощность увеличивает уровни мРНК рецептора 5-гидрокситриптамина1B в ядре спинного шва крысы. Биол Психиатрия. 1997. 41 (6): 668–74.
CAS PubMed Статья Google Scholar

Доусон Л.А., Хьюз З.А., Старр К.Р., Стори Д.Д., Беттелини Л., Бакки Ф. и др. Характеристика селективного антагониста рецептора 5-HT1B SB-616234-a (1- [6- (цис-3,5-диметилпиперазин-1-ил) -2,3-дигидро-5-метоксииндол-1-ил] -1 – [2′-метил-4 ‘- (5-метил-1,2,4-оксадиазол-3-ил) бифенил-4-ил] метанон гидрохлорид): нейрохимические и поведенческие свидетельства анксиолитической / антидепрессивной активности in vivo.Нейрофармакология. 2006. 50 (8): 975–83.
CAS PubMed Статья Google Scholar
Tatarczynska E, Klodzinska A, Chojnacka-Wojcik E. Эффекты комбинированного введения антагонистов рецепторов 5-HT1A и / или 5-HT1B и пароксетина или флуоксетина в тесте принудительного плавания на крысах. Pol J Pharmacol. 2002. 54 (6): 615–23.
CAS PubMed Google Scholar
Tatarczynska E, Klodzinska A, Stachowicz K, Chojnacka-Wojcik E. Эффекты селективного агониста и антагонистов рецептора 5-HT1B на животных моделях тревоги и депрессии. Behav Pharmacol. 2004. 15 (8): 523–34.
CAS PubMed Статья Google Scholar
Tatarczynska E, Klodzinska A, Stachowicz K, Chojnacka-Wojcik E. Эффект комбинированного введения антагонистов 5-HT1A или 5-HT1B / 1D рецепторов и антидепрессантов в тесте принудительного плавания.Eur J Pharmacol. 2004. 487 (1–3): 133–42.
CAS PubMed Статья Google Scholar
Banasr M, Hery M, Printemps R, Daszuta A. Индуцированное серотонином увеличение пролиферации и нейрогенеза взрослых клеток опосредуется различными и общими подтипами рецепторов 5-HT в зубчатой извилине и субвентрикулярной зоне. Нейропсихофармакология. 2004. 29 (3): 450–60.
CAS PubMed Статья Google Scholar
Carr GV, Lucki I. Роль подтипов рецепторов серотонина в лечении депрессии: обзор исследований на животных. Психофармакология. 2011. 213 (2–3): 265–87.
CAS PubMed Статья Google Scholar
Niswender CM, Herrick-Davis K, Dilley GE, Meltzer HY, Overholser JC, Stockmeier CA, et al. Редактирование РНК рецептора серотонина 5-HT2C человека. Изменения в суициде и последствиях серотонинергической фармакотерапии.Нейропсихофармакология. 2001. 24 (5): 478–91.
CAS PubMed Статья Google Scholar
Kennedy SH, Emsley R. Плацебо-контролируемое испытание агомелатина при лечении большого депрессивного расстройства. Eur Neuropsychopharmacol. 2006. 16 (2): 93–100.
CAS PubMed Статья Google Scholar
Оли Дж. П., Каспер С. Эффективность агомелатина, агониста рецептора MT1 / MT2 с антагонистическими свойствами 5-HT2C, при большом депрессивном расстройстве.Int J Neuropsychopharmacol. 2007. 10 (5): 661–73.
CAS PubMed Google Scholar
Loo H, Hale A, D’Haenen H. Определение дозы агомелатина, мелатонинергического агониста и селективного антагониста 5-HT (2C), при лечении большого депрессивного расстройства: плацебо-контролируемая доза диапазон исследования. Int Clin Psychopharmacol. 2002. 17 (5): 239–47.
CAS PubMed Статья Google Scholar
Rainer Q, Xia L, Guilloux JP, Gabriel C, Mocaer E, Hen R, et al. Благоприятные поведенческие и нейрогенные эффекты агомелатина на модели депрессии / тревоги. Int J Neuropsychopharmacol. 2012. 15 (3): 321–35.
CAS PubMed Статья Google Scholar
Papp M, Gruca P, Boyer PA, Mocaer E. Эффект агомелатина в модели депрессии у крыс с хроническим умеренным стрессом. Нейропсихофармакология. 2003. 28 (4): 694–703.
CAS PubMed Статья Google Scholar
Gorman JM, Liebowitz MR, Fyer AJ, Goetz D, Campeas RB, Fyer MR, et al. Открытое испытание флуоксетина в лечении панических атак. J Clin Psychopharmacol. 1987. 7 (5): 329–32.
CAS PubMed Статья Google Scholar
Westenberg HG, den Boer JA. Препараты, влияющие на серотонин, в лечении панического расстройства.Психопатология. 1989; 22 (Приложение 1): 68–77.
PubMed Статья Google Scholar
Burghardt NS, Bush DE, McEwen BS, LeDoux JE. Острые селективные ингибиторы обратного захвата серотонина усиливают условное выражение страха: блокада антагонистом рецептора 5-HT (2C). Биол Психиатрия. 2007. 62 (10): 1111–8.
CAS PubMed PubMed Central Статья Google Scholar
Dekeyne A, Denorme B, Monneyron S, Millan MJ. Циталопрам снижает социальное взаимодействие у крыс за счет активации рецепторов серотонина (5-HT) (2C). Нейрофармакология. 2000. 39 (6): 1114–7.
CAS PubMed Статья Google Scholar
Belzung C, Le Guisquet AM, Barreau S, Calatayud F. Исследование механизмов, ответственных за острые индуцированные флуоксетином анксиогенные эффекты у мышей. Behav Pharmacol. 2001. 12 (3): 151–62.
CAS PubMed Статья Google Scholar
Marcinkiewcz CA, Mazzone CM, D’Agostino G, Halladay LR, Hardaway JA, DiBerto JF, et al. Серотонин задействует цепь, вызывающую тревогу и страх, в расширенной миндалине. Природа. 2016; 537 (7618): 97–101.
CAS PubMed PubMed Central Статья Google Scholar
Охцуки Т., Исигуро Х., Детера-Уодли С.Д., Тойота Т, Симидзу Х., Ямада К. и др.Связь между полиморфизмом гена рецептора серотонина 4 и биполярным расстройством в японских образцах случай-контроль и биполярных родословных генетической инициативы NIMH. Мол Психиатрия. 2002. 7 (9): 954–61.
CAS PubMed Статья Google Scholar
Rosel P, Arranz B, Urretavizcaya M, Oros M, San L, Navarro MA. Изменены постсинаптические рецепторы 5-HT2A и 5-HT4 и их внутриклеточные сигнальные системы IP3 и cAMP в головном мозге жертв жестокого самоубийства в депрессии.Нейропсихобиология. 2004. 49 (4): 189–95.
CAS PubMed Статья Google Scholar
Madsen K, Torstensen E, Holst KK, Haahr ME, Knorr U, Frokjaer VG, et al. Семейный риск большой депрессии связан с более низким связыванием рецептора 5-HT (4) полосатого тела. Int J Neuropsychopharmacol. 2014; 18 (1): 1-7.
Compan V, Zhou M, Grailhe R, Gazzara RA, Martin R, Gingrich J, et al. Ослабленная реакция на стресс и новизна и гиперчувствительность к судорогам у мышей с нокаутом рецептора 5-HT4.J Neurosci. 2004. 24 (2): 412–9.
CAS PubMed Статья Google Scholar
Conductier G, Dusticier N, Lucas G, Cote F, Debonnel G, Daszuta A, et al. Адаптивные изменения серотониновых нейронов ядер шва у мышей с нокаутом по 5-HT (4) рецепторам. Eur J Neurosci. 2006. 24 (4): 1053–62.
PubMed Статья Google Scholar
Лукас Дж., Рымар В.В., Ду Дж., Мни-Филали О., Бисгаард С., Манта С. и др.Агонисты рецепторов серотонина (4) (5-HT (4)) являются предполагаемыми антидепрессантами с быстрым началом действия. Нейрон. 2007. 55 (5): 712–25.
CAS PubMed Статья Google Scholar
Mendez-David I, David DJ, Darcet F, Wu MV, Kerdine-Romer S, Gardier AM, et al. Быстрые анксиолитические эффекты агониста рецептора 5-HT (4) опосредуются независимым от нейрогенеза механизмом. Нейропсихофармакология. 2014; 39 (6): 1366–78.
CAS PubMed PubMed Central Статья Google Scholar
Cryan JF, Lucki I. Рецепторы 5-HT4 не опосредуют антидепрессантные поведенческие эффекты флуоксетина в модифицированном тесте принудительного плавания. Eur J Pharmacol. 2000. 409 (3): 295–9.
CAS PubMed Статья Google Scholar
Carr GV, Schechter LE, Lucki I. Антидепрессантные и анксиолитические эффекты селективных агонистов рецептора 5-HT6 у крыс. Психофармакология. 2011. 213 (2–3): 499–507.
CAS PubMed Статья Google Scholar
Wesolowska A. Возможная роль рецептора 5-HT6 при депрессии и тревоге: обзор доклинических данных. Pharmacol Rep. 2010; 62 (4): 564–77.
CAS PubMed Статья Google Scholar
Wesolowska A, Nikiforuk A. Эффекты проникающего в мозг и селективного антагониста рецептора 5-HT6 SB-399885 на животных моделях тревоги и депрессии. Нейрофармакология. 2007. 52 (5): 1274–83.
CAS PubMed Статья Google Scholar
Wesolowska A, Nikiforuk A. Селективный антагонист рецептора 5-HT (6) SB-399885 усиливает действие антидепрессантов против неподвижности у крыс. Eur J Pharmacol. 2008. 582 (1–3): 88–93.
CAS PubMed Статья Google Scholar
Svenningsson P, Tzavara ET, Qi H, Carruthers R, Witkin JM, Nomikos GG, et al. Биохимические и поведенческие доказательства антидепрессивных эффектов стимуляции рецептора 5-HT6. J Neurosci.2007. 27 (15): 4201–9.
CAS PubMed Статья Google Scholar
Laplante P, Diorio J, Meaney MJ. Серотонин регулирует экспрессию рецептора глюкокортикоидов в гиппокампе через рецептор 5-HT7. Brain Res Dev Brain Res. 2002. 139 (2): 199–203.
CAS PubMed Статья Google Scholar
Яу Дж. Л., Благородный Дж., Секл Дж. Р. Острый сдерживающий стресс увеличивает экспрессию мРНК рецептора 5-HT7 в гиппокампе крысы.Neurosci Lett. 2001. 309 (3): 141–4.
CAS PubMed Статья Google Scholar
Mullins UL, Gianutsos G, Eison AS. Влияние антидепрессантов на регуляцию рецепторов 5-HT7 в гипоталамусе крыс. Нейропсихофармакология. 1999. 21 (3): 352–67.
CAS PubMed Статья Google Scholar
Hedlund PB, Huitron-Resendiz S, Henriksen SJ, Sutcliffe JG.Ингибирование и инактивация рецептора 5-HT7 вызывает антидепрессантоподобное поведение и характер сна. Биол Психиатрия. 2005. 58 (10): 831–7.
CAS PubMed Статья Google Scholar
Бонавентура П., Келли Л., Алуизио Л., Шелтон Дж., Лорд Б., Галичи Р. и др. Селективная блокада рецепторов 5-гидрокситриптамина (5-HT) 7 усиливает передачу 5-HT, антидепрессантоподобное поведение и подавление сна с быстрым движением глаз, вызванное циталопрамом у грызунов.J Pharmacol Exp Ther. 2007. 321 (2): 690–8.
CAS PubMed Статья Google Scholar
Guscott M, Bristow LJ, Hadingham K, Rosahl TW, Beer MS, Stanton JA, et al. Исследования генетического нокаута и фармакологической блокады рецептора 5-HT7 предполагают терапевтический потенциал при депрессии. Нейрофармакология. 2005. 48 (4): 492–502.
CAS PubMed Статья Google Scholar
Wesolowska A, Nikiforuk A, Stachowicz K. Возможные анксиолитические и антидепрессивные эффекты селективного антагониста 5-HT7 рецепторов SB 269970 после внутригиппокампального введения крысам. Eur J Pharmacol. 2006. 553 (1–3): 185–90.
CAS PubMed Статья Google Scholar
Wesolowska A, Nikiforuk A, Stachowicz K, Tatarczynska E. Эффект селективного антагониста рецептора 5-HT7 SB 269970 на животных моделях тревоги и депрессии.Нейрофармакология. 2006. 51 (3): 578–86.
CAS PubMed Статья Google Scholar
Abbas AI, Hedlund PB, Huang XP, Tran TB, Meltzer HY, Roth BL. Амисульприд является мощным антагонистом 5-HT7: имеет значение для антидепрессивного действия in vivo. Психофармакология. 2009. 205 (1): 119–28.
CAS PubMed PubMed Central Статья Google Scholar
Кастрен Э. Нейротрофины как медиаторы воздействия лекарств на настроение, зависимость и нейрозащиту. Mol Neurobiol. 2004. 29 (3): 289–302.
CAS PubMed Статья Google Scholar
Думан RS, Ли Н. Нейротрофическая гипотеза депрессии: роль синаптогенеза в действиях антагонистов рецепторов NMDA. Philos Trans R Soc Lond B Biol Sci. 2012. 367 (1601): 2475–84.
CAS PubMed PubMed Central Статья Google Scholar
Думан RS, Монтеджиа LM. Нейротрофическая модель расстройств настроения, связанных со стрессом. Биол Психиатрия. 2006. 59 (12): 1116–27.
CAS PubMed Статья Google Scholar
Jacobs BL, van Praag H, Gage FH. Нейрогенез мозга взрослых и психиатрия: новая теория депрессии. Мол Психиатрия. 2000. 5 (3): 262–9.
CAS PubMed Статья Google Scholar
Miller BR, Hen R. Современное состояние нейрогенной теории депрессии и тревоги. Curr Opin Neurobiol. 2015; 30: 51–8.
CAS PubMed Статья Google Scholar
Фурнье Н.М., Думан Р.С. Роль фактора роста эндотелия сосудов в нейрогенезе гиппокампа взрослых: значение для патофизиологии и лечения депрессии. Behav Brain Res. 2012. 227 (2): 440–9.
CAS PubMed Статья Google Scholar
Кришнан В., Нестлер Э.Дж. Молекулярная нейробиология депрессии. Природа. 2008. 455 (7215): 894–902.
CAS PubMed PubMed Central Статья Google Scholar
Монтеджиа Л.М., Луикарт Б., Барро М., Теобольд Д., Малковска И., Неф С. и др. Условные нокауты нейротрофических факторов мозга показывают гендерные различия в поведении, связанном с депрессией. Биол Психиатрия. 2007. 61 (2): 187–97.
CAS PubMed Статья Google Scholar
Karege F, Vaudan G, Schwald M, Perroud N, La Harpe R. Уровни нейротрофинов в посмертном мозге жертв самоубийства и последствия прижизненной диагностики и психотропных препаратов. Brain Res Mol Brain Res. 2005. 136 (1–2): 29–37.
CAS PubMed Статья Google Scholar
Чен Б., Доулатшахи Д., Маккуин ГМ, Ван Дж. Ф., Янг LT. Повышенная иммунореактивность гиппокампа BDNF у субъектов, получавших лечение антидепрессантами.Биол Психиатрия. 2001. 50 (4): 260–5.
CAS PubMed Статья Google Scholar
Нибуя М., Моринобу С., Думан Р.С. Регулирование мРНК BDNF и trkB в головном мозге крыс с помощью хронических электросудорожных припадков и лечения антидепрессантами. J Neurosci. 1995. 15 (11): 7539–47.
CAS PubMed Google Scholar
Нестлер Э.Дж., Барро М., ДиЛеоне Р.Дж., Эйш А.Дж., Голд С.Дж., Монтеджиа Л.М.Нейробиология депрессии. Нейрон. 2002. 34 (1): 13–25.
CAS PubMed Статья Google Scholar
Li Y, Luikart BW, Birnbaum S, Chen J, Kwon CH, Kernie SG, et al. TrkB регулирует нейрогенез гиппокампа и регулирует чувствительность к антидепрессивному лечению. Нейрон. 2008. 59 (3): 399–412.
CAS PubMed PubMed Central Статья Google Scholar
Сайранен М., Лукас Дж., Эрнфорс П., Кастрен М., Кастрен Э. Нейротрофический фактор головного мозга и антидепрессанты имеют разные, но скоординированные эффекты на обновление нейронов, пролиферацию и выживаемость в зубчатой извилине у взрослых. J Neurosci. 2005. 25 (5): 1089–94.
CAS PubMed Статья Google Scholar
Монтеджия Л.М., Барро М., Пауэлл С.М., Бертон О., Галанис В., Джемелли Т. и др. Существенная роль нейротрофического фактора головного мозга в функции гиппокампа взрослых.Proc Natl Acad Sci U S. A. 2004; 101 (29): 10827–32.
CAS PubMed PubMed Central Статья Google Scholar
Ван Дж.В., Дэвид Дж. Д., Монктон Дж. Э., Батталья Ф. Хен Ра. Хронический флуоксетин стимулирует созревание и синаптическую пластичность гранулярных клеток гиппокампа у взрослых. J Neurosci. 2008. 28 (6): 1374–84.
CAS PubMed Статья Google Scholar
Castren E, Voikar V, Rantamaki T. Роль нейротрофических факторов в депрессии. Curr Opin Pharmacol. 2007. 7 (1): 18–21.
CAS PubMed Статья Google Scholar
Рантамаки Т., Хендолин П., Канкаанпа А., Миятович Дж., Пиппонен П., Доменичи Е. и др. Фармакологически разнообразные антидепрессанты быстро активируют рецептор нейротрофического фактора мозга TrkB и индуцируют сигнальные пути фосфолипазы-Cgamma в мозге мышей.Нейропсихофармакология. 2007. 32 (10): 2152–62.
CAS PubMed Статья Google Scholar
Warner-Schmidt JL, Duman RS. VEGF является важным медиатором нейрогенного и поведенческого действия антидепрессантов. Proc Natl Acad Sci U S. A. 2007; 104 (11): 4647–52.
CAS PubMed PubMed Central Статья Google Scholar
Fujioka T, Fujioka A, Duman RS.Активация передачи сигналов цАМФ способствует морфологическому созреванию новорожденных нейронов в гиппокампе взрослых. J Neurosci. 2004. 24 (2): 319–28.
CAS PubMed Статья Google Scholar
Сираяма Ю., Чен А.С., Накагава С., Рассел Д.С., Думан Р.С. Нейротрофический фактор головного мозга оказывает антидепрессивное действие в поведенческих моделях депрессии. J Neurosci. 2002. 22 (8): 3251–61.
CAS PubMed Google Scholar
Доу А.Л., Рассел Д.С., Думан Р.С. Регулирование мРНК активина и фосфорилирования Smad2 при лечении антидепрессантами в мозге крысы: эффекты в поведенческих моделях. J Neurosci. 2005. 25 (20): 4908–16.
CAS PubMed Статья Google Scholar
Боррото-Эскуэла Д.О., Ромеро-Фернандес В., Мудо Дж., Перес-Алеа М., Сируэла Ф., Тараканов А.О. и др. Рецептор фактора роста фибробластов 1- 5-гидрокситриптамин 1A гетерорецепторные комплексы и их усиление пластичности гиппокампа.Биол Психиатрия. 2012. 71 (1): 84–91.
CAS PubMed Статья Google Scholar
Ганеа К., Менке А., Шмидт М.В., Лука С., Раммес Дж., Либл С. и др. Конвергентные данные, полученные от животных и людей, предполагают, что путь активин / ингибин участвует в антидепрессивном ответе. Перевод Психиатрия. 2012; 2: e177.
CAS PubMed PubMed Central Статья Google Scholar
Грин Дж, Банаср М, Ли Б., Уорнер-Шмидт Дж, Думан Р.С. Передача сигналов фактора роста эндотелия сосудов необходима для поведенческих действий лечения антидепрессантами: фармакологическая и клеточная характеристика. Нейропсихофармакология. 2009. 34 (11): 2459–68.
CAS PubMed PubMed Central Статья Google Scholar
Hoshaw BA, Malberg JE, Lucki I. Центральное введение IGF-I и BDNF приводит к длительным эффектам, подобным антидепрессантам.Brain Res. 2005; 1037 (1-2): 204-8.
CAS PubMed Статья Google Scholar
Turner CA, Gula EL, Taylor LP, Watson SJ, Akil H. Антидепрессантоподобные эффекты внутрицеребровентрикулярного FGF2 у крыс. Brain Res. 2008; 1224: 63–8.
CAS PubMed PubMed Central Статья Google Scholar
Warner-Schmidt JL, Duman RS. VEGF как потенциальная мишень для терапевтического вмешательства при депрессии.Curr Opin Pharmacol. 2008; 8 (1): 14–9.
CAS PubMed Статья Google Scholar
Siuciak JA, Lewis DR, Wiegand SJ, Lindsay RM. Антидепрессантоподобный эффект нейротрофического фактора головного мозга (BDNF). Pharmacol Biochem Behav. 1997. 56 (1): 131–7.
CAS PubMed Статья Google Scholar
Lang UE, Hellweg R, Kalus P, Bajbouj M, Lenzen KP, Sander T. и др.Связь функционального полиморфизма BDNF и тревожных черт личности. Психофармакология. 2005. 180 (1): 95–9.
CAS PubMed Статья Google Scholar
Сен С., Нессе Р.М., Столтенберг С.Ф., Ли С., Глейберман Л., Чакраварти А. и др. Вариант кодирования BDNF связан с невротизмом области личностного инвентаря NEO, фактором риска депрессии. Нейропсихофармакология. 2003. 28 (2): 397–401.
CAS PubMed Статья Google Scholar
Verhagen M, van der Meij A, van Deurzen PA, Janzing JG, Arias-Vasquez A, Buitelaar JK, et al. Мета-анализ полиморфизма BDNF Val66Met при большом депрессивном расстройстве: влияние пола и этнической принадлежности. Мол Психиатрия. 2010. 15 (3): 260–71.
CAS PubMed Статья Google Scholar
Хван Дж. П., Цай С.Дж., Хонг СиДжей, Ян СН, Лирнг Дж.Ф., Ян Ю.М. Полиморфизм Val66Met гена нейротрофического фактора головного мозга связан с гериатрической депрессией.Neurobiol Aging. 2006. 27 (12): 1834–7.
CAS PubMed Статья Google Scholar
Ига Дж., Уэно С., Ямаути К., Нумата С., Тайоши-Сибуя С., Киноути С. и др. Полиморфизм Val66Met гена нейротрофического фактора головного мозга связан с психотическими особенностями и суицидальным поведением у японских пациентов с тяжелой депрессией. Am J Med Genet B Neuropsychiatr Genet. 2007. 144B (8): 1003–6.
PubMed Статья Google Scholar
Chen ZY, Jing D, Bath KG, Ieraci A, Khan T, Siao CJ, et al. Генетический вариант полиморфизма BDNF (Val66Met) изменяет поведение, связанное с тревогой. Наука. 2006. 314 (5796): 140–3.
CAS PubMed PubMed Central Статья Google Scholar
Anttila S, Huuhka K, Huuhka M, Rontu R, Hurme M, Leinonen E, et al. Взаимодействие между генотипами 5-HT1A и BDNF увеличивает риск устойчивой к лечению депрессии. J Neural Transm (Вена).2007. 114 (8): 1065–8.
CAS Статья Google Scholar
Киши Т., Цунока Т., Икеда М., Кавасима К., Окочи Т., Китадзима Т. и др. Ген рецептора серотонина 1A и большое депрессивное расстройство: исследование ассоциации и метаанализ. J Hum Genet. 2009. 54 (11): 629–33.
CAS PubMed Статья Google Scholar
Киши Т., Йошимура Р., Фукуо Ю., Окочи Т., Мацунага С., Умене-Накано В. и др.Ген рецептора серотонина 1А придает предрасположенность к расстройствам настроения: результат расширенного метаанализа пациентов с большой депрессией и биполярным расстройством. Eur Arch Psychiatry Clin Neurosci. 2013. 263 (2): 105–18.
PubMed Статья Google Scholar
Autry AE, Monteggia LM. Нейротрофический фактор головного мозга и нервно-психические расстройства. Pharmacol Rev.2012; 64 (2): 238–58.
CAS PubMed PubMed Central Статья Google Scholar
Мартинович К., Лу Б. Взаимодействие между BDNF и серотонином: роль в расстройствах настроения. Нейропсихофармакология. 2008. 33 (1): 73–83.
CAS PubMed Статья Google Scholar
Гулд Э. Серотонин и нейрогенез гиппокампа. Нейропсихофармакология. 1999; 21 (2 доп.): 46С – 51С.
CAS PubMed Статья Google Scholar
Мосснер Р., Даниэль С., Альберт Д., Хейлс А., Окладнова О., Шмитт А. и др.Функция транспортера серотонина модулируется нейротрофическим фактором мозга (BDNF), но не фактором роста нервов (NGF). Neurochem Int. 2000. 36 (3): 197–202.
CAS PubMed Статья Google Scholar
Паскуаль-Бразо Дж., Кастро Э., Диаз А., Валдизан Э.М., Пилар-Куэльяр Ф., Видал Р. и др. Модуляция путей нейропластичности и антидепрессантоподобных поведенческих реакций после кратковременного (3 и 7 дней) введения агониста рецептора 5-HT (4) RS67333.Int J Neuropsychopharmacol. 2012; 15 (5): 631–43.
CAS PubMed Статья Google Scholar
Samarajeewa A, Goldemann L, Vasefi MS, Ahmed N, Gondora N, Khanderia C, et al. Активация рецептора 5-HT7 способствует увеличению экспрессии и фосфорилирования рецептора TrkB. Front Behav Neurosci. 2014; 8: 391.
PubMed PubMed Central Статья CAS Google Scholar
Цао Л., Цзяо Х, Цузга Д.С., Лю И, Фонг Д.М., Янг Д. и др. VEGF связывает активность гиппокампа с нейрогенезом, обучением и памятью. Нат Жене. 2004. 36 (8): 827–35.
CAS PubMed Статья Google Scholar
Боррото-Эскуэла Д.О., Тараканов А.О., Фьюкс К. Гетерорецепторные комплексы FGFR1-5-HT1A: значение для понимания и лечения большой депрессии. Trends Neurosci. 2016; 39 (1): 5–15.
CAS PubMed Статья Google Scholar
Borroto-Escuela DO, Narvaez M, Perez-Alea M, Tarakanov AO, Jimenez-Beristain A, Mudo G, et al. Доказательства существования комплексов гетерорецепторов FGFR1-5-HT1A в 5-HT системе шва среднего мозга. Biochem Biophys Res Commun. 2015; 456 (1): 489–93.
CAS PubMed Статья Google Scholar
Borroto-Escuela DO, Perez-Alea M, Narvaez M, Tarakanov AO, Mudo G, Jimenez-Beristain A, et al. Усиление передачи сигналов FGFR1 в гетерорецепторном комплексе FGFR1-5-HT1A в системах нейронов шва среднего мозга 5-HT.Актуальность для нейропластичности и депрессии. Biochem Biophys Res Commun. 2015; 463 (3): 180–6.
CAS PubMed Статья Google Scholar
Мин G-L, Сонг Х. Нейрогенез взрослых в центральной нервной системе млекопитающих. Annu Rev Neurosci. 2005; 28: 223–50.
CAS PubMed Статья Google Scholar
Сух Х., Дэн В., Гейдж Ф. Сигнализация во взрослом нейрогенезе.Annu Rev Cell Dev Biol. 2009; 25: 253–75.
CAS PubMed Статья Google Scholar
Malberg JE. Влияние нейрогенеза гиппокампа взрослых на действие антидепрессантов. J Psychiatry Neurosci. 2004. 29 (3): 196–205.
PubMed PubMed Central Google Scholar
Айран Р.Д., Мельцер Л.А., Рой М., Гонг Й., Чен Х., Дейссерот К. Высокоскоростная визуализация выявляет нейрофизиологические связи с поведением на животной модели депрессии.Наука. 2007. 317 (5839): 819–23.
CAS PubMed Статья Google Scholar
Дэвид Д. Д., Сэмюэлс Б. А., Райнер К., Ван Дж. В., Марстеллер Д., Мендес И. и др. Нейрогенез-зависимые и независимые эффекты флуоксетина в животной модели тревоги / депрессии. Нейрон. 2009. 62 (4): 479–93.
CAS PubMed PubMed Central Статья Google Scholar
Boldrini M, Santiago AN, Hen R, Dwork AJ, Rosoklija GB, Tamir H, et al. Число нейронов гранул гиппокампа и объем зубчатой извилины при большой депрессии, леченной и не леченной антидепрессантами. Нейропсихофармакология. 2013. 38 (6): 1068–77.
CAS PubMed PubMed Central Статья Google Scholar
Сахай А., Скоби К.Н., Хилл А.С., О’Кэрролл С.М., Хейрбек М.А., Бургхардт Н.С. и др. Усиления нейрогенеза в гиппокампе у взрослых достаточно для улучшения разделения паттернов.Природа. 2011. 472 (7344): 466–70.
CAS PubMed PubMed Central Статья Google Scholar
Ishizuka T, Goshima H, Ozawa A, Watanabe Y. Стимуляция рецептора 5-HT4 усиливает дифференцировку индуцированных мышами плюрипотентных стволовых клеток в нервные клетки-предшественники. Clin Exp Pharmacol Physiol. 2014; 41 (5): 345–50.
CAS PubMed Статья Google Scholar
Tamburella A, Micale V, Navarria A, Drago F. Антидепрессивные свойства частичного агониста рецептора 5-HT4, SL65.0155: поведенческие и нейрохимические исследования на крысах. Prog Neuro-Psychopharmacol Biol Psychiatry. 2009. 33 (7): 1205–10.
CAS Статья Google Scholar
Лю М.Т., Куан Ю.Х., Ван Дж., Хен Р., Гершон, доктор медицины. Опосредованная рецептором 5-HT4 нейрозащита и нейрогенез в кишечной нервной системе взрослых мышей.J Neurosci. 2009. 29 (31): 9683–99.
CAS PubMed PubMed Central Статья Google Scholar
Имото Ю., Кира Т., Сукено М., Нишитани Н., Нагаясу К., Накагава Т. и др. Роль рецептора 5-HT4 в хронической нейрогенной активности, вызванной лечением флуоксетином, и дематурации гранулярных клеток в зубчатой извилине. Мол мозг. 2015; 8: 29.
PubMed PubMed Central Статья CAS Google Scholar
Кобаяси К., Икеда Ю., Сакаи А., Ямасаки Н., Ханеда Е., Миякава Т. и др. Обратное созревание нейронов гиппокампа серотонинергическими антидепрессантами. Proc Natl Acad Sci U S. A. 2010; 107 (18): 8434–9.
CAS PubMed PubMed Central Статья Google Scholar
Карпова Н.Н., Пикенхаген А., Линдхольм Дж., Тирабоски Е., Кулеская Н., Агустсдоттир А. и др. Стирание страха у мышей требует синергии между антидепрессантами и тренировкой вымирания.Наука. 2011. 334 (6063): 1731–4.
CAS PubMed PubMed Central Статья Google Scholar
Охира К., Такеучи Р., Иванага Т., Миякава Т. Хроническое лечение флуоксетином снижает экспрессию парвальбумина и перинейрональные сети в гамма-аминомасляных ацидергических интернейронах лобной коры у взрослых мышей. Мол мозг. 2013; 6: 43.
PubMed PubMed Central Статья CAS Google Scholar
Ли Д.А., Блэкшоу С. Функциональные последствия гипоталамического нейрогенеза в мозге взрослых млекопитающих. Int J Dev Neurosci. 2012; 30 (8): 615–21.
PubMed PubMed Central Статья Google Scholar
Гулд Э. Насколько широко распространен нейрогенез взрослых у млекопитающих? Nat Rev Neurosci. 2007. 8 (6): 481–8.
CAS PubMed Статья Google Scholar
Охира К., Такеучи Р., Сёдзи Х., Миякава Т. Флуоксетин-индуцированный кортикальный нейрогенез взрослых. Нейропсихофармакология. 2013. 38 (6): 909–20.
CAS PubMed PubMed Central Статья Google Scholar
Люшер Б., Шен К., Сахир Н. Гипотеза ГАМКергического дефицита основного депрессивного расстройства. Мол Психиатрия. 2011. 16 (4): 383–406.
CAS PubMed Статья Google Scholar
Burghardt NS, Park EH, Hen R, Fenton AA. Нейроны гиппокампа, рожденные взрослыми, способствуют когнитивной гибкости у мышей. Гиппокамп. 2012. 22 (9): 1795–808.
PubMed PubMed Central Статья Google Scholar
Дрю Л.Дж., Хейрбек М.А., Луна В.М., Денни К.А., Клоидт М.А., Ву М.В. и др. Активация локальных тормозных цепей в зубчатой извилине нейронами, рожденными взрослыми. Гиппокамп. 2016; 26 (6): 763–78.
CAS PubMed PubMed Central Статья Google Scholar
Икрар Т., Го Н., Хе К., Беснард А., Левинсон С., Хилл А. и др. Взрослый нейрогенез изменяет возбудимость зубчатой извилины. Передние нервные цепи. 2013; 7: 204.
PubMed PubMed Central Статья CAS Google Scholar
Lacefield CO, Itskov V, Reardon T, Hen R, Gordon JA. Влияние образовавшихся взрослых гранулярных клеток на скоординированную сетевую активность в зубчатой извилине. Гиппокамп. 2012. 22 (1): 106–16.
PubMed Статья Google Scholar
Fanselow MS, Донг HW. Различны ли функционально дорсальный и вентральный гиппокамп? Нейрон. 2010. 65 (1): 7–19.
CAS PubMed PubMed Central Статья Google Scholar
Pothuizen HH, Zhang WN, Jongen-Relo AL, Feldon J, Yee BK. Диссоциация функций спинного и вентрального гиппокампа в способности крысы к пространственному обучению: внутрисубъектное, внутризадачное сравнение эталонной и рабочей пространственной памяти.Eur J Neurosci. 2004. 19 (3): 705–12.
PubMed Статья Google Scholar
Хейрбек М.А., Дрю Л.Дж., Бургхардт Н.С., Костантини Д.О., Танненхольц Л., Ахмари С.Е. и др. Дифференциальный контроль обучения и тревоги вдоль дорсовентральной оси зубчатой извилины. Нейрон. 2013. 77 (5): 955–68.
CAS PubMed PubMed Central Статья Google Scholar
Bagot RC, Parise EM, Pena CJ, Zhang HX, Maze I, Chaudhury D, et al. Вентральные афференты гиппокампа к прилежащему ядру регулируют предрасположенность к депрессии. Nat Commun. 2015; 6: 7062.
CAS PubMed PubMed Central Статья Google Scholar
Тай К.М., Пракаш Р., Ким С.И., Фенно Л.Е., Гросеник Л., Зараби Н. и др. Схема миндалевидного тела, обеспечивающая обратимый и двунаправленный контроль тревоги. Природа. 2011. 471 (7338): 358–62.
CAS PubMed PubMed Central Статья Google Scholar
Padilla-Coreano N, Bolkan SS, Pierce GM, Blackman DR, Hardin WD, Garcia-Garcia AL, et al. Прямой вентральный гиппокампально-префронтальный вход необходим для нервной активности и поведения, связанных с тревогой. Нейрон. 2016. 89 (4): 857–66.
Wu MV, Sahay A, Duman RS, Hen R. Функциональная дифференцировка нейронов, рожденных взрослыми, вдоль перегородочной оси зубчатой извилины.Cold Spring Harb Perspect Biol. 2015; 7 (8): a018978.
PubMed Статья CAS Google Scholar
Gage FH, Thompson RG, Valdes JJ. Эндогенный норэпинефрин и серотонин в формации гиппокампа во время развития и восстановления после гиперреактивности перегородки. Pharmacol Biochem Behav. 1978. 9 (3): 359–67.
CAS PubMed Статья Google Scholar
Границы | Роль N-арахидоноил-серотонина (AA-5-HT) в архитектуре цикла сна-бодрствования, гомеостазе сна и нейротрансмиттерах Регламент
Введение
Эндоканнабиноидная система представляет собой сложную биологическую структуру, которая выполняет множество модулирующих функций (Kendall and Yudowski, 2016; Ligresti et al., 2016; Аргуэта и ДиПатрицио, 2017; Bennett et al., 2017; Дос Аньос-Гарсия и др., 2017; Sun et al., 2017). Компоненты эндоканнабиноидной системы включают наиболее изученные эндоканнабиноиды анандамид (AEA) и 2-арахидоноилглицерин (2-AG; Di Marzo and Piscitelli, 2015; Yuan et al., 2016; Biernacki and Skrzydlewska, 2016; Correa et al., 2016). ; Huang et al., 2016; Lu and Mackie, 2016; Fakhoury, 2017). Эндоканнабиноиды связываются с трансмембранными белками, обозначенными каннабиноидными рецепторами CB 1 и CB 2 (Kendall and Yudowski, 2016; Kruk-Slomka et al., 2016; Чен и др., 2017; Фитцпатрик и Даунер, 2017; Маллипедди и др., 2017). Кроме того, сообщалось, что основными ферментами, ответственными за гидролиз AEA и 2-AG, являются амидгидролаза жирных кислот (FAAH) или моноацилглицеринлипаза (MAGL), соответственно (Piomelli, 2014; Biernacki and Skrzydlewska, 2016; Janssen and van дер Стелт, 2016). Наконец, несколько доказательств показали, что мембранный транспортер анандамида (AMT) также играет ключевую роль в модулировании множества биологических функций (Leung et al., 2013; Николусси и Герч, 2015). Среди разнообразных результатов, которые предполагают участие эндоканнабиноидной системы в регуляции нескольких нейробиологических явлений, различные отчеты показали, что цикл сна-бодрствования, вероятно, находится под контролем этой системы (Santucci et al., 1996; Murillo-Rodríguez et al., 1998, 2001, 2003, 2008a, 2011a, 2013, 2016; Herrera-Solis et al., 2010; Rueda-Orozco et al., 2010; Pava et al., 2014, 2016). В связи с недавними результатами, касающимися взаимосвязи между AEA и членом 1 подсемейства катионного канала временного рецептора (TRPV1), также известного как рецептор капсаицина или ваниллоидный рецептор 1, возрастающий научный интерес к нейромолекулярным свойствам TRPV1 (Tóth et al., 2009; Лоуин и Штрауб, 2015; Чен и др., 2016; Киркедал и др., 2016). Экспериментальный подход, направленный на описание нейромолекулярной роли TRPV1, включал фармакологические средства с использованием N -арахидоноилсеротонина (AA-5-HT). Например, сообщалось, что этот препарат является двойным блокатором FAAH / TRPV1, вызывая анксиолитические эффекты у мышей (Micale et al., 2009). Более того, различные дозы AA-5-HT (0–0,5 нмоль), вводимые в базолатеральную миндалину перед тестированием в приподнятом крестообразном лабиринте, увеличивали время пребывания в открытых рукавах (John and Currie, 2012).Кроме того, было показано, что AA-5-HT вызывает противосудорожные эффекты (Micale et al., 2009; Vilela et al., 2014), снижает депрессивно-подобное поведение (Navarria et al., 2014; Kirkedal et al., 2016) и способствует восстановлению после стресса (Sartim et al., 2017). Несмотря на накопление данных о нейробиологической роли AA-5-HT, нет данных о том, может ли это соединение модулировать цикл сна-бодрствования. Таким образом, мы исследовали эффекты AA-5-HT (5, 10, 20 мг / кг, i.p.) на сон, а также внеклеточное содержание нейротрансмиттеров, связанных с циклом сна-бодрствования, таких как дофамин (DA), норадреналин (NE), адреналин (EP), серотонин (5-HT), а также аденозин (AD). Более того, чтобы предоставить дополнительные доказательства относительно предполагаемых свойств AA-5-HT вызывать сон, мы проверили, будет ли этот препарат блокировать бдительность, вызванную фитоканнабиноидом каннабидиолом (CBD, 30 мг / кг, внутрибрюшинно; Murillo-Rodríguez et al. , 2006, 2008b, 2011b; Kim, 2012) или стимулирующий препарат модафинил (MOD, 30 мг / кг, i.п.; Сухал и др., 2015; Wang et al., 2015; Барато и др., 2016). Кроме того, роль AA-5-HT в гомеостазе сна была изучена у животных, получавших CBD или MOD, которые были лишены сна. Наше исследование предоставляет новые данные о возможной роли AA-5-HT в модуляции сна.
Материалы и методы
Эксперимент 1: Влияние на сон после инъекций AA-5-HT во время периода включения или выключения света
Подопытные животные
крыс-самцов линии Wistar ( n = 50; 250–300 г) содержали по одному в поликарбонатных клетках (48.26 см × 26,67 см × 20,32 см; Harlan Laboratories. México) при цикле свет-темнота (включение: 07: 00–19: 00 ч), контролируемой влажности (60 ± 10%) и постоянной температуре (21 ± 1 ° C). Крысы имели свободный доступ к Purina Rat Chow (Purina, México), а также к водопроводной воде. Протоколы экспериментов были одобрены Комитетом по исследованиям и этике нашего университета в соответствии с национальными и международными стандартами защиты животных, включая мексиканские стандарты использования и обращения с лабораторными животными (DOF.NOM-062-Z00-1999), а также Национальные институты здравоохранения (публикация NIH № 80–23, пересмотренная в 1996 г.). В течение всего исследования принимались во внимание усилия по минимизации страданий животных, и по этическим соображениям использовалось меньшее количество животных.
Химическая промышленность
AA-5-HT был предоставлен профессором Винченцо Ди Марцо, и химическое вещество было растворено в растворе носителя (VEH), состоящем из полиэтиленгликоля / физиологического раствора (5:95 об. / Об.), Как описано ранее (Murillo-Rodriguez et al. , 2003, 2011а, 2013).Все реагенты, химикаты и материалы были приобретены у Sigma – Aldrich (Сент-Луис, Миссури, США).
Операции по записи сна
Животных ( n = 25) анестезировали смесью ацепромазина (0,75 мг / кг; внутрибрюшинно), ксилазина (2,5 мг / кг; внутрибрюшинно) и кетамина (22 мг / кг; внутрибрюшинно) и помещали в стереотаксическую среду. рама (David Kopf Instruments, Tujunga, CA, США) для имплантации стандартных электродов для записи сна. Вкратце, два винтовых электрода из нержавеющей стали были вставлены на 2 мм с каждой стороны сагиттального синуса и на 3 мм кпереди от Bregma (лобной коры), тогда как два других винта были расположены на 3 мм с каждой стороны от сагиттального синуса и в 6 мм позади Bregma ( затылочная кора).Эта установка электрода регистрировала сигнал электроэнцефалограммы (ЭЭГ) путем получения биполярного (дифференциального) сигнала ЭЭГ, записанного с двух контралатеральных винтовых электродов (лобно-затылочный). Кроме того, запись электромиограммы (ЭМГ) заключалась в сигнале, полученном имплантацией двух проволочных электродов в спинные мышцы шеи каждой крысы. Затем провода ЭЭГ / ЭМГ были вставлены в шестиконтактную пластиковую заглушку (Plastics One, Роанок, Вирджиния, США), и вся установка электрода была прикреплена к черепу с помощью промышленного стоматологического цемента.По завершении операций ЭЭГ / ЭМГ всех животных поместили в индивидуальные клетки с обычной подстилкой и стандартными контейнерами для воды и корма на период послеоперационного восстановления. Крысы с имплантированными электродами были подключены к 6-канальному коммутатору с контактными кольцами через кабель длиной 50 см (Plastics One, Роанок, Вирджиния, США), который позволял животным свободно вращаться и перемещаться в боксе в течение всего исследования. . Хирургическая процедура, а также условия привыкания (в течение 7 дней) были выполнены, как описано в других источниках (Murillo-Rodríguez et al., 2011б).
Анализ записей сна
Сигналы ЭЭГ / ЭМГ оценивались в 12-секундных периодах, что, в свою очередь, позволяет охарактеризовать цикл сна-бодрствования в W, SWS или REMS с помощью программы оценки сна (ICELUS). Подробно: Провода ЭЭГ / ЭМГ были подключены к системе контактных колец (Plastics One, Роанок, Вирджиния, США) и к усилителю, позволяющему фильтровать сигнал ЭЭГ с частотой 70 Гц (фильтр нижних частот) и 0,3 Гц ( фильтр верхних частот; модель M15LT 15A54; Grass Instruments. Куинси, Массачусетс, США).Сигналы ЭЭГ непрерывно регистрировались с частотой 128 Гц с помощью 100-битной платы аналого-цифрового преобразователя (многофункциональная плата ввода-вывода NI PCI-6033E и программное обеспечение NI-DAQ, экранированный соединительный блок SCB-100. National Instruments. Austin, Техас, США). С помощью программного обеспечения для оценки сна (ICELUS) цикл сна и бодрствования был охарактеризован следующим образом: наличие десинхронизированной ЭЭГ, а также высокая активность ЭМГ относятся к определению W, тогда как медленные волны высокой амплитуды с низкой ЭМГ тон по отношению к бодрствованию относятся к критериям SWS.Наконец, REMS характеризовался регулярной тета-активностью на ЭЭГ в сочетании с низкой ЭМГ, сопровождаемой миоклоническими особенностями. Процедуры записи сна выполнялись, как сообщалось ранее (Мурильо-Родригес и др., 2006, 2007b, 2008b, c; Mijangos-Moreno et al., 2016).
Фармакологические управления
Сразу после 7 дней после операции и соответствующего периода привыкания животных помещали в один из экспериментальных блоков цикла свет-темнота: период включения света (07: 00–19: 00 ч) или период отключения света (19 часов). : 00–07: 00 ч).Согласно соответствующему экспериментальному блоку свет-темнота, животные были отключены от системы регистрации сна, и экспериментальные испытания проводились случайным образом следующим образом: VEH ( n = 5), AA-5-HT (5, 10, 20 мг / кг). Кг, ip; n = 5 в каждой группе). Независимо от того, может ли инъекция VEH изменить цикл сна-бодрствования, дополнительная группа, названная «фиктивная» ( n = 5), была включена в период светлого-темного. В эту группу вошли животные, получившие вставку (т.p.) иглы, но введение не было. После проведения экспериментов животных снова подключили к системе регистрации сна, и данные о сне были собраны в течение следующих 4 часов. В текущем эксперименте лечение проводилось случайным образом с использованием одинарного слепого метода эксперимента по латинскому квадрату.
Анализ спектра мощности
Помимо влияния AA-5-HT на сон, был проведен анализ быстрого преобразования Фурье во время цикла включения или выключения света после соответствующих обработок для анализа качественных параметров сна, таких как альфа-, дельта- и тета-спектры мощности.В нескольких отчетах было высказано предположение, что спектры мощности являются критическим элементом для определения расхождений между объективными и субъективными показателями цикла сна и бодрствования (Tobler and Borbély, 1990; Franken et al., 1991; Corsi-Cabrera et al., 2001; Christensen et al. ., 2015; Jing et al., 2016; Imperatori et al., 2016; Svetnik et al., 2017). Таким образом, следуя стандартизированным процедурам, спектры мощности для альфа (во время W, α = 8–12 Гц), дельты (по SWS, Δ = 0,5–4,0 Гц) или тета (для REMS, = 6,0–12,0 Гц) были проанализированы после экспериментальные испытания, о которых сообщалось ранее (Murillo-Rodríguez et al., 2011а).
Статистический анализ
Данные исследований сна и экспериментов со спектрами мощности были представлены как среднее ± стандартная ошибка среднего. Статистические различия определяли с помощью однофакторного дисперсионного анализа (ANOVA) с последующим апостериорным тестом Scheffé для множественных сравнений между экспериментальными группами. Все статистические анализы выполнялись с использованием StatView (версия 5.0.0, Институт SAS, США), и статистические различия между группами определялись, если P <0.05.
Эксперимент 2: Влияние на внеклеточные уровни моноаминов или AD после введения AA-5-HT во время периода выключения света
Экспериментальные животные, химикаты и фармакологические администрации
Как описано в эксперименте 1.
Операции микродиализа
Множественные эксперименты подтвердили ключевую роль прилежащего ядра (AcbC) в модуляции сна (Lazarus et al., 2012, 2013; Qiu et al., 2012; Zhang et al., 2013; Liu et al., 2016). Кроме того, предыдущие исследования нашей лаборатории показали надежные измерения моноаминов, а также AD от AcbC (Murillo-Rodríguez et al., 2006; Mijangos-Moreno et al., 2014, 2016). Таким образом, эти доказательства позволили нам собрать образцы микродиализа из AcbC, чтобы определить, могут ли изменения сна, вызванные AA-5-HT, быть связаны с эффектами в содержимом нейромедиаторов. Для достижения этой цели новую группу животных ( n = 25) под наркозом поместили в стереотаксическую рамку (David Kopf Instruments.Tujunga, Калифорния, США) для имплантации направляющей канюли для микродиализа (руководство IC; BioAnalytical Systems [BAS], West Lafayette, IN, United States). Канюлю односторонне помещали в AcbC (координаты: A = +1,2 и L = +2,0, H = –7,0 мм, со ссылкой на Bregma [Paxinos and Watson, 2005]), и сразу после операции крыс по отдельности помещали в чаша для микродиализа (автономная система Raturn Microdialysis. MD-1404, BAS. West Lafayette, IN, США) для восстановления, а также привыкания к экспериментальным условиям в течение 7 дней.Все хирургические процедуры с использованием зондов для микродиализа были выполнены, как сообщалось ранее (Мурильо-Родригес и др., 2007a; Миджангос-Морено и др., 2014, 2016).
Процедуры отбора проб для микродиализа
Через семь дней после операции микродиализа крыс удаляли из чашек для микродиализа и извлекали стилет из направляющей канюли. Позже зонд для микродиализа (длина 1 мм; полиакрилонитрил, MWCO = 30 000 дальтон; OD 340 мкм; BAS. West Lafayette, IN, США) был вставлен в 07:00.Затем через миниатюрную трубку (наружный диаметр 0,65 мм × внутренний диаметр 0,12 мм; BAS, West Lafayette, IN, США), присоединенный к шприцу объемом 2,5 мл (BAS, West Lafayette, IN, США), и с помощью насоса (скорость потока: 0,25 мкл / мин; BAS Bee, West Lafayette, IN, США) вводили искусственную спинномозговую жидкость [aCSF (состав: NaCl 147 мМ, KCl 3 мМ, CaCl 1,2 мМ, MgCl 1,0 мМ, pH 7,2)]. Важно отметить, что предыдущие эксперименты показали, что после введения зонду для микродиализа требуется примерно 6-7 часов для стабилизации (Porkka-Heiskanen et al., 2000; Мурильо-Родригес и др., 2004). Однако в текущем отчете датчики были стабилизированы в течение 24 часов. Позже, по достижении периода стабилизации, образцы для микродиализа собирали ежечасно через 4 часа у каждого животного после экспериментальных испытаний. Затем диализаты усредняли по четырем временным точкам как общее значение для каждого соединения в каждом экспериментальном состоянии [Sham ( n = 5), VEH ( n = 5), AA-5-HT (5, 10, или 20 мг / кг, отдельно все дозы ( n = 5, каждая доза))].Стоит отметить, что зонды для микродиализа использовались не более 5 дней, так как предыдущие отчеты показали, что последующие вставки, глиоз значительно снижает способность мембраны переносить жидкость через поры (Porkka-Heiskanen et al., 2000; Murillo-Rodriguez и др., 2004). Дополнительный тест на восстановление микродиализных зондов заключался во введении зондов в тестовый раствор, содержащий внешние известные концентрации моноаминов или AD с перфузией aCSF при рабочей скорости потока (0.25 мкл / мин). Микродиализаты собирали в этих экспериментальных условиях после трех повторностей с использованием другой партии зондов для микродиализа. Каждый образец, собранный в ходе исследования восстановления in vitro , был проанализирован, и соотношение площадей пиков было рассчитано относительно известных стандартов представляющих интерес нейромедиаторов. Для получения скорости восстановления данные рассчитывали следующим образом: Степень извлечения (%) = (отношение площадей пиков образца из микродиализного образца) / (отношение площадей пиков образца в исследуемом растворе).Все процедуры микродиализа выполнялись, как и в предыдущих отчетах (Porkka-Heiskanen et al., 2000; Blanco-Centurion et al., 2006; Mijangos-Moreno et al., 2014, 2016; Murillo-Rodríguez et al., 2016).
Фармакологический вызов
Через семь дней после операции микродиализа и через 24 часа после стабилизации микродиализных зондов животным были предложены следующие экспериментальные задачи: из-за сна, индуцированного AA-5-HT (см. Раздел «Результаты»), животных отключили от системы микродиализа и назначили лечение случайным образом. в период отключения света.Сразу после проведения экспериментальных испытаний крыс снова подключили к системе микродиализа, поместили их обратно в чаши для микродиализа и через 4 часа собирали образцы. Позже все образцы хранили (-80 ° C) для дальнейшего анализа.
Процедура высокоэффективной жидкостной хроматографии (ВЭЖХ)
Собранные диализаты вводили в ВЭЖХ для обнаружения и количественного определения DA, NE, EP, 5-HT и AD. Подробно: диализаты фильтровали (Millipore 0,22 мкм; Merck Millipore.Дармштадт, Германия) и вводили в ВЭЖХ (Modular Prominence, Shimadzu, Япония). Подвижная фаза для моноаминов состояла из мононатрийфосфата [(7 мМ, pH 3,0) плюс метанол (3,5%)], перфузируемого насосом (LC-20AT, Shimadzu, Япония) при скорости потока 80 мкл / мин. Разделение молекул было достигнуто с помощью микрокапиллярной колонки [октадециловый диоксид кремния (3 мкм, 100 × 1 мм), BAS, West Lafayette, IN, США] с контролем температуры (22 ° C; печь CTO-20A. Shimadzu, Япония). . Образцы вводились автоматически (SIL-20A HT Prominence HPLC, Shimadzu.Япония) в ВЭЖХ, а хроматографические данные сохраняли на персональном компьютере (через компьютерный контроллер CBM-20A, Shimadzu, Japan). Для определения концентраций DA, NE, EP, 5-HT готовили серию разведений известных внешних стандартов этих нейромедиаторов и вводили в ВЭЖХ. Чтобы гарантировать специфичность метода ВЭЖХ, были проведены дополнительные процедуры, как было предложено производителем, а также сообщалось другими (Sartori et al., 2008; Mazzucchelli et al., 2011). Моноамины были обнаружены с помощью электрохимического детектора (LC-4C; BAS, West Lafayette, IN, США), как описано другими (Gold et al., 2011; Grupe et al., 2013). С другой стороны, для обнаружения AD требовалась подвижная фаза, приготовленная из 10 мМ дигидрофосфата натрия (pH 4,5) и метанола (9%), вводимая при скорости потока 80 мкл / мин с использованием насоса (LC-20AT. Prominence HPLC, Shimadzu , Япония). Обнаружение AD осуществляли с помощью УФ-детектора (SPD-20A Prominence. Shimadzu, Япония), настроенного на длину волны 254 нм (дейтериевая лампа). Все хроматографические данные для обнаружения AD регистрировались на персональном компьютере, а высоты пиков AD в диализатах сравнивались с внешними стандартами с использованием программного обеспечения для отчетов хроматографа (LC Solution, Shimadzu, Япония).Процедура обнаружения и измерения моноаминов или AD с использованием средств ВЭЖХ была разработана в соответствии с предыдущими отчетами (Murillo-Rodríguez et al., 2016).
Гистологическая проверка местоположения зонда
После экспериментов по микродиализу всех крыс умерщвляли летальной дозой пентобарбитала для стандартной процедуры перфузии сосудов. Животным проводили транскардиальную перфузию через желудочковый катетер, помещенный в левый желудочек. Далее было рассечено правое предсердие и солевой раствор (0.9%) перфузировали, а затем формальдегид (4%). Головной мозг удаляли и фиксировали в течение ночи в формальдегиде (4%) с последующим добавлением 10, 20 и 30% сахарозы / PBS (0,1 М) в течение 24 часов для каждой концентрации. Наконец, все мозги были разрезаны на коронковые срезы (20 мкм) с использованием портативного настольного криостата (Leica CM1100, Германия) и собраны в последовательном порядке 1: 5. Одна серия была использована для определения местоположения зонда и была идентифицирована путем построения графика с использованием атласа мозга крысы (Paxinos and Watson, 2005).
Статистический анализ
Как описано в Эксперименте 1.
Эксперимент 3: AA-5-HT блокирует побуждающие свойства каннабидиола или модафинила во время периода включения света
Экспериментальные животные, химикаты и фармакологические администрации
Как описано в эксперименте 1.
Фармакологические управления
Для получения дополнительных доказательств способности AA-5-HT вызывать сон, мы проверили, может ли это соединение блокировать бодрствование, вызванное фармакологическими средствами. В этом разделе исследования вводили, как описано в эксперименте 1.Однако мы включили в этот раздел исследования наивысшую дозу AA-5-HT (20 мг / кг; внутрибрюшинно), а также вызывающее пробуждение лекарство каннабидиол (CBD, 30 мг / кг; внутрибрюшинно) или стимулятор. соединение модафинил (MOD, 30 мг / кг; внутрибрюшинно). Чтобы оценить, будет ли AA-5-HT предотвращать бдительность, вызванную CBD или MOD, был введен модулятор FAAH / TRPV1 (5, 10 или 20 мг / кг; n = 5, каждая группа) за 15 минут до инъекции. либо MOD, либо CBD (30 мг / кг; каждый отдельно; n = 5, каждая группа).
Анализ спектра мощности и статистический анализ
Анализ спектров мощности после одновременного введения либо AA-5-HT + CBD, либо AA-5-HT + MOD и дальнейший статистический анализ выполняли, как описано в эксперименте 1.
Эксперимент 4: Воздействие на внеклеточные уровни моноаминов или AD, вызванное каннабидиолом или модафинилом во время периода включения света, предотвращается инъекцией AA-5-HT
Экспериментальные животные, химикаты, операции микродиализа, процедуры отбора проб микродиализа, процедуры ВЭЖХ, гистологическая проверка местоположения зонда и статистический анализ
Как описано в экспериментах 2 и 3.Для исследований, когда AA-5-HT вводили перед инъекцией CBD или MOD, статистические различия определяли с помощью двухфакторного дисперсионного анализа с последующим апостериорным тестом Шеффе для множественных сравнений между экспериментальными группами. Все статистические анализы выполнялись с использованием StatView (версия 5.0.0, Институт SAS, США), и статистические различия между группами определялись, если P <0,05.
Фармакологические проблемы
Анализ внеклеточных уровней DA, NE, EP, 5-HT и AD после совместного введения либо AA-5-HT + MOD, либо AA-5-HT + CBD проводили, как описано в экспериментах 3 и 4.
Эксперимент 5: блокада восстановления сна, вызванная каннабидиолом или модафинилом, после полной депривации сна предотвращается AA-5-HT
Экспериментальные животные, химические препараты и операции по записи сна
Как описано в экспериментах 1–4.
Полная процедура лишения сна
Продолжительное бодрствование осуществлялось путем поддержания крыс в постоянной непрерывной бдительности в течение 6 часов в течение периода включения света (с 07:00 до 13:00) путем помещения новых предметов в клетки, генерирования звуков и ударов по внешним стенкам клетки. клетки и др.(Mijangos-Moreno et al., 2015). Во время полной депривации сна (TSD, 6 ч) данные о сне постоянно записывались и анализировались, как сообщалось (Murillo-Rodríguez et al., 2016).
Фармакологические управления
Чтобы проверить, будет ли AA-5-HT предотвращать блокирующий эффект на восстановление сна после продолжительного бодрствования у крыс, получавших CBD или инъецированных MOD, сразу после окончания TSD крысы были отключены от системы записи сна и были проведены экспериментальные испытания. вводили случайным образом в течение периода включения света следующим образом: Sham ( n = 5), VEH ( n = 5), AA-5-HT (5, 10, 20 мг / кг, т.е.п.; n = 5, каждое соединение), CBD (30 мг / кг, внутрибрюшинно), MOD (30 мг / кг, внутрибрюшинно), AA-5-HT (20 мг / кг, внутрибрюшинно) и через 15 минут CBD (30 мг / кг, внутрибрюшинно). мг / кг, внутрибрюшинно) или MOD (30 мг / кг, внутрибрюшинно). После выполнения инъекций крыс снова подключили к системе регистрации сна, и данные о сне были собраны в течение следующих 4 часов.
Спектры мощности и статистический анализ
Как описано в эксперименте 4.
Эксперимент 6. Блокада компенсирующего восстановления внеклеточных уровней моноаминов или AD после полной депривации сна каннабидиолом или модафинилом предотвращается AA-5-HT в течение периода включения света
Экспериментальные животные, химические вещества, операции микродиализа, процедуры отбора проб микродиализа, процедуры ВЭЖХ, гистологическая проверка местоположения зонда, полное лишение сна, процедура фармакологических проблем и статистический анализ
Как описано в экспериментах 1–5.
Результаты
Эксперимент 1: Влияние на сон после инъекций AA-5-HT во время периода включения или выключения света
В первом эксперименте AA-5-HT, введенный в период включения света, не вызывал значительных различий в цикле сна-бодрствования между экспериментальными группами (W: F (4,20) = 2,062, P = 0,1; SWS: F (4,20) = 1,561, P = 0,2; REMS: F (4,20) = 1.614, P = 0,2; Рисунки 1A – C соответственно). Однако, AA-5-HT, введенный в начале периода выключения света, уменьшил W ( F (4,20) = 7 ,991, P < 0,0005) и улучшил SWS ( F (4,20) = 9,425 , P < 0,05), а также REMS ( F (4,20) = 8,324 , P < 0,0004, рисунки 1D – F соответственно). Апостериорный тест Scheffé показал статистические различия между имитацией / наполнителем и AA-5-HT (5, 10, 20 мг / кг) для пробуждения, SWS и REMS в период выключения света ( P <0.01). Из полученных данных мы заключаем, что AA-5-HT вызывал стимуляцию засыпания при введении в период включения света.
РИСУНОК 1. Влияние на общее время (4 часа записи сна) бодрствования (W), медленного сна (SWS) и сна с быстрым движением глаз (REMS) после экспериментальных испытаний: имитация, носитель или AA-5 -HT (5, 10 или 20 мг / кг; внутрибрюшинно). Никаких эффектов не наблюдалось в W, SWS или REMS, когда AA-5-HT вводили в период включения света ( A – C , соответственно).Однако при введении в начале периода выключения AA-5-HT уменьшал W и увеличивал SWS и REMS ( D – F , соответственно; Среднее ± SEM ∗ по сравнению с имитацией / носителем, P <0,05; & по сравнению с AA-5-HT-5, P <0,05; # по сравнению с AA-5-HT-10, P <0,05). Сокращения: бодрствование (W), медленный сон (SWS) и сон с быстрым движением глаз (REMS), AA-5-HT-5 мг / кг (AA-5-HT-5), AA-5-HT-10 мг. / Кг (AA-5-HT-10), AA-5-HT-20 мг / кг (AA-5-HT-20).
Что касается анализа спектров мощности (альфа, полученная во время W, дельта, полученная во время SWS, и тета, зарегистрированная во время REMS), мы обнаружили, что системные инъекции во время периода включения различных доз AA-5-HT (5, 10 или 20 мг / кг, i.p.) не изменяли значения для альфа ( F (4,20) = 0,964, P = 0,4) или тета ( F (4,20) = 1,524, P = 0,2), тогда как увеличенные дельта-спектры мощности ( F (4,20) = 3,053, P < 0,02; рисунки 2A – C, соответственно). Когда AA-5-HT вводили в начале периода выключения, было обнаружено дозозависимое снижение альфа ( F (4,20) = 3,12 , P < 0.03), при этом наблюдалось увеличение дельта ( F (4,20) = 3,377 , P < 0,02), а также дозозависимое усиление спектров тета-мощности ( F (4,20 ) = 3,456, P < 0,02; рисунки 2D – F соответственно). Апостериорный тест Scheffé показал межгрупповые различия между имитацией / наполнителем и AA-5-HT (5, 10, 20 мг / кг) для альфа-, дельта- и тета-спектров мощности ( P <0,02). Таким образом, мы обнаружили, что AA-5-HT вызывает дозозависимый эффект в спектрах мощности сна за счет уменьшения альфа и усиления спектров мощности тета.Мы пришли к выводу, что уменьшение, наблюдаемое при бодрствовании, вероятно, соответствует уменьшению, обнаруженному в альфа-спектрах мощности. И наоборот, усиление SWS и REMS, вызванное AA-5-HT, сопровождается увеличением дельта- и тета-спектров мощности соответственно.
РИСУНОК 2. Влияние на спектры мощности ЭЭГ (альфа, дельта или тета) после экспериментальных испытаний: имитация, наполнитель или AA-5-HT (5, 10 или 20 мг / кг; внутрибрюшинно). Никаких эффектов не наблюдалось в альфа- или тета-спектрах мощности, когда AA-5-HT вводили во время периода включения света. (A – C) соответственно). Затем, когда AA-5-HT вводили в начале периода выключения света, соединение уменьшало альфа (для W: α = 8–12 Гц) и увеличивало дельта (для SWS: Δ = 0,5–4,0 Гц). а также тета (для REMS: = 6,0–12,0 Гц; (D – F) . Среднее ± SEM * по сравнению с имитацией / наполнителем, P <0,05; & по сравнению с AA-5-HT-5 , P <0,05; # по сравнению с AA-5-HT-10, P <0,05). Сокращения: медленный сон (SWS) и сон с быстрым движением глаз (REMS), AA-5-HT-5 мг / кг (AA-5-HT-5), AA-5-HT-10 мг / кг (AA- 5HT-10), AA-5-HT-20 мг / кг (AA-5-HT-20).
Эксперимент 2: Влияние на внеклеточные уровни моноаминов или AD после введения AA-5-HT во время периода выключения света
В экспериментах по микродиализу положение зонда было подтверждено, как показано на фиг. 3A. Поскольку мы наблюдали, что AA-5-HT увеличивал сон во время активного периода крыс, в экспериментах по микродиализу соединение давали в начале периода выключения света. После инъекции AA-5-HT мы обнаружили снижение содержания DA ( F (4,20) = 7775.000, P < 0,0001), NE ( F (4,20) = 1066,582, P < 0,0001), EP ( F (4,20) = 873,101; P < 0,0001) и 5-HT ( F (4,20) = 1288,113, P < 0,0001), тогда как уровни AA-5-HT повышены AD ( F (4,20) = 773.200, P < 0,0001; рисунки 3B – F соответственно). Дозозависимый эффект наблюдался в содержании DA, NE, EP, 5-HT и AD у крыс, получавших AA-5-HT.Апостериорный тест Scheffé показал межгрупповые различия между имитацией / наполнителем и AA-5-HT (5, 10, 20 мг / кг) для DA, NE, EP, 5-HT и AD ( P < 0,0001). Мы пришли к выводу, что инъекция AA-5-HT во время периода выключения снижала дозозависимым образом содержание моноаминов, в то время как вызывала противоположный эффект в уровнях AD.
РИСУНОК 3. Влияние введения AA-5-HT (5, 10 или 20 мг / кг; внутрибрюшинно) в период выключения света на внеклеточные уровни дофамина (DA), норадреналина (NE), адреналин (EP), серотонин (5-HT) и аденозин (AD). Схематическое изображение положения микродиализного зонда в AcbC ( A – черные точки представляют собой кончик микродиализного зонда). AA-5-HT снизил содержание DA, NE, EP и 5-HT, тогда как уровни AD были увеличены ( B – F , соответственно. Среднее ± SEM * по сравнению с имитацией / наполнителем, P <0,05 ; & по сравнению с AA-5-HT-5, P <0,05; # по сравнению с AA-5-HT-10, P <0,05). Сокращения: AA-5-HT-5 мг / кг (AA-5-HT-5), AA-5-HT-10 мг / кг (AA-5-HT-10), AA-5-HT-20 мг. / Кг (AA-5-HT-20).Рисунок взят из Атласа мозга крыс Паксиноса и Ватсона (2005 г.).
Эксперимент 3: AA-5-HT блокирует побуждающие свойства каннабидиола или модафинила во время периода включения света
Затем мы проверили, блокирует ли AA-5-HT побуждающие к пробуждению эффекты CBD в период включения света. По этическим причинам животные из исследования «сон-бодрствование» (группы имитации, носителя и AA-5-HT, рис. 1) были использованы для сравнений в этом разделе эксперимента. Как описано ранее, AA-5-HT не вызывал статистических различий в пробуждении, SWS и REMS при введении во время периода включения света.Однако введение CBD (30 мг / кг, внутрибрюшинно) усиливало настороженность и уменьшало SWS, а также REMS. Этот эффект подтвердил предыдущие сообщения (Murillo-Rodríguez et al., 2006, 2008b). Интересно, что AA-5-HT, введенный за 15 минут до инъекции CBD, был способен блокировать увеличение W ( F (4,20) = 14,502, P < 0,0001), а также усиление SWS ( F (4,20) = 8,372, P < 0,0004) и REMS ( F (4,20) = 3.675, P < 0,02; Рисунки 4A – C соответственно). Апостериорный тест Scheffé показал межгрупповые различия между имитацией / носителем и AA-5-HT, а также CBD и AA-5-HT + CBD для бодрствования, SWS и REMS ( P <0,01). Мы пришли к выводу, что AA-5-HT блокирует пробуждающие эффекты CBD.
РИСУНОК 4. Влияние на общее время W, SWS и REMS после экспериментальных испытаний, введенных в начале периода включения света: имитация, носитель, AA-5-HT (20 мг / кг; i.p.), каннабидиол (CBD, 30 мг / кг; i.p.) или AA-5-HT (20 мг / кг; i.p.) и 15 мин CBD (30 мг / кг; i.p.). AA-5-HT не вызывал статистической разницы в пробуждении, SWS и REMS. Однако CBD (30 мг / кг; внутрибрюшинно) повысил настороженность и снизил SWS, а также REMS. Интересно, что AA-5-HT, введенный за 15 минут до инъекции CBD, блокировал увеличение W, а также усиление SWS и частично REMS, вызванное CBD ( A – C , соответственно; Среднее ± SEM * по сравнению с ложным / Автомобиль, P <0.05; & по сравнению с AA-5-HT-20, P <0,05; # по сравнению с CBD-30, P <0,05). Сокращения: каннабидиол 30 мг / кг, внутрибрюшинно. (CBD-30), AA-5-HT-20 мг / кг (AA-5-HT-20).
Нашей следующей целью было определить, может ли AA-5-HT предотвратить бдительность, вызванную MOD, если его ввести в начале периода включения света. По этическим причинам для сравнения в этом эксперименте были использованы крысы из исследования «сон-бодрствование» (фиктивные группы, группы носителя и AA-5-HT, рис. 1). Как описано ранее, AA-5-HT не вызывал статистических различий в бдительности, SWS и REMS при введении в начале периода включения света.Тем не менее, введение MOD (30 мг / кг, внутрибрюшинно) увеличивало W и уменьшало SWS. Статистических различий в REMS не обнаружено. Эффекты индуцирующих пробуждение свойств MOD в нашем эксперименте подтвердили предыдущие наблюдения (Scammell et al., 2000; Murillo-Rodríguez et al., 2007a, 2008c). Важно отметить, что введение AA-5-HT за 15 минут до инъекции MOD предотвращало усиление бодрствования ( F (4,20) = 9,941, P < 0,0001), а также уменьшение SWS ( F (4,20) = 9.409, P <0,0002; Рисунки 5A – C соответственно). Апостериорный тест Scheffé показал межгрупповые различия между имитацией / носителем и AA-5-HT, а также MOD и AA-5-HT + MOD для пробуждения, SWS и REMS ( P <0,0001). Наши данные предполагают, что AA-5-HT блокировал увеличение, наблюдаемое при бодрствовании, а также снижение SWS, вызванное MOD.
РИСУНОК 5. Влияние на общее время W, SWS и REMS после экспериментальных испытаний: имитация, носитель, AA-5-HT (20 мг / кг; i.p.), модафинил (MOD, 30 мг / кг; i.p.) или AA-5-HT (20 мг / кг; i.p.) и 15 мин MOD (30 мг / кг; i.p.). AA-5-HT не вызывал статистической разницы в пробуждении, SWS и REMS при введении в начале периода включения света. Однако введение MOD (30 мг / кг; внутрибрюшинно) увеличивало W и уменьшало SWS. Статистических изменений REMS у животных, получавших MOD, не наблюдалось. Интересно, что введение AA-5-HT за 15 минут до инъекции MOD предотвращало усиление пробуждения, а также уменьшение SWS.Несмотря на то, что MOD не вызывал никаких эффектов в REMS, комбинация AA-5-HT + MOD уменьшала REMS ( A – C , соответственно), индуцированную стимулятором (Среднее ± SEM * по сравнению с имитацией / носителем, P < 0,05; & по сравнению с AA-5-HT-20, P <0,05; # по сравнению с MOD-30, P <0,05). Сокращения: модафинил 30 мг / кг, внутрибрюшинно. (MOD-30), AA-5-HT-20 мг / кг (AA-5-HT-20).
В следующем эксперименте мы оценили, будет ли AA-5-HT блокировать эффекты CBD в спектрах мощности (альфа, полученная во время W, дельта, полученная во время SWS, и тета, зарегистрированная во время REMS) во время периода включения света.По этическим причинам для сравнения в этом эксперименте использовали животных из эксперимента со спектрами мощности (группы имитации, носителя и AA-5-HT, рис. 2). Как было показано ранее, введение AA-5-HT не вызывало статистических изменений в спектрах мощности при введении во время периода включения света. Несмотря на это, CBD (30 мг / кг, внутрибрюшинно) увеличивал альфа и уменьшал дельта- и тета-спектры мощности. Более того, введение AA-5-HT перед инъекцией CBD предотвращало усиление, вызванное каннабиноидом, в альфа ( F (4,20) = 5.463, P < 0,002), частично предотвращает уменьшение дельта ( F (4,20) = 5,995 , P < 0,002) и тета-спектров мощности ( F (4,20 ) = 5,112, P < 0,002; рисунки 6A – C соответственно). Дальнейший анализ внутригрупповых эффектов (тест Schefféś post hoc ) показал различия между имитацией / наполнителем и AA-5-HT, а также CBD и AA-5-HT + CBD для альфа-, дельта- и тета-спектров мощности ( P < 0.001). Мы пришли к выводу, что AA-5-HT был способен блокировать увеличение альфа-спектров мощности, тогда как ограниченный предотвращающий эффект наблюдался в дельта- и тета-спектрах мощности у крыс, получавших CBD.
РИСУНОК 6. Влияние на спектры мощности ЭЭГ (альфа [для W: α = 8–12 Гц], дельта [для SWS: Δ = 0,5–4,0 Гц] или тета [для REMS: Θ = 6,0–12,0 Гц] ]) после экспериментальных испытаний: имитация, носитель, AA-5-HT (20 мг / кг; внутрибрюшинно), каннабидиол (CBD; 30 мг / кг; внутрибрюшинно) или AA-5-HT (20 мг / кг; внутрибрюшинно) и 15 мин CBD (30 мг / кг; i.p.) в начале периода включения света. Введение AA-5-HT не вызывало статистических изменений спектров мощности. Однако CBD (30 мг / кг; i.p.) увеличивал альфа и уменьшал дельта, а также мощность тета. Кроме того, введение AA-5НТ перед инъекцией КБР предотвратить изменения, вызванные каннабиноидов в альфа и частично дельта и тета-спектры мощности спектров ( А-С , соответственно, среднее значение ± SEM * по сравнению с Sham / Транспортное средство, P <0,05; & vs.AA-5-HT-20, P <0,05; # по сравнению с CBD-30, P <0,05). Сокращения: каннабидиол 30 мг / кг, внутрибрюшинно. (CBD-30), AA-5-HT-20 мг / кг (AA-5-HT-20).
Сможет ли AA-5-HT блокировать эффекты MOD в спектрах мощности во время периода включения света, оценивали в следующем эксперименте. По этическим соображениям в этом разделе исследования для сравнений использовались крысы из эксперимента со спектрами мощности (фиктивные группы, группы носителя и AA-5-HT, рис. 2). Мы обнаружили, что AA-5-HT не вызывает статистических эффектов в спектрах мощности.Однако введение MOD (30 мг / кг, внутрибрюшинно) во время периода включения света увеличивало альфа-спектры и уменьшало дельта- и тета-спектры мощности. Более того, введение AA-5-HT перед инъекцией MOD блокировало увеличение альфа ( F (4,20) = 6,872, P < 0,01) и предотвращало уменьшение дельта ( F (4,20) = 4,715, P < 0,01) и тета-спектры мощности ( F (4,20) = 5,501, P < 0.01; Рисунки 7A – C соответственно). Статистический внутригрупповой анализ (апостериорный тест Scheffé ) показал различия между имитацией / наполнителем и AA-5-HT, а также MOD и AA-5-HT + MOD для альфа-, дельта- и тета-спектров мощности ( P <0,01 ). В заключение, мы заметили, что AA-5-HT предотвращает повышение альфа, а также уменьшение дельта- и тета-спектров мощности, вызванное инъекцией MOD.
РИСУНОК 7. Влияние на спектры мощности ЭЭГ (альфа [для W: α = 8–12 Гц], дельта [для SWS: Δ = 0.5-4,0 Гц] или тета [для REMS: = 6,0–12,0 Гц]) после экспериментальных испытаний: имитация, носитель, AA-5-HT (20 мг / кг; внутрибрюшинно), модафинил (MOD; 30 мг / кг ; ip) или AA-5-HT (20 мг / кг; ip) и 15 мин MOD (30 мг / кг; ip) в начале периода включения света. AA-5-HT не вызывал статистических эффектов в спектрах мощности. Однако введение MOD (30 мг / кг; внутрибрюшинно) увеличивало альфа-спектры и уменьшало дельта- и тета-спектры мощности. Более того, инъекция AA-5-HT перед введением MOD могла блокировать усиление альфа-, дельта- и тета-спектров мощности ( A – C , соответственно.Среднее ± SEM * по сравнению с имитацией / наполнителем, P <0,05; & по сравнению с AA-5-HT-20, P <0,05; # по сравнению с MOD-30, P <0,05). Сокращения: модафинил 30 мг / кг, внутрибрюшинно. (MOD-30), AA-5-HT-20 мг / кг (AA-5-HT-20).
Эксперимент 4: Воздействие на внеклеточные уровни моноаминов или AD, вызванное каннабидиолом или модафинилом во время периода включения света, предотвращается инъекцией AA-5-HT
В следующей серии экспериментов мы описали блокирующие эффекты AA-5-HT на внеклеточные уровни DA, NE, EP, 5-HT и AD в ответ на CBD в течение периода включения света.Введение AA-5-HT уменьшало внеклеточное содержание DA, NE, 5-HT, а также увеличивало уровни AD. Статистических различий в содержании ФП не обнаружено. Более того, инъекция CBD (30 мг / кг, внутрибрюшинно) увеличивала все изученные нейротрансмиттеры. Затем введение AA-5-HT перед инъекцией CBD частично предотвращало усиление DA, как показано двухфакторным дисперсионным анализом ANOVA ( F (4,20) = 1964,342, P <0,0001) и блокировало увеличение NE ( F (4,20) = 451.868, P <0,0001), EP ( F (4,20) = 107,976, P <0,0001), 5-HT ( F (4,20) = 790,761, P <0,0001), а также повышение уровней AD ( F (4,20) = 843,897, P <0,0001; Рисунки 8A – E, соответственно). Дальнейший апостериорный тест Scheffé показал межгрупповые различия между имитацией / носителем и AA-5-HT, а также CBD и AA-5-HT + CBD в уровнях DA, NE, EP, 5-HT, AD ( Р <0.0001). Мы пришли к выводу, что инъекция AA-5-HT перед введением CBD была способна блокировать повышение уровней нейромедиаторов в ответ на введение CBD.
РИСУНОК 8. Влияние на внеклеточные уровни DA, NE, EP, 5-HT и AD после введения AA-5-HT (20 мг / кг; внутрибрюшинно), каннабидиола (CBD; 30 мг / Кг; внутрибрюшинно) или AA-5-HT (20 мг / кг; внутрибрюшинно) и 15 мин CBD (30 мг / кг; внутрибрюшинно) в начале периода включения света. Введение AA-5-HT уменьшило внеклеточное содержание DA, NE, 5-HT, а также увеличило уровни AD.Статистических различий в содержании NE не наблюдалось. Более того, инъекция CBD (30 мг / кг; внутрибрюшинно) увеличивала уровни всех нейротрансмиттеров. Затем введение AA-5-HT перед инъекцией CBD могло предотвратить повышение уровней DA, NE, EP, 5-HT, а также AD ( A – E , соответственно. Среднее ± SE. ∗ против имитации / наполнителя, P <0,05; & против AA-5-HT-20, P <0,05; # против CBD-30, P <0,05). Сокращения: каннабидиол 30 мг / кг, т.е.п. (CBD-30), AA-5-HT-20 мг / кг (AA-5-HT-20).
Чтобы дополнительно охарактеризовать, будет ли AA-5-HT, введенная до MOD, предотвращать эффекты этого стимулятора на внеклеточные уровни DA, NE, EP, 5-HT и AD в течение периода включения света, мы разработали соответствующий эксперимент. Принимая во внимание этические причины, животные из исследования микродиализа (фиктивные группы, группы носителя и AA-5-HT, фиг. 8) были использованы для сравнения в этом эксперименте. Было обнаружено, что AA-5-HT снижало внеклеточное содержание DA, NE, 5-HT, тогда как повышало уровни AD.Статистических различий во внеклеточном содержимом ВП не обнаружено. С другой стороны, MOD вызывал противоположные эффекты, так как увеличивал внеклеточные уровни DA, NE, EP, 5-HT при снижении содержания AD. Важно отметить, что введение AA-5-HT перед инъекцией MOD частично предотвращало повышение уровней DA, как было проанализировано с помощью двухфакторного дисперсионного анализа ( F (4,20) = 1798,342, P < 0,0001), NE ( F (4,20) = 237,909, P < 0.0001), EP ( F (4,20) = 208,533, P <0,0001), 5-HT ( F (4,20) = 1198,470, P < 0,0001) и заблокирован снижение уровней AD ( F (4,20) = 1614,168, P < 0,0001; Рисунки 9A – E, соответственно). Дальнейший апостериорный тест Шеффе показал межгрупповые различия между имитацией / носителем и AA-5-HT, а также MOD и AA-5-HT + MOD для DA, NE, EP, 5-HT и AD ( P <0.0001). Основываясь на наших выводах, можно сделать вывод, что AA-5-HT предотвращает эффекты нейротрансмиттеров, вызванные инъекцией CBD в период включения света.
РИСУНОК 9. Влияние на внеклеточные уровни DA, NE, EP, 5-HT и AD после введения AA-5-HT (20 мг / кг; внутрибрюшинно), модафинила (MOD; 30 мг / Кг; внутрибрюшинно) или AA-5-HT (20 мг / кг; внутрибрюшинно) и 15 мин MOD (30 мг / кг; внутрибрюшинно) в начале периода включения света. AA-5-HT уменьшало внеклеточное содержание катехоламинов, тогда как повышало уровни AD.С другой стороны, MOD вызвал противоположные эффекты. Важно отметить, что введение AA-5-HT перед инъекцией MOD частично предотвращало повышение уровней DA, NE, EP, 5-HT и блокировало снижение содержания AD ( A – E соответственно. Среднее ± SEM * по сравнению с имитацией / наполнителем, P <0,05; & по сравнению с AA-5-HT-20, P <0,05; # по сравнению с MOD-30, P <0,05). Сокращения: модафинил 30 мг / кг, внутрибрюшинно. (MOD-30), AA-5-HT-20 мг / кг (AA-5-HT-20).
Эксперимент 5: блокада восстановления сна, вызванная каннабидиолом или модафинилом, после полной депривации сна предотвращается AA-5-HT
Как было показано ранее в текущем отчете, AA-5-HT способствовал сну в течение активного периода у крыс и уменьшал количество нейромедиаторов, связанных с бодрствованием. Более того, этот препарат блокировал пробуждающие эффекты CBD или MOD. Однако, чтобы охарактеризовать свойства AA-5-HT в гомеостазе сна, мы исследовали возможность того, что блокада CBD или MOD компенсаторного восстановления сна после TSD может быть предотвращена путем инъекции AA-5-HT во время включения света. период.Мы обнаружили, что AA-5-HT не вызывал статистических изменений в W, SWS или REMS, если вводился после TSD. Более того, во время периода восстановления сна CBD или MOD увеличивают пробуждение и уменьшают SWS и REMS. Интересно, что во время периода восстановления сна введение AA-5-HT перед инъекцией CBD частично предотвращало увеличение W, как показано с помощью двухфакторного дисперсионного анализа ( F, , (6,28) = 6063,477; P < 0,0001), а также снижение SWS ( F (6,28) = 3649.311; P < 0,0001) и REMS ( F (6,28) = 1050,707; P < 0,0001; рисунки 10A – C соответственно). Аналогичные результаты были получены, когда AA-5-HT вводили перед MOD во время периода восстановления сна. Дальнейший апостериорный тест Scheffé показал межгрупповые различия между имитацией / носителем и AA-5-HT, а также CBD, MOD, AA-5-HT + CB, а также AA-5-HT + MOD для бдительности, SWS. и REMS ( P <0,0001). Мы пришли к выводу, что AA-5-HT был частично способен предотвратить повышение W, а также снижение SWS или REMS, вызванное либо CDB, либо MOD во время периода восстановления сна после TSD.
РИСУНОК 10. Эффекты экспериментальных испытаний: имитация, носитель, AA-5-HT (20 мг / кг; внутрибрюшинно), каннабидиол (CBD; 30 мг / кг; ip), AA-5-HT (20 мг / Кг; внутрибрюшинно) и через 15 минут CBD (30 мг / кг; ip), модафинил (MOD; 30 мг / кг; внутрибрюшинно) или AA-5-HT (20 мг / кг; внутрибрюшинно) и через 15 минут MOD ( 30 мг / кг; ip) на W, SWS или REMS во время периода восстановления сна у лишенных сна крыс. AA-5-HT не вызывал статистических изменений в W, SWS или REMS после полного лишения сна. Более того, во время периода восстановления сна CBD или MOD увеличивают пробуждение и уменьшают SWS и REMS.Интересно, что во время периода восстановления сна введение AA-5-HT перед инъекцией CBD предотвращало увеличение W, а также снижение SWS и частично REMS у крыс, получавших CBD. Аналогичные результаты в отношении сна были получены, когда AA-5-HT вводили перед MOD после полного лишения сна ( A – C , соответственно. Среднее ± SEM ∗ по сравнению с имитацией / наполнителем, P <0,05; & vs. AA-5HT-20, P <0,05; # по сравнению с CBD-30, P <0.05; % по сравнению с MOD-30, P <0,05). Сокращения: каннабидиол 30 мг / кг, внутрибрюшинно. (CBD-30), модафинил 30 мг / кг, внутрибрюшинно (MOD-30), AA-5-HT-20 мг / кг (AA-5-HT-20).
Затем мы оценили изменения спектров мощности после TSD у животных, которые получали AA-5-HT, CBD, MOD или введение AA-5-HT перед CBD или MOD. Мы обнаружили, что AA-5-HT не влиял на спектры мощности. Однако CBD или MOD увеличивали альфа-спектры и уменьшали дельта- и тета-спектры мощности. Наиболее заметный эффект наблюдался у животных, получавших CBD.Важно отметить, что введение AA-5-HT перед инъекцией CBD или MOD частично уменьшало увеличение альфа, как было проверено с помощью двухфакторного дисперсионного анализа ( F (6,28) = 4,474, P < 0,004) , заблокировал уменьшение дельта ( F (6,28) = 4,079, P < 0,004) и предотвратил уменьшение спектров тета-мощности ( F (6,28) = 5,109, P < 0,004; Рисунки 11A – C, соответственно) в период восстановления сна.Кроме того, межгрупповые различия были обнаружены с помощью апостериорного теста Шеффе для альфа-, дельта- и тета-спектров мощности ( P <0,0001). Мы пришли к выводу, что AA-5-HT был частично способен предотвратить влияние на спектры мощности после инъекции CBD или MOD после TSD.
РИСУНОК 11. Эффекты экспериментальных испытаний: имитация, носитель, AA-5-HT (20 мг / кг; внутрибрюшинно), каннабидиол (CBD; 30 мг / кг; ip), AA-5-HT (20 мг / Кг; внутрибрюшинно) и через 15 минут CBD (30 мг / кг; i.p.), модафинил (MOD; 30 мг / кг; внутрибрюшинно) или AA-5-HT (20 мг / кг; внутрибрюшинно) и через 15 минут MOD (30 мг / кг; внутрибрюшинно) во время периода восстановления сна во сне. лишенные крысы по спектрам мощности альфа (для W: α = 8–12 Гц), дельта (для SWS: Δ = 0,5–4,0 Гц) или тета-спектрам мощности (для REMS: Θ = 6,0–12,0 Гц). AA-5-HT не оказал влияния на спектры мощности. Однако CBD или MOD частично увеличили альфа и уменьшили дельта- и тета-спектры мощности. Важно отметить, что введение AA-5-HT перед инъекцией CBD или MOD предотвращало повышение альфа, а также блокировало уменьшение дельта и тета ( A – C , соответственно.Среднее ± SEM * по сравнению с имитацией / наполнителем, P <0,05; & по сравнению с AA-5-HT-20, P <0,05; # по сравнению с CBD-30, P <0,05; % по сравнению с MOD-30, P <0,05). Сокращения: каннабидиол 30 мг / кг, внутрибрюшинно. (CBD-30), модафинил 30 мг / кг, внутрибрюшинно (MOD-30), AA-5-HT-20 мг / кг (AA-5-HT-20).
Эксперимент 6. Блокада компенсирующего восстановления внеклеточных уровней моноаминов или AD после полной депривации сна каннабидиолом или модафинилом предотвращается AA-5-HT в течение периода включения света
В последнем эксперименте мы исследовали, можно ли предотвратить блокаду компенсаторного баланса внеклеточных уровней DA, NE, EP, 5-HT или AD после TSD посредством CBD или MOD с помощью AA-5-HT.Мы обнаружили, что AA-5-HT снижает после TSD внеклеточные уровни изученных нейромедиаторов. Кроме того, инъекция CBD или MOD увеличивала содержание DA, как определено с помощью двухфакторного дисперсионного анализа ( F (6,28) = 1952,414, P < 0,0001), NE ( F (6, 28) = 436,222, P < 0,0001), EP ( F (6,28) = 212,732, P < 0,0001), 5-HT ( F (6,28) = 1384.522; P < 0.0001) и AD ( F (6,28) = 1743,304 , P < 0,0001; рисунки 12A – E, соответственно) в период восстановления сна. Наиболее заметный эффект наблюдался у крыс, которым вводили CBD. Более того, межгрупповые различия были обнаружены Шеффе post hoc между имитацией / носителем и AA-5-HT, CBD, MOD, AA-5-HT + CB, а также AA-5-HT + MOD для DA, NE. , EP, 5-HT и AD ( P <0,0001). Комбинация AA-5-HT + CBD частично блокировала повышение уровней нейромедиаторов у животных, получавших CBD.Подобные результаты наблюдались у крыс, получавших MOD. Мы пришли к выводу, что AA-5-HT не полностью блокировал усиление нейротрансмиттеров у животных, которые получали CBD или MOD во время периода восстановления сна после TSD.
РИСУНОК 12. Эффекты экспериментальных испытаний: имитация, носитель, AA-5-HT (20 мг / кг; внутрибрюшинно), каннабидиол (CBD; 30 мг / кг; ip), AA-5-HT (20 мг / Кг; внутрибрюшинно) и через 15 минут CBD (30 мг / кг; внутрибрюшинно), модафинил (MOD; 30 мг / кг; внутрибрюшинно) или AA-5-HT (20 мг / кг; i.p.) и 15 минут спустя MOD (30 мг / кг; внутрибрюшинно) во время периода восстановления сна у лишенных сна крыс на внеклеточных уровнях DA, NE, EP, 5-HT и AD. AA-5-HT снижает внеклеточные уровни всех изученных нейромедиаторов. Более того, инъекция CBD или MOD увеличивала содержание DA, NE, EP, 5-HT и AD ( A – E соответственно) во время периода восстановления во сне. Важно отметить, что комбинация AA-5-HT + CBD частично предотвращала вызванные CBD изменения в нейромедиаторах. Сходные результаты наблюдались у крыс, получавших AA-5-HT + MOD, по сравнению с животными, которым вводили MOD (Среднее ± SEM * vs.Имитация / наполнитель, P <0,05; & по сравнению с AA-5-HT-20, P <0,05; # по сравнению с CBD-30, P <0,05; % по сравнению с MOD-30, P <0,05). Сокращения: каннабидиол 30 мг / кг, внутрибрюшинно. (CBD-30), модафинил 30 мг / кг, внутрибрюшинно (MOD-30), AA-5-HT-20 мг / кг (AA-5-HT-20).
Обсуждение
Эндоканнабиноидная система выполняет множество сложных модулирующих физиологических функций (Kendall and Yudowski, 2016; Ligresti et al., 2016; Argueta and DiPatrizio, 2017; Bennett et al., 2017; Дос Аньос-Гарсия и др., 2017; Sun et al., 2017). Например, совокупные данные свидетельствуют о том, что элементы эндоканнабиноидной системы, включая FAAH, контролируют цикл сна-бодрствования (Santucci et al., 1996; Murillo-Rodríguez et al., 1998, 2001, 2003, 2008a, 2011a, 2013). , 2016; Herrera-Solis et al., 2010; Rueda-Orozco et al., 2010; Pava et al., 2014, 2016). В связи с этим экспериментальный подход к изучению роли FAAH в модуляции сна заключался в характеристике свойств ингибиторов FAAH, таких как URB597 (Murillo-Rodríguez et al., 2007b, 2011a, 2016). Среди ингибиторов FAAH несколько соединений, таких как 1-гетероарилпропан-2-оны, пиперидинилтиазолизоксазолин, PF-04457845, JNJ-42165279, BIA 10-2474, AM3506, URB694, ARN146333-карбоксамидо-5-арил-изоксазол изоксазол -арил 2-арилоксиацетамиды обладают значительной активностью. Стоит упомянуть, что URB597 был описан как один из самых мощных ингибиторов FAAH до сих пор (Carnevali et al., 2015; Keith et al., 2015; Lodola et al., 2015; Doenni et al., 2016; Panlilio). и другие., 2016; Пембер и др., 2016; Тонг и др., 2016; Tuo et al., 2016; Захов и др., 2017; Сундуру и др., 2017; Wilkerson et al., 2017). Однако полное описание этих ингибиторов FAAH для контроля сна требует внимания.
AA-5-HT – двойной блокатор FAAH / TRPV1, который проявляет множественные фармакологические эффекты. Например, это соединение оказывает анксиолитическое действие на мышей, испытанных в открытых руках (Micale et al., 2009; John and Currie, 2012). Более того, AA-5-HT также снижает депрессивно-подобное поведение без изменения двигательной активности (Navarria et al., 2014; Сартим и др., 2017). Стоит отметить, что, насколько нам известно, в текущем отчете впервые представлены прямые доказательства влияния AA-5-HT на модуляцию сна. В связи с этим системное введение этого препарата увеличивало сон во время выключенного света, но не в период включения света. Более того, мы обнаружили, что AA-5-HT увеличивали спектры мощности, связанные со сном, такие как дельта и тета, тогда как количество нейротрансмиттеров, связанных с бодрствованием, снижалось в период выключения света.Кроме того, в наших экспериментах AA-5-HT предотвращал блокировку эффектов CBD или MOD на восстановление сна, а также изменения нейротрансмиттеров после TSD. Взятые вместе, эти данные предполагают, что либо FAAH, либо TRPV1, либо оба могут модулировать сон, гомеостаз сна и содержание нейротрансмиттеров в присутствии соединений, способствующих бодрствованию, таких как CBD или MOD. Хотя в текущем отчете механистический подход не рассматривался, мы хотели бы изобразить следующий гипотетический сценарий (Рисунок 13): AA-5-HT ингибирует FAAH, что приводит к увеличению содержания олеоилэтаноламида (OEA), пальмитоилэтаноламида (PEA) в виде а также AEA (de Lago et al., 2005; Палаццо и др., 2010). В то время как OEA связывается с PPARα в ядре и способствует пробуждению (Mijangos-Moreno et al., 2016), AEA может диффундировать через клеточную мембрану с помощью AMT, связываясь либо с каннабиноидными рецепторами CB 1 , либо с TRPV1 (Leung et al., 2013; Николусси, Герч, 2015). Хотя активация каннабиноидного рецептора CB 1 была связана с генерацией сна (Murillo-Rodríguez et al., 2001, 2003, 2016), необходимы дополнительные доказательства, чтобы полностью определить, модулирует ли активация TRPV1 сон.Как упоминалось ранее, ингибирование FAAH может также приводить к увеличению уровней других субстратов FAAH. Действительно, Wollank et al. (2015) продемонстрировали более высокие уровни AEA, 2-AG, OEA и PEA в клетках, инкубированных с AA-5-HT. Эти данные были подтверждены анализом ЖХ-МС в исследованиях in vitro , показывающих повышение уровней AEA-подобных липидов, включая OEA и PEA, с URB597 (Winkler et al., 2016). Если ингибирование FAAH способствует пробуждению (Murillo-Rodríguez et al., 2007b, 2011a), как мы можем согласовать текущие результаты с имеющимися доказательствами в литературе? Поскольку AEA модулирует множество нейробиологических функций, также действуя через TRPV1 (Di Marzo and De Petrocellis, 2012; Starowicz et al., 2012), то гипотетическим объяснением было бы то, что AA-5-HT увеличивает сон за счет вовлечения TRPV1, активируемого AEA. Однако капсаицин, агонист TRPV1, увеличивает частоту возбуждения DA нейронов крысы (Marinelli et al., 2005; Chakraborty et al., 2016). Более того, AA-5-HT способствует электрофизиологической активности нейронов голубого пятна (de Novellis et al., 2008). Широко признано, что обе области мозга связаны с бодрствованием, поскольку частота их активности выше во время бодрствования, а не во время сна (Berridge et al., 2012). Таким образом, расхождения между текущими результатами и доступными доказательствами могут включать – помимо методологических различий между сопоставимыми исследованиями – более сложный механизм действия. Можно предположить, что AA-5-HT индуцирует сон не только путем облегчения эндогенного действия AEA на каннабиноидные рецепторы CB1, но также путем противодействия его активации TRPV1. Таким образом, наши данные заставляют нас предположить, что AA-5-HT действует как двойной ингибитор FAAH и антагонист TRPV1, но это не обязательно следует рассматривать как предсказуемое, потому что: (1) активность AA-5-HT проявляется при более высокой концентрации по сравнению с другими более мощными ингибиторами FAAH, такими как URB597, и (2) роль AEA как полного или частичного агониста TRPV1 в улучшении сна полностью не изучена.Тем не менее, AA-5-HT может демонстрировать более высокую предпочтительность связывания с TRPV1, что приводит к увеличению уровней AEA, что, в свою очередь, приведет к стимулированию сна за счет активации каннабиноидных рецепторов CB 1 , а не к ингибированию сна через TRPV1. Исследования, посвященные вытеснению AA-5-HT из сайтов связывания TRPV1 одновременно с AEA, дадут новое понимание функции эндоканнабиноидной системы.
РИСУНОК 13. Гипотетический молекулярный механизм действия AA-5-HT на стимуляцию сна. В этом эмпирическом предложении AA-5-HT ингибирует амидгидролазу жирных кислот (FAAH), которая, в свою очередь, увеличивает уровни олеоилэтаноламида (OEA) и пальмитоилэтаноламида (PEA), а также анандамида (AEA). В то время как OEA связывается с PPARα в ядре и способствует пробуждению, AEA способна диффундировать через клеточную мембрану с помощью мембранного переносчика анандамида (AMT), связываясь либо с каннабиноидным рецептором CB 1 , либо с TRPV1. AA-5-HT может также противодействовать действию AEA на TRPV1. Хотя активация каннабиноидного рецептора CB 1 была связана с генерацией сна, необходимы дополнительные доказательства, чтобы определить, будет ли активация или ингибирование TRPV1 модулировать сон.
Наконец, хотя сравнительные исследования различных экспериментальных подходов относительно того, что могло бы быть наиболее надежной стратегией манипулирования эндоканнабиноидной системой (эндогенные лиганды, рецепторы, переносчики, синтезирующие / разлагающие ферменты и т. Д.) При модуляции сна, не проводились, В текущем отчете представлены дополнительные доказательства роли AA-5-HT в контроле сна. На данный момент нейромолекулярная роль эндоканнабиноидной системы в регуляции цикла сна и бодрствования дала важную информацию об участии AEA, каннабиноидного рецептора CB 1 , AMT, а также FAAH (Murillo-Rodríguez et al., 1998, 2001, 2003, 2007b, 2008a, 2011a, 2016; Эррера-Солис и др., 2010; Пава и др., 2014, 2016; Руэда-Ороско и др., 2010). Однако идентификация новых соединений, нацеленных на эндокананбиноидную систему, поможет подтвердить роль этой системы в контроле сна, выделив наиболее подходящее лекарство с наивысшей эффективностью и наименьшими побочными эффектами на животных моделях (Lauria et al., 2015; Nimczick and Decker, 2015). ; Bertini et al., 2016; Aizpurua-Olaizola et al., 2017; Chicca et al., 2017).
Действительно, необходимы исследования, чтобы полностью описать механизм действия, с помощью которого AA-5-HT способствует сну и модулирует активность спектров мощности.Более того, регуляция ферментов, ответственных за синтез и / или деградацию нейротрансмиттеров под влиянием AA-5-HT, также неизвестна. Кроме того, роль AA-5-HT как блокатора настороженности, вызванного CBD или MOD в период восстановления сна после TSD, все еще требует дальнейших исследований.
Заключение
Таким образом, AA-5-HT способствует сну и снижает количество нейротрансмиттеров, связанных с бодрствованием, в период выключения света и модулирует гомеостаз сна в присутствии соединений, способствующих бодрствованию, таких как CBD или MOD.Полное понимание механизмов, посредством которых AA-5-HT модулирует сон, требует дальнейших исследований.
Авторские взносы
EM-R разработал, провел эксперименты, проанализировал данные и написал рукопись. В.Д. оказал методическую поддержку и способствовал написанию рукописи. SM пересмотрел экспериментальный план. NBR, ABV, GAMN, HB, OA-C, пересмотренные проанализированные данные. GA-S оказала методическую поддержку. Все авторы отредактировали всю статью и одобрили окончательную версию рукописи.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарность
Эта работа была поддержана финансированием (предоставленным EM-R; номер гранта 03-1213) от Escuela de Medicina, Universidad Anáhuac Mayab, Мерида, Юкатан, Мексика.
Список литературы
Айзпуруа-Олайзола, О., Элезгарай, И., Рико-Баррио, И., Зарандона, И., Эчебаррия, Н., и Усобиага, А. (2017). Ориентация на эндоканнабиноидную систему: будущие терапевтические стратегии. Drug Discov. Сегодня 1, 105–110. DOI: 10.1016 / j.drudis.2016.08.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Аргуэта, Д. А., и Ди Патрицио, Н. В. (2017). Передача периферических эндоканнабиноидов контролирует гиперфагию при ожирении, вызванном западной диетой. Physiol. Behav. 171, 32–39. DOI: 10.1016 / j.физбех.2016.12.044
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беннетт М. Р., Арнольд Дж., Хаттон С. Н. и Лагопулос Дж. (2017). Регулирование угасания страха длительной депрессией: роль эндоканнабиноидов и нейротрофического фактора мозга. Behav. Brain Res. 319, 148–164. DOI: 10.1016 / j.bbr.2016.11.029
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бертини, С., Чикка, А., Арена, К., Чикка, С., Saccomanni, G., Gertsch, J., et al. (2016). Синтез и фармакологическая оценка новых бифенильных производных как лигандов рецептора CB2. Eur. J. Med. Chem. 116, 252–266. DOI: 10.1016 / j.ejmech.2016.03.072
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Biernacki, M., and Skrzydlewska, E. (2016). Метаболизм эндоканнабиноидов. Постэпы Выс. Med. Dosw. 70, 830–843. DOI: 10.5604 / 17322693.1213898
CrossRef Полный текст | Google Scholar
Бланко-Центурион, К., Сюй, М., Мурильо-Родригес, Э., Геращенко, Д., Широмани, А. М., Салин-Паскуаль, Р. Дж. И др. (2006). Аденозин и гомеостаз сна в базальном переднем мозге. J. Neurosci. 26, 8092–8100. DOI: 10.1523 / JNEUROSCI.2181-06.2006
CrossRef Полный текст | Google Scholar
Carnevali, L., Vacondio, F., Rossi, S., Macchi, E., Spadoni, G., Bedini, A., et al. (2015). Кардиозащитные эффекты ингибитора амидгидролазы жирных кислот URB694 на модели тревожности на грызунах. Sci. Реп. 5: 18218. DOI: 10.1038 / srep18218
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чакраборти, С., Ребекки, М., Качоча, М., и Пуополо, М. (2016). Дофаминовая модуляция временного рецепторного потенциала ваниллоидного рецептора типа 1 (TRPV1) в нейронах ганглиев задних корешков. J. Physiol. 594, 1627–1642. DOI: 10.1113 / JP271198
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чен, Д. Дж., Гао, М., Гао, Ф.Ф., Су, К. X., Ву, Дж. (2017). Каннабиноидный рецептор 2 мозга: экспрессия, функция и модуляция. Acta Pharmacol. Грех. 38, 312–316. DOI: 10.1038 / aps.2016.149
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Chen, J., Varga, A., Selvarajah, S., Jenes, A., Dienes, B., Sousa-Valente, J., et al. (2016). Пространственное распределение рецепторов каннабиноидов типа 1 и капсаицина может вносить вклад в сложность их перекрестных помех. Sci. Реп. 6: 33307.DOI: 10.1038 / srep33307
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Chicca, A., Berg, R., Jessen, H.J., Marck, N., Schmid, F., Burch, P., et al. (2017). Биологическая оценка пиридоновых алкалоидов на эндоканнабиноидную систему. Bioorg. Med. Chem. 16, 31251–31252. DOI: 10.1016 / j.bmc.2017.02.031
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кристенсен, Дж. А., Мунк, Э. Г., Пеппард, П. Э., Янг, Т., Миньо, Э., Соренсен, Х. Б. и др. (2015). Диагностическая ценность анализа спектров мощности электроэнцефалографии сна у больных нарколепсией. Sleep Med. 16, 1516–1527. DOI: 10.1016 / j.sleep.2015.09.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Корреа, Ф., Вольфсон, М. Л., Валчи, П., Айсемберг, Дж., И Франчи, А. М. (2016). Эндоканнабиноидная система и беременность. Репродукция 152, R191 – R200. DOI: 10.1530 / rep-16-0167
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Корси-Кабрера, М., Перес-Гарси, Э., Дель Рио-Портилья, Ю., Угальде, Э., и Гевара, М. А. (2001). Полосы ЭЭГ во время бодрствования, медленного сна и парадоксального сна в результате анализа главных компонентов у крысы. Сон 24, 374–380. DOI: 10.1093 / сон / 24.4.374
PubMed Аннотация | CrossRef Полный текст | Google Scholar
de Lago, E., Petrosino, S., Valenti, M., Morera, E., Ortega-Gutierrez, S., Fernandez-Ruiz, J., et al. (2005). Влияние повторного системного введения селективных ингибиторов инактивации эндоканнабиноидов на уровни эндоканнабиноидов в головном мозге крыс. Biochem. Pharmacol. 70, 446–452. DOI: 10.1016 / j.bcp.2005.05.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
de Novellis, V., Palazzo, E., Rossi, F., De Petrocellis, L., Petrosino, S., Guida, F., et al. (2008). Обезболивающее действие N-арахидоноил-серотонина, ингибитора FAAH и антагониста рецептора TRPV1, связано с изменениями активности ростральных вентромедиальных клеток мозгового вещества и голубого пятна у крыс. Нейрофармакология 55,1105–1113.DOI: 10.1016 / j.neuropharm.2008.06.023
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ди Марцо, В., и Де Петрочеллис, Л. (2012). Почему у каннабиноидных рецепторов более одного эндогенного лиганда? Philos. Пер. R. Soc. Лондон. B Biol. Sci. 367, 3216–3228. DOI: 10.1098 / rstb.2011.0382
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Доэнни В. М., Грей, Дж. М., Сонг, К. М., Патель, С., Хилл, М. Н., и Питтман, К. Дж.(2016). Нарушение социального поведения подростков после воспаления в раннем возрасте улучшается за счет усиления передачи сигналов анандамида. Brain Behav. Иммун. 58, 237–247. DOI: 10.1016 / j.bbi.2016.07.152
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дос Аньос-Гарсия, Т., Уллах, Ф., Фалькони-Собриньо, Л. Л., и Коимбра, Н. К. (2017). Передача сигналов анандамида, опосредованная каннабиноидным рецептором CB1, снижает защитное поведение, вызванное блокадой рецептора GABAA в дорсомедиальном отделе вентромедиального гипоталамуса. Нейрофармакология 113, 156–166. DOI: 10.1016 / j.neuropharm.2016.04.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фицпатрик, Дж. К., и Даунер, Э. Дж. (2017). Передача сигналов Toll-подобного рецептора в качестве каннабиноидной мишени при рассеянном склерозе. Нейрофармакология 113, 618–626. DOI: 10.1016 / j.neuropharm.2016.04.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Франкен П., Дейк Д. Дж., Тоблер И. и Борбели А.А. (1991). Депривация сна у крыс: влияние на спектры мощности ЭЭГ, состояние бдительности и корковую температуру. Am. J. Physiol. 261, R198 – R208.
Google Scholar
Голд П. Э., Земляк Р. А., Дукала Д. и Чанг К. (2011). Высвобождение ацетилхолина в гиппокампе и прелимбической коре головного мозга во время приобретения социально передаваемых пищевых предпочтений. Neurobiol. Учить. Mem. 96, 498–503. DOI: 10.1016 / j.nlm.2011.08.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Grupe, М., Паолоне, Г., Дженсен, А. А., Сандагер-Нильсен, К., Сартер, М., и Граннет, М. (2013). Селективное усиление никотиновых ацетилхолиновых рецепторов (альфа4) 3 (бета2) 2 увеличивает амплитуду префронтальных ацетилхолиновых и никотин-вызванных глутаматергических переходных процессов у крыс. Biochem. Pharmacol. 86, 1487–1496. DOI: 10.1016 / j.bcp.2013.09.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эррера-Солис, А., Васкес, К. Г., и Просперо-Гарсия, О. (2010). Острое и субхроническое введение анандамида или олеамида увеличивает фазу быстрого сна у крыс. Pharmacol. Biochem. Behav. 95, 106–112. DOI: 10.1016 / j.pbb.2009.12.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хуанг, В. Дж., Чен, В. В., и Чжан, X. (2016). Эндоканнабиноидная система: роль в депрессии, вознаграждении и контроле над болью (Обзор). Мол. Med. Rep. 14, 2899–2903. DOI: 10.3892 / mmr.2016.5585
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Imperatori, C., Della Marca, G., Brunetti, R., Carbone, G.А., Массулло, К., Валенти, Э. М. и др. (2016). Изменения сети в режиме по умолчанию при алекситимии: спектры мощности ЭЭГ и исследование связности. Sci Rep. 6: 36653. DOI: 10.1038 / srep36653
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Янссен, Ф. Дж., И ван дер Стелт, М. (2016). Ингибиторы диацилглицеринлипаз при нейродегенеративных и метаболических нарушениях. Bioorg. Med. Chem. Lett. 26, 3831–3837. DOI: 10.1016 / j.bmcl.2016.06.076
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Jing, W., Wang, Y., Fang, G., Chen, M., Xue, M., Guo, D., et al. (2016). Полосы ЭЭГ бодрствования, медленного сна и сна с быстрым движением глаз в различных областях мозга крыс. Перед. Comput. Neurosci. 10:79. DOI: 10.3389 / fncom.2016.00079
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джон С. С. и Карри П. Дж. (2012). N-арахидоноил-серотонин в базолатеральной миндалине усиливает анксиолитическое поведение в приподнятом крестообразном лабиринте. Behav. Brain Res. 233, 382–388.DOI: 10.1016 / j.bbr.2012.05.025
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кейт, Дж. М., Джонс, В. М., Тиченор, М., Лю, Дж., Зейерстад, М., Палмер, Дж. А. и др. (2015). Доклиническая характеристика ингибитора FAAH JNJ-42165279. ACS Med. Chem. Lett. 6, 1204–1208. DOI: 10.1021 / acsmedchemlett.5b00353
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кендалл, Д. А., Юдовски, Г. А. (2016). Рецепторы каннабиноидов в центральной нервной системе: их сигналы и роль в заболевании. Перед. Cell Neurosci. 10: 294. DOI: 10.3389 / fncel.2016.00294
CrossRef Полный текст | Google Scholar
Киркедал, К., Вегенер, Г., Морейра, Ф., Йока, С. Р., и Либенберг, Н. (2016). Двойной ингибитор каналов FAAH и TRPV1 демонстрирует дозозависимый эффект на депрессивное поведение у крыс. Acta Neuropsychiatr. 12, 1–6. DOI: 10.1017 / neu.2016.68
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Крук-Сломка, М., Дзик, А., Будзинская, Б., Бяла, Г. (2016). Эндоканнабиноидная система: прямое и косвенное участие в процессах памяти и обучения – краткий обзор. Мол. Neurobiol. DOI: 10.1007 / s12035-016-0313-5 [Epub перед печатью].
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лаурия, С., Касати, С., Чиуффреда, П. (2015). Синтез и характеристика нового флуорогенного субстрата для моноацилглицерин липазы и применение в исследованиях ингибирования. Анал.Биоанал. Chem. 407, 8163–8167. DOI: 10.1007 / s00216-015-8991-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лазарус М., Чен Дж. Ф., Ураде Ю. и Хуанг З. Л. (2013). Роль базальных ганглиев в контроле сна и бодрствования. Curr. Opin. Neurobiol. 23, 780–785. DOI: 10.1016 / j.conb.2013.02.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Lazarus, M., Huang, Z. L., Lu, J., Urade, Y., and Chen, J.Ф. (2012). Как базальные ганглии регулируют поведение сна и бодрствования? Trends Neurosci. 35, 723–732. DOI: 10.1016 / j.tins.2012.07.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Леунг К., Элмс М. В., Глейзер С. Т., Дойч Д. Г. и Качоха М. (2013). Роль FAAH-подобного переносчика анандамида в инактивации анандамида. PLoS ONE 8: e79355. DOI: 10.1371 / journal.pone.0079355
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лигрести, А., Де Петрочеллис, Л., и Ди Марцо, В. (2016). От фитоканнабиноидов до каннабиноидных рецепторов и эндоканнабиноидов: плейотропные физиологические и патологические роли через сложную фармакологию. Physiol. Ред. 96, 1593–1659. DOI: 10.1152 / Physrev.00002.2016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лю, З., Ван, Ю., Цай, Л., Ли, Ю., Чен, Б., Дун, Ю., и др. (2016). Префронтальная кора к прилежащим проекциям в регуляции вознаграждения во сне. Дж.Neurosci. 36, 7897–7910. DOI: 10.1523 / JNEUROSCI.0347-16.2016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лодола, А., Кастелли, Р., Мор, М., и Ривара, С. (2015). Ингибиторы амидгидролазы жирных кислот: патентный обзор (2009-2014). Мнение эксперта. Ther. Пат. 25, 1247–1266. DOI: 10.1517 / 13543776.2015.1067683
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лоуин, Т., и Штрауб, Р. Х. (2015). Препараты на основе каннабиноидов, нацеленные на CB1 и TRPV1, симпатическую нервную систему и артрит. Arthritis Res. Ther. 17, 226. DOI: 10.1186 / s13075-015-0743-x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Маллипедди С., Джанеро Д. Р., Звонок Н. и Макрияннис А. (2017). Функциональная селективность в отношении рецепторов, связанных с G-белком: продвижение каннабиноидных рецепторов в качестве мишеней для лекарств. Biochem. Pharmacol. 128, 1–11. DOI: 10.1016 / j.bcp.2016.11.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Маринелли, С., Паскуччи, Т., Бернарди, Г., Пуглиси-Аллегра, С., и Меркури, Н. Б. (2005). Активация TRPV1 в VTA возбуждает дофаминергические нейроны и увеличивает вызванное химическими и вредными веществами высвобождение дофамина в прилежащем ядре. Нейропсихофармакология 30, 864–870. DOI: 10.1038 / sj.npp.1300615
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Mazzucchelli, I., Rapetti, M., Fattore, C., Franco, V., Gatti, G., and Perucca, E. (2011). Разработка и валидация анализа HPLC-UV для определения руфинамида в плазме и слюне человека. Анал. Биоанал. Chem. 401, 1013–1021. DOI: 10.1007 / s00216-011-5126-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Micale, V., Cristino, L., Tamburella, A., Petrosino, S., Leggio, G.M., Drago, F., et al. (2009). Анксиолитические эффекты у мышей двойного блокатора амидгидролазы жирных кислот и временного рецепторного потенциала ваниллоидных каналов типа 1. Нейропсихофармакология 34, 593–606. DOI: 10.1038 / npp.2008.98
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Миджангос-Морено, С., Поот-Аке, А., Аранковски-Сандовал, Г., и Мурильо-Родригес, Э. (2014). Внутригипоталамическая инъекция каннабидиола увеличивает внеклеточные уровни аденозина в прилежащем ядре у крыс. Neurosci. Res. 84, 60–63. DOI: 10.1016 / j.neures.2014.04.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Миджангос-Морено, С., Поот-Аке, А., Гусман, К., Аранковски-Сандовал, Г., Ариас-Каррион, О., Залдивар-Рэй, Дж. И др. (2016). Сон и нейрохимическая модуляция с помощью ядерного рецептора, активируемого пролифератором пероксисом, альфа (PPAR-альфа) у крыс. Neurosci. Res. 105, 65–69. DOI: 10.1016 / j.neures.2015.09.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Михангос-Морено, С., Поот-Аке, А., Сарро-Рамирес, А., Хименес-Морено, Р., Пачеко-Пантоха, Э., Акино-Эрнандес, П. и др. (2015). Циркадные и депривационные вариации монофосфорилированной МАР-киназы в гипоталамусе и мостах крыс. Biol. Rhythm Res. 46, 763–770. DOI: 10.1080 / 092.2015.1052651
CrossRef Полный текст | Google Scholar
Мурильо-Родригес, Э., Бланко-Центурион, К., Геращенко, Д., Салин-Паскуаль, Р. Дж., И Широмани, П. Дж. (2004). Суточный ритм уровня аденозина в базальной части переднего мозга молодых и старых крыс. Неврология 123, 361–370. DOI: 10.1016 / j.neuroscience.2003.09.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мурильо-Родригес, Э., Бланко-Центурион, К., Санчес, К., Пиомелли, Д., и Широмани, П. Дж. (2003). Анандамид увеличивает внеклеточные уровни аденозина и вызывает сон: исследование микродиализа in vivo. Сон 26, 943–947. DOI: 10,1093 / сон / 26.8.943
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мурильо-Родригес, Э., Кабеса, Р., Мендес-Диас, М., Наварро, Л., и Просперо-Гарсия, О. (2001). Анандамид-индуцированный сон блокируется SR141716A, антагонистом рецептора CB1, и U73122, ингибитором фосфолипазы C. Neuroreport 12, 2131–2136. DOI: 10.1097 / 00001756-200107200-00018
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мурильо-Родригес, Э., Аро, Р., Паломеро-Риверо, М., Миллан-Альдако, Д., и Друкер-Колин, Р. (2007a). Модафинил увеличивает внеклеточные уровни дофамина в прилежащем ядре и увеличивает бодрствование у крыс. Behav. Brain Res. 176, 353–357.
PubMed Аннотация | Google Scholar
Мурильо-Родригес, Э., Мачадо, С., Роча, Н. Б., Бадде, Х., Юань, Т. Ф. и Ариас-Падаль, О. (2016). Выявление роли модуляторов эндоканнабиноидной системы SR141716A, URB597 и VDM-11 в гомеостазе сна. Неврология 339, 433–449. DOI: 10.1016 / j.neuroscience.2016.10.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мурильо-Родригес, Э., Миллан-Альдако, Д., Ди Марцо, В., и Друкер-Колин, Р. (2008a). Ингибитор мембранного переносчика анандамида, VDM-11, модулирует сон и экспрессию c-Fos в головном мозге крысы. Неврология 157, 1–11. DOI: 10.1016 / j.neuroscience.2008.08.056
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мурильо-Родригес, Э., Миллан-Алдако, Д., Паломеро-Риверо, М., Мешулам, Р., и Друкер-Колин, Р. (2008b). Каннабидиол, непсихоактивный компонент каннабиса, вызывает пробуждение. Behav. Neurosci. 122, 1378–1382. DOI: 10.1037 / a0013278
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мурильо-Родригес, Э., Миллан-Альдако, Д., Паломеро-Риверо, М., Мешулам, Р., и Друкер-Колин, Р. (2006). Каннабидиол, входящий в состав Cannabis sativa, регулирует сон у крыс. FEBS Lett. 580, 4337–4345. DOI: 10.1016 / j.febslet.2006.04.102
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мурильо-Родригес, Э., Паломеро-Риверо, М., Миллан-Альдако, Д., Ариас-Каррион, О., и Друкер-Колин, Р. (2011a). Введение URB597, олеоилэтаноламида или пальмитоилэтаноламида увеличивает пробуждение и дофамин у крыс. PLoS ONE 6: e20766. DOI: 10.1371 / journal.pone.0020766
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мурильо-Родригес, Э., Паломеро-Риверо, М., Миллан-Альдако, Д., и Ди Марцо, В. (2013). Введение ингибиторов захвата эндоканнабиноидов OMDM-2 или VDM-11 способствует сну и снижает внеклеточные уровни дофамина у крыс. Physiol. Behav. 109, 88–95. DOI: 10.1016 / j.physbeh.2012.11.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мурильо-Родригес, Э., Паломеро-Риверо, М., Миллан-Альдако, Д., Мешулам, Р., и Друкер-Колин, Р. (2011b). Влияние на сон и уровни дофамина микродиализной перфузии каннабидиола в боковой гипоталамус крыс. Life Sci. 88, 504–511. DOI: 10.1016 / j.lfs.2011.01.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мурильо-Родригес, Э., Санчес-Алавес, М., Наварро, Л., Мартинес-Гонсалес, Д., Друкер-Колин, Р., и Просперо-Гарсия, О. (1998). Анандамид модулирует сон и память у крыс. Brain Res. 812, 270–274. DOI: 10.1016 / S0006-8993 (98) 00969-X
CrossRef Полный текст | Google Scholar
Мурильо-Родригес, Э., Васкес-Луис, Э., Миллан-Алдако, Д., Аро, Р. и Друкер-Колин, Р. (2008c). Стимулирование бодрствования введением модафинила в передний гипоталамус и в ядро покрышки ножки и папоротника у крыс. Neurosci. Lett. 436, 283–287. DOI: 10.1016 / j.neulet.2008.03.048
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мурильо-Родригес, Э., Васкес, Э., Миллан-Альдако, Д., Паломеро-Риверо, М., и Друкер-Колин, Р. (2007b). Влияние ингибитора амидгидролазы жирных кислот URB597 на цикл сна-бодрствование, экспрессию c-Fos и уровни дофамина у крыс. Eur. J. Pharmacol. 562, 82–91.
PubMed Аннотация | Google Scholar
Наваррия, А., Тамбурелла, А., Яннотти, Ф. А., Микале, В., Камиллиери, Г., Гоццо, Л., и др. (2014). Двойной блокатор N-арахидоноилсеротонина FAAH / TRPV1 обращает вспять поведенческое отчаяние, вызванное стрессом у крыс, и модулирует ось HPA. Pharmacol. Res. 87, 151–159. DOI: 10.1016 / j.phrs.2014.04.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Нимчик, М., и Деккер, М. (2015). Новые подходы к конструированию и разработке лигандов каннабиноидных рецепторов: многофункциональных и двухвалентных соединений. ChemMedChem. 10, 773–786. DOI: 10.1002 / cmdc.201500041
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Палаццо, Э., Луонго, Л., Новеллис, В., Росси, Ф., и Майоне, С. (2010). Роль каннабиноидных рецепторов в нисходящей модуляции боли. Pharmaceuticals 3, 2661–2673. DOI: 10.3390 / ph4082661
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Панлилио, Л.В., Торндайк, Э. Б., Никас, С. П., Алапафуджа, С. О., Бандиера, Т., Краватт, Б. Ф. и др. (2016). Влияние ингибиторов амидгидролазы жирных кислот (FAAH) на рабочую память у крыс. Психофармакология 233, 1879–1888. DOI: 10.1007 / s00213-015-4140-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пава, М. Дж., Ден Хартог, К. Р., Бланко-Центурион, К., Широмани, П. Дж., И Вудворд, Дж. Дж. (2014). Эндоканнабиноидная модуляция корковых состояний и медленный сон. PLOS ONE 9: e88672. DOI: 10.1371 / journal.pone.0088672
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Paxinos, G., and Watson, C. (2005). Мозг крысы в стереотаксических координатах. Сан-Диего, Калифорния: Academic Press.
Google Scholar
Пембер, С. О., Мехиа, Г. Л., Прайс, Т. Дж., И Пастерис, Р. Дж. (2016). Изоксазолины пиперидинилтиазола: новая серия высокоэффективных, медленно обратимых ингибиторов FAAH с анальгетическими свойствами. Bioorg. Med. Chem. Lett. 26, 2965–2973. DOI: 10.1016 / j.bmcl.2016.02.061
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Поркка-Хейсканен, Т., Стрекер, Р. Э. и Маккарли, Р. В. (2000). Сайт-специфичность изменений внеклеточной концентрации аденозина в мозге во время депривации сна и спонтанного сна: исследование микродиализа in vivo. Неврология 99, 507–517. DOI: 10.1016 / S0306-4522 (00) 00220-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цю, М.Х., Лю В., Цюй В. М., Ураде Ю., Лу Дж. И Хуанг З. Л. (2012). Роль ядра / оболочки прилежащего ядра в регуляции сна и бодрствования и их участие в возбуждении, индуцированном модафинилом. PLoS ONE 7: e45471. DOI: 10.1371 / journal.pone.0045471
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Руэда-Ороско, П. Э., Сориа-Гомес, Э., Монтес-Родригес, К. Дж., Перес-Моралес, М., и Просперо-Гарсия, О. (2010). Внутригиппокампальное введение анандамида увеличивает фазу быстрого сна. Neurosci. Lett. 473, 158–162. DOI: 10.1016 / j.neulet.2010.02.044
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Santucci, V., Storme, J. J., Soubrie, P., and Le Fur, G. (1996). Усиливающие возбуждение свойства антагониста каннабиноидного рецептора CB1 SR 141716A у крыс по оценке с помощью электроэнцефалографического спектрального анализа и анализа цикла сна-бодрствования. Life Sci. 58, L103 – L110. DOI: 10.1016 / 0024-3205 (95) 02319-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сартим, А.Г., Морейра Ф.А. и Джока С. Р. (2017). Вовлечение рецепторов CB1 и TRPV1, расположенных в вентральной медиальной префронтальной коре, в модуляции поведения по преодолению стресса. Неврология 340, 126–134. DOI: 10.1016 / j.neuroscience.2016.10.031
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сартори, Т., Сейги Мураками, Ф., Пинейро Крус, А., и Мачадо де Кампос, А. (2008). Разработка и валидация метода быстрой ОФ-ВЭЖХ для определения эффективности захвата метотрексата полимерными нанокапсулами. J. Chromatogr. Sci. 46, 505–509. DOI: 10.1093 / chromsci / 46.6.505
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Скаммелл, Т. Е., Эстабрук, И. В., Маккарти, М. Т., Чемелли, Р. М., Янагисава, М., Миллер, М. С. и др. (2000). Области возбуждения гипоталамуса активируются во время бодрствования, вызванного модафинилом. J. Neurosci. 20, 8620–8628.
PubMed Аннотация | Google Scholar
Старович К., Макух В., Осикович М., Piscitelli, F., Petrosino, S., Di Marzo, V., et al. (2012). Спинальный анандамид вызывает обезболивание у нейропатических крыс: возможные CB (1) – и TRPV1-опосредованные механизмы. Нейрофармакология 62, 1746–1755. DOI: 10.1016 / j.neuropharm.2011.11.021
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сухал С., Халид М., Тулаймат А. (2015). Влияние средств, способствующих бодрствованию, на сонливость у пациентов с апноэ во сне, получавших CPAP: метаанализ. J. Clin. Sleep Med. 11, 1179–1186. DOI: 10.5664 / jcsm.5096
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Sun, L., Tai, L., Qiu, Q., Mitchell, R., Fleetwood-Walker, S., Joosten, E.A., et al. (2017). Эндоканнабиноидная активация рецепторов CB1 способствует длительному обращению невропатической боли за счет повторяющейся стимуляции спинного мозга. Eur. Дж. Пейн 21, 804–814. DOI: 10.1002 / ejp.983
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сундуру, Н., Свенссон, М., Сиприано, М., Марваха, С., Андерссон, К. Д., Свенссон, Р. и др. (2017). N-арил-2-арилоксиацетамиды как новый класс ингибиторов амидгидролазы жирных кислот (FAAH). J. Enzyme Inhib. Med. Chem. 32, 513–521. DOI: 10.1080 / 14756366.2016.1265520
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Светник, В., Снайдер, Э. С., Ма, Дж., Тао, П., Лайнс, К., и Херринг, В. Дж. (2017). Спектральный анализ ЭЭГ медленного сна в большой выборке пациентов с бессонницей и хорошим сном: влияние возраста, пола и продолжительности ночи. J. Sleep Res. 26, 92–104. DOI: 10.1111 / jsr.12448
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тоблер И. и Борбели А. А. (1990). Влияние 3-х и 6-ти часового лишения сна на сон и спектры ЭЭГ крысы. Behav. Brain Res. 36, 73–78. DOI: 10.1016 / 0166-4328 (90)-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тонг, Дж., Мизрахи, Р., Хоул, С., Киш, С. Дж., Буало, И., Нобрега, Дж. И др.(2016). Ингибирование амидгидролазы жирных кислот с помощью BIA 10-2474 в головном мозге крысы. J. Cereb. Blood Flow Metab. DOI: 10.1177 / 0271678X16668890
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тот, А., Блумберг, П. М., и Бокзан, Дж. (2009). Анандамид и ваниллоидный рецептор (TRPV1). Vitam Horm. 81, 389–419. DOI: 10.1016 / S0083-6729 (09) 81015-7
CrossRef Полный текст | Google Scholar
Tuo, W., Leleu-Chavain, N., Barczyk, A., Renault, N., Lemaire, L., Chavatte, P., et al. (2016). Дизайн, синтез и биологическая оценка сильнодействующих ингибиторов FAAH. Bioorg. Med. Chem. Lett. 26, 2701–2705. DOI: 10.1016 / j.bmcl.2016.04.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вилела, Л. Р., Медейрос, Д. К., де Оливейра, А. К., Мораес, М. Ф., и Морейра, Ф. А. (2014). Противосудорожные эффекты N-арахидоноил-серотонина, гидролазы двойного амида жирных кислот. (фермент) и блокатор каналов временного рецепторного потенциала ваниллоидного типа-1 (TRPV1) при экспериментальных припадках: роль каннабиноидных рецепторов CB1 и каналов TRPV1. Basic Clin. Pharmacol. Toxicol. 115, 330–334. DOI: 10.1111 / bcpt.12232
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wang, D., Bai, X. X., Williams, S. C., Hua, S. C., Kim, J. W., Marshall, N. S., et al. (2015). Модафинил увеличивает активацию ЭЭГ в бодрствующем состоянии и улучшает показатели при обструктивном апноэ во сне во время непрерывной отмены положительного давления в дыхательных путях. Сон 38, 1297–1303. DOI: 10.5665 / sleep.4912
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вилкерсон, Дж.Л., Гош, С., Мустафа, М., Абдулла, Р. А., Нифакис, М. Дж., Кабрера, Р. и др. (2017). Ингибитор гидролиза эндоканнабиноидов SA-57: внутренние антиноцицептивные эффекты, усиление антиноцицепции, индуцированной морфином, и ослабление поведения, связанного с поиском героина у мышей. Нейрофармакология 114, 156–167. DOI: 10.1016 / j.neuropharm.2016.11.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Винклер К., Рамер Р., Дитмер С., Иванов И., Меркорд Дж. И Хинц Б.(2016). Ингибиторы амидгидролазы жирных кислот оказывают противоинвазивное и антиметастатическое действие на клетки рака легких. Oncotarget 7, 15047–15064. DOI: 10.18632 / oncotarget.7592
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уолланк, Ю., Рамер, Р., Иванов, И., Саламон, А., Петерс, К., и Хинц, Б. (2015). Ингибирование FAAH увеличивает миграцию стволовых клеток через PPARalpha. J. Lipid Res. 56, 1947–1960. DOI: 10.1194 / мл. M061473
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Захов, С., Гарзинский Д., Ханекамп В. и Лер М. (2017). 1-Гетероарилпропан-2-оны как ингибиторы амидгидролазы жирных кислот: исследования взаимосвязей структура-активность и метаболической стабильности. Bioorg. Med. Chem. 25, 825–837. DOI: 10.1016 / j.bmc.2016.11.025
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Zhang, J. P., Xu, Q., Yuan, X. S., Cherasse, Y., Schiffmann, S. N., De Kerchove D’exaerde, A., et al. (2013). Проекции нейронов аденозинового рецептора A2A ядра прилежащего ядра в мозге мышей и их значение в опосредовании регуляции сна и бодрствования. Перед. Нейроанат. 7:43. DOI: 10.3389 / fnana.2013.00043
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рекомендации USPSTF по скринингу на АГ у взрослых
Из этой рекомендации по скринингу на гипертонию (АГ) у взрослых, которую следует запомнить, из Целевой группы США по профилактическим службам следует помнить:
- Определение HTN варьируется в зависимости от организации. АГ определяется как ≥140/90 мм рт. Ст. Или ≥130/80 мм рт. Ст.
- Все взрослые должны пройти скрининг на АГ, факторы риска, повышающие риск заболевания, и диетические факторы.
- Факторы риска включают пожилой возраст, чернокожую расу, семейный анамнез, избыточный вес и ожирение, недостаток физической активности, стресс и употребление табака.
- К диетическим факторам относятся диета с высоким содержанием жира или натрия, диета с низким содержанием калия или чрезмерное употребление алкоголя.
- Research поддерживает получение офисных измерений артериального давления на плечевой артерии (плече), когда пациент находится в сидячем положении после 5 минут отдыха и медицинский персонал присутствует во время измерения.
- Амбулаторный мониторинг артериального давления и домашний мониторинг артериального давления с помощью проверенных и точных устройств следует использовать вне клинических условий для подтверждения диагноза.
- Скрининг АГ следует проводить ежегодно у взрослых старше 40 лет и взрослых с повышенным риском АГ.
- Скрининг каждые 3-5 лет подходит для взрослых 18-39 лет, которые не относятся к группе повышенного риска АГ и имеют подтвержденное нормальное кровяное давление.
- Лечение может включать изменение образа жизни, фармакотерапию или и то, и другое.Лечение должно основываться на возрасте, факторах риска и тяжести АГ.
- В сопутствующей редакционной статье отмечается, что коронавирусная болезнь 2019 повлияла на способность проверять группы высокого риска на АГ и высветила расовые / этнические различия в состоянии здоровья.
Клинические темы: Диабет и кардиометаболические заболевания, профилактика, диета, гипертония
Ключевые слова: Гипертония, Артериальное давление, Мониторинг артериального давления, амбулаторно, Определение артериального давления, Взрослый, Факторы риска, Диета, Сидячий образ жизни4, Натрий , Ожирение, афроамериканцы, Употребление алкоголя, Употребление табака, Калий, COVID-19, тяжелый острый респираторный синдром, коронавирус 2
<Вернуться к спискам
.
