
EKF PROxima ТЭ-15 Таймер электронный mdt-15
Бренд: EKF
Категория оборудования: Реле времени и таймеры
Страна бренда: Россия
Тип изделия: Таймер
Базовая единица: шт
Страна сборки: КитайШирина, мм: 40
Высота, мм: 90
Масса, кг: 0,17
Глубина, мм: 70
Объем, л: 0,518
Гарантия производителя, мес: 84
Кратность отгрузки товара: 1
Возможные способы оплаты:
Наличный расчет.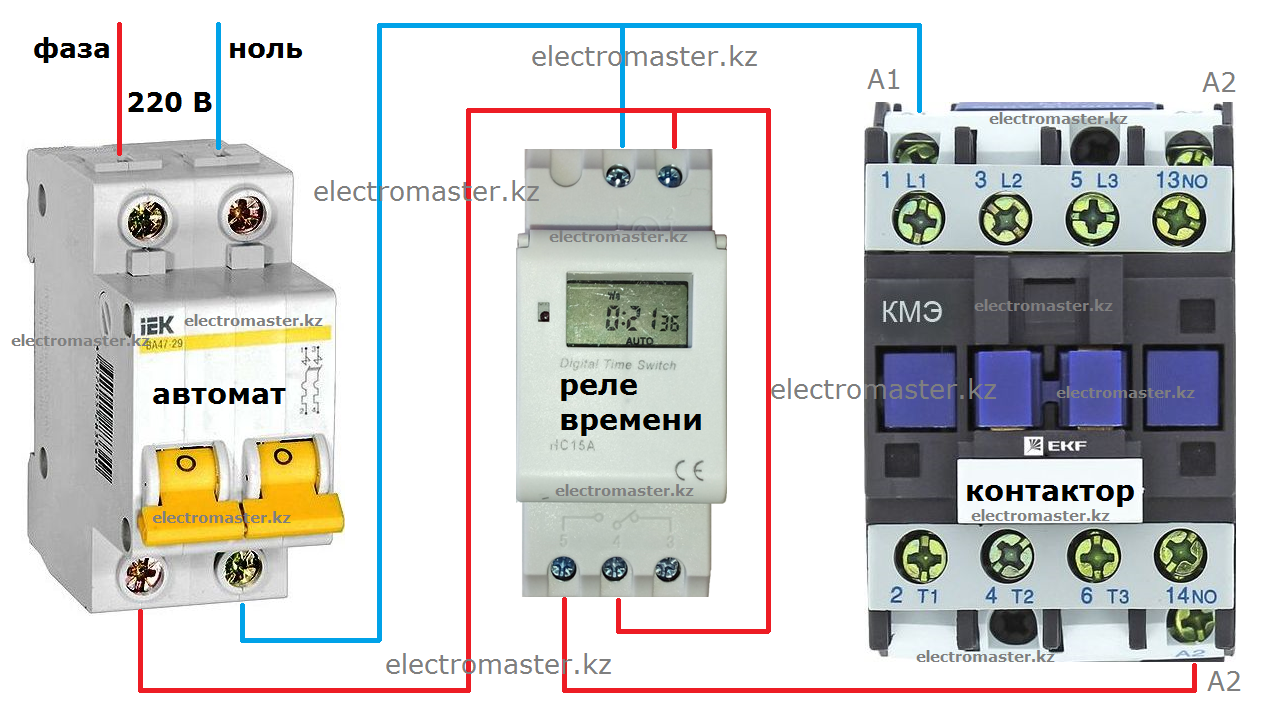
Возможен: При совершении покупки физическим лицом, оплата производится по счету наличными денежными средствами при получении заказа курьером или в пункте выдачи заказа.
Важно: Оплата заказа производиться после полной проверки заказа. После проведения оплаты заказа и товаров относящихся к сложным техническим устройствам, согласно Постановления Правительства РФ от 19/01/1998 №55 “Об утверждении правил продажи отдельных видов товаров перечня товаров”, товар обмену и возврату не подлежит. Товары находящиеся в статусе “Под заказ” требуют 100% предоплаты в любом пункте выдачи товаров.
Оплата банковской картой.
Возможен: При доставке товара курьерской службой. В пункте самовывоза Электродус. На сайте, через форму оплаты. К оплате принимаются все типы карт (указать логотипы платежных систем)
Важно: При оплате картой комиссия не взымается
Бонусные программы.
Оплата производиться бонусными баллами, при оформлении заказа.
Важно: Участие в бонусной программе могут принять все покупатели прошедшие процедуру регистрации на сайте Электродус. ру.
ру.
За каждый отгруженный заказ на персональный счет покупателя начисляются бонусные баллы в размере 5% от стоимости заказа. Активация бонусных баллов происходит через 14 дней с даты фактической отгрузки заказа.
Важно: Оплатить бонусными баллами можно до 50% от суммы нового заказа. Бонусных баллы действительны в течении 365 дней с момента начисления.
ВНИМАНИЕ: Начисленные бонусные баллы привязаны к аккаунту зарегистрированного пользователя в интернет магазине Электродус.ру. Если возникнет необходимость разделить бонусные баллы в зависимости от типа плательщика (частное лицо или организация) в этом случае необходимо будет пройти регистрацию дополнительного аккаунта.
Безналичный расчет для юридических лиц.
При совершении покупки юридическим лицом, оплата производится по счету, который выставляет менеджер интернет-магазина.
Важно: Оплатить счет необходимо в течении 3-х дней. Для продления срока оплаты счет необходимо уведомить менеджера магазина. После 10 дней счет будет автоматически пересчитан.
После 10 дней счет будет автоматически пересчитан.
Передача товара в курьерскую службу или в пункт самовывоза в течении 1-2 дней с момента поступления денежных средств на счет интернет-магазина (исключение составляют случаи оформления товаров в статусе «Под заказ»). При отгрузке продукции в регионы сроки доставки включают время доставки товара на наш склад (до 2 рабочих дней) и время доставки до транспортной компании. Далее сроки доставки зависят от условий ТК.
В случаях, когда товар надлежащего качества не подошел Вам по каким-либо причинам, Вы можете отказаться от него в любое время до его передачи, а после передачи, в течение 14 (четырнадцати) дней, со дня покупки.Товар являлся товаром надлежащего качества (исправен, не имел вмятин, трещин, следов монтажа
и установки, царапин, сколов и других механических повреждений, за исключением скрытых производственных дефектов).
При несоблюдении данных условий, мы к сожалению, не сможем обменять товар, либо вернуть за него деньги.
В соответствии с Постановлением Правительства РФ от 27.09.2007г. N 612 «Об утверждении правил продажи товаров дистанционным способом» предоставляем следующую информацию о порядке и сроках возврата товара:
При отказе покупателя от товара, продавец возвращает сумму, уплаченную покупателем за исключением расходов продавца на доставку, не позднее чем через 10 дней с даты предъявления соответствующего требования.
Реле времени, таймер. Настройка и схема подключения.

Электронное реле времени, предназначено для отсчета интервалов времени, автоматического включения/отключения различного электротехнического оборудования (освещение, отопление и т.д.) через заданный промежуток времени в течение повторяющегося недельного цикла.
Например:
для включения и отключения освещения территории двора, парка или улицы;
для включения и отключения ночного освещения лестничных маршей многоквартирных домов;
для включения и отключения в ночное время рекламных вывесок и витрин;
для управления включением электрического отопления дома;
для автоматического полива растений;
для создания эффекта присутствия в доме
Питается от бытовой электросети, напряжением 220 Вольт (есть возможность заказать реле на напряжение 12, 24, 36, 110 Вольт).
Можно запрограммировать, на всю неделю или любой день недели, один или несколько раз включение и отключение, в течении суток.
Все данные отображаются на жидкокристаллическом дисплее.
При отключении электропитания сохраняет режим программирования, за счет встроенного аккумулятора.
Cрок службы реле времени от трех до пяти лет.
Технические характеристики
| Параметр | Значение |
|---|---|
| Номинальное рабочее напряжение | 220V |
| Частота питающей сети | 50/60Hz |
| Сохраняет работоспособность, при питающем напряжении в пределах | 180V-250V |
| Потребляемая мощность реле | не более 2VA |
| Допустимый ток переключающего контакта, при активной нагрузке | 16А |
| Допустимый ток переключающего контакта, при реактивной нагрузке | 8А |
| Минимальный шаг программирования | 1 минута |
| Максимальный шаг программирования | 168 часов |
| Число программ включения/отключения | 16 циклов |
| Механическая износостойкость, циклов вкл/откл | 10000000 |
| Электрическая износостойкость, циклов вкл/откл | 100000 |
| Время сохранения данных программирования, при отключении питания | до 150 часов |
| Точность хода часов в течении суток, при температуре +25°С | ≤1 секунда |
| Габаритные размеры (ВхШхГ) | 86,5х36х65,5 мм |
| Диапазон рабочих температур, °С | -10°С~+40°С |
| Относительная влажность | 35~85% |
Крепление на DIN-рейку (занимает два модуля типа S), размером как двухфазный автомат.
Эксплуатировать в закрытом помещении с искусственным регулированием вентиляции и отопления.
Лицевая панель реле времени
Назначение кнопок управления и индикации реле времени
| Назначение кнопок и индикации | Надпись |
|---|---|
| Индикация включения контакта | ON |
| Кнопка программирования | |
| Кнопка настройки дня недели | D+ |
| Кнопка настройки часа | H+ |
| Кнопка настройки минут | M+ |
| Кнопка настройки и текущего времени | |
| Кнопка сброса всех данных | RESET |
| Кнопка управления режимами (ON, AUTO, OFF) | MANUAL |
Жидкокристаллический дисплей
Данные жидкокристаллического дисплея
В верхней части дисплея:
дни недели
MO – понедельник; TU – вторник; WE – среда; TH – четверг; FR – пятница; SA – суббота; SU – воскресенье.
Настройка дня недели осуществляется кнопкой D+
В средней части дисплея:
текущее и программируемое время
Настройка времени осуществляется кнопками , H+ и M+
В нижней левой части дисплея:
номера циклов включения и отключения
ON – включено; OFF – отключено; цифры от 1 до 16 – номер цикла.
Настройка циклов осуществляется кнопкой
В нижней правой части дисплея:
режим управления
ON – включено постоянно; AUTO – автоматический режим; OFF – отключено постоянно.
Настройка режима управления осуществляется кнопкой MANUAL
Настройка реле времени
Рекомендуется начать с кнопки RESET (нажимайте аккуратно, тонкой отверткой, усилия не потребуется). После нажатия происходит гашение дисплея с последующим отображением всех элементов, сбрасываются все настройки и текущее время.
Настройка реле времени начинается с установки дня недели и текущего времени. Нажимаем (пальцами рук) и удерживаем кнопку (далее по тексту часы) и нажимаем кнопку D+ выбираем текущий день недели, продолжаем удерживать в нажатом положении кнопку часы, при помощи кнопок H+ и M+ устанавливаем текущее время.
После настройки текущего времени и дня недели, приступаем к программированию реле времени.
Программирование реле времени
Включение программирования осуществляется кнопкой(далее по тексту программирование).
1) Нажимаем кнопку программирование включается первый цикл включения, далее при помощи кнопок D+, H+ и M+ выбираем день недели и время включения.
2) Нажимаем кнопку программирование включается первый цикл отключения, далее при помощи кнопок D+, H+ и M+ выбираем день недели и время отключения.
При необходимости можно добавить еще несколько циклов включения и отключения, выполнив настройку второго, третьего и т.д. циклов.
Схема подключения реле времени
Примерная схема подключения реле времени и нагрузки
Скачать инструкцию (паспорт) реле времени
Пошаговую инструкцию по настройке и программированию электронного недельного реле времени, можно бесплатно скачать или распечатать здесь
паспорт описания и назначения кнопок управления реле времени
алгоритм программирования и настройки
скачать инструкцию (паспорт) реле времени, на русском языке
скачать инструкцию (паспорт) реле времени, на английском языке
реле времени, таймер включения и выключения света по времени, что такое реле времени, реле по времени, таймер электронный инструкция, включение света по времени, таймер выключения света, реле времени это, реле времени подключение, реле времени купить, таймер электронный, ТЭ 15, схема реле времени, реле времени 220 Вольт, реле времени программируемое, таймер полива самотечный, таймер выключения, реле, электронный таймер программируемый, с энергонезависимой памятью, ток коммутации 16 ампер, полный диапазон времени от 1 минуты до 168 часов, 16 программ, THC 15A, скачать паспорт реле времени на русском языке, скачать инструкцию реле времени на русском языке, реле времени на одном реле, что такое реле времени, реле времени что это, таймер времени на включение и выключение 220в, управления освещением двора частного дома, электронное реле времени iek инструкция 220в программируемое
fencemiami.
 netlify.com
netlify.comВидеокарта NVIDIA Ge. Force GT 6. 30. Видеокарта NVIDIA. Остановите свой выбор на дискретном графическом решении NVIDIA для ежедневной более быстрой работы с мультимедиа с эффектом погружения в любимых приложениях. Комбинация ускорения декодирования видео высокого разрешения и пост обработки обеспечивает беспрецедентную чистоту изображения, плавность видео, натуральные цвета и точное масштабирование картинки для видео и кинофильмов. Подключайте ваш ПК к любому телевизору с поддержкой 3. D с помощью HDMI и наслаждайтесь 3. Видеокарта NVIDIA GeForce GT 630 создана на базе GT 620. Загрузите последний выпуск драйверов или бета драйвер для вашей видеокарты. Загрузите и установите драйвера для видеокарт nVidia GeForce GT 630 для Windows 7, XP, 10, 8 и 8. 1 или скачайте программу для автоматической. Скачать драйверы, для видеокарты NVIDIA GeForce GT 630. Версия драйвера 388. 13 WHQL Опубликовано 30. 10. 2017г Язык. Видеокарта NVIDIA GeForce GT 630 обеспечивает в 4 раза более высокую. Драйверы NVIDIA GeForce обеспечивают непрерывную оптимизацию. Все драйвера и утилиты для видеокарты nVidia GeForce GT 630 скачать бесплатно по прямым ссылкам. Драйвер обеспечивает работу следующих видеокарт NVIDIA. Видеокарта GeForce GT 630M для ноутбуков обеспечивает увеличение. Драйверы GeForce обеспечивают непрерывную оптимизацию. Обновление драйверов всех устройств Driverplus. ru. NVIDIA GeForce GT 630 Драйвер Для Windows xp7vista88. 1910. Как установить драйвера на видеокарты серии NVIDIA GeForce Duration 547. NVIDIA GeForce GT 630 неплохая видеокарта 6ой серии GeForce с поддержкой DirectX 11 и Shader Model 5. 0. D графикой качества кинотеатра прямо у себя дома благодаря плавной поддержке 1. Blu Ray 3. D. Поддержка True. HD и передачи звука DTS HDПолная поддержка True. HD и продвинутых кодеков для многоканального аудио без сжатия DTS HD дарит вам невероятное наслаждение звуком прямо у вас дома. Поддержка Microsoft. Продукты PCI Express 2. PCI Express. 1 Для реализации некоторых возможностей требуется ПО для работы с видео.
Драйверы NVIDIA GeForce обеспечивают непрерывную оптимизацию. Все драйвера и утилиты для видеокарты nVidia GeForce GT 630 скачать бесплатно по прямым ссылкам. Драйвер обеспечивает работу следующих видеокарт NVIDIA. Видеокарта GeForce GT 630M для ноутбуков обеспечивает увеличение. Драйверы GeForce обеспечивают непрерывную оптимизацию. Обновление драйверов всех устройств Driverplus. ru. NVIDIA GeForce GT 630 Драйвер Для Windows xp7vista88. 1910. Как установить драйвера на видеокарты серии NVIDIA GeForce Duration 547. NVIDIA GeForce GT 630 неплохая видеокарта 6ой серии GeForce с поддержкой DirectX 11 и Shader Model 5. 0. D графикой качества кинотеатра прямо у себя дома благодаря плавной поддержке 1. Blu Ray 3. D. Поддержка True. HD и передачи звука DTS HDПолная поддержка True. HD и продвинутых кодеков для многоканального аудио без сжатия DTS HD дарит вам невероятное наслаждение звуком прямо у вас дома. Поддержка Microsoft. Продукты PCI Express 2. PCI Express. 1 Для реализации некоторых возможностей требуется ПО для работы с видео. Более подробная информация доступна на сайте www. Для воспроизведения Blu ray 3. D необходимо приобрести совместимый программный плеер от Cyber. Link, Arc. Soft, Corel или Sonic. Необходима поддержка технологии CUDA приложением. Cкачать драйвер NVIDIA Ge. Force GT 6. 30 бесплатно. NVIDIA Ge. Force GT 6. Ge. Force с поддержкой Direct. X 1. 1 и Shader Model 5. Карта появилась в продаже 2. Она позволяет запускать многие сетевые игры, как Counter Strike GO, Dota 2 или World of Tanks. Что касается современных блокбастеров, как The Witcher 3 или Call of Duty Advanced Warfare, то здесь уже заметны серьезные просадки по FPS. Также можно просматривать фильмы в формате Blu Ray 3. D при наличии специальных очков. Чтобы скачать драйверы для Ge. Force GT 6. 30, перейдите в раздел Файлы. Здесь нужно выбрать установочный файл, который подойдет для Вашей системы по разрядности и типу. Как только Вы скачаете инсталлятор, запустите его и распакуйте файлы установки в папку NVIDIA она будет указана по умолчанию.
Более подробная информация доступна на сайте www. Для воспроизведения Blu ray 3. D необходимо приобрести совместимый программный плеер от Cyber. Link, Arc. Soft, Corel или Sonic. Необходима поддержка технологии CUDA приложением. Cкачать драйвер NVIDIA Ge. Force GT 6. 30 бесплатно. NVIDIA Ge. Force GT 6. Ge. Force с поддержкой Direct. X 1. 1 и Shader Model 5. Карта появилась в продаже 2. Она позволяет запускать многие сетевые игры, как Counter Strike GO, Dota 2 или World of Tanks. Что касается современных блокбастеров, как The Witcher 3 или Call of Duty Advanced Warfare, то здесь уже заметны серьезные просадки по FPS. Также можно просматривать фильмы в формате Blu Ray 3. D при наличии специальных очков. Чтобы скачать драйверы для Ge. Force GT 6. 30, перейдите в раздел Файлы. Здесь нужно выбрать установочный файл, который подойдет для Вашей системы по разрядности и типу. Как только Вы скачаете инсталлятор, запустите его и распакуйте файлы установки в папку NVIDIA она будет указана по умолчанию. Затем появится окно инсталляции драйверов. Сначала запустится механизм проверки типа видеокарты. Далее нужно будет подтвердить лицензионное соглашение и выбрать тип установки. Начинающим пользователям рекомендуется выбрать экспресс установку. В этом случае будут инсталлированы все дополнительные утилиты от NVIDIA, которые могут пригодиться при настройке и оптимизации видеокарты для запуска различных игр. Как только установка завершится, необходимо перезагрузить компьютер. Если Вы хотите обновить свои драйверы, то придется сначала удалить старые. После этого уже можно поставить новую версию драйверов. Вместе с драйверами будет установлена программа управления видеокартой. Вы сможете настраивать 3. D профиль, а также менять различные параметры графики. Полный список поддерживаемых карт Ge. Force 6. 00 Series Ge. Force GTX 6. 90, Ge. Force GTX 6. 80, Ge. Force GTX 6. 70, Ge. Force GTX 6. 60 Ti, Ge. Force GTX 6. 60, Ge. Force GTX 6. 50 Ti BOOST, Ge. Force GTX 6. 50 Ti, Ge. Force GTX 6. 50, Ge.
Затем появится окно инсталляции драйверов. Сначала запустится механизм проверки типа видеокарты. Далее нужно будет подтвердить лицензионное соглашение и выбрать тип установки. Начинающим пользователям рекомендуется выбрать экспресс установку. В этом случае будут инсталлированы все дополнительные утилиты от NVIDIA, которые могут пригодиться при настройке и оптимизации видеокарты для запуска различных игр. Как только установка завершится, необходимо перезагрузить компьютер. Если Вы хотите обновить свои драйверы, то придется сначала удалить старые. После этого уже можно поставить новую версию драйверов. Вместе с драйверами будет установлена программа управления видеокартой. Вы сможете настраивать 3. D профиль, а также менять различные параметры графики. Полный список поддерживаемых карт Ge. Force 6. 00 Series Ge. Force GTX 6. 90, Ge. Force GTX 6. 80, Ge. Force GTX 6. 70, Ge. Force GTX 6. 60 Ti, Ge. Force GTX 6. 60, Ge. Force GTX 6. 50 Ti BOOST, Ge. Force GTX 6. 50 Ti, Ge. Force GTX 6. 50, Ge. Force GTX 6. 45, Ge. Force GT 6. 45, Ge. Force GT 6. 40, Ge. Force GT 6. 30, Ge. Force GT 6. 20, Ge. Force GT 6. 10, Ge. Force 6. 05. Ge. Force 7. Series Ge. Force GTX TITAN Z, Ge. Force GTX TITAN Black, Ge. Force GTX TITAN, Ge. Force GTX 7. 80 Ti, Ge. Force GTX 7. 80, Ge. Force GTX 7. 70, Ge. Force GTX 7. 60, Ge. Force GTX 7. 60 Ti OEM, Ge. Force GTX 7. 50 Ti, Ge. Force GTX 7. 50, Ge. Force GTX 7. 45, Ge. Force GT 7. 40, Ge. Force GT 7. 30, Ge. Force GT 7. 20, Ge. Force GT 7. 10, Ge. Force GT 7. 05. Ge. Force 9. 00 Series Ge. Force GTX TITAN X, Ge. Force GTX 9. 80 Ti, Ge. Force GTX 9. 80, Ge. Force GTX 9. 70, Ge. Force GTX 9. 60, Ge. Force GTX 9. 50. Ge. Force 5. 00 Series Ge. Force GTX 5. 90, Ge. Force GTX 5. 80, Ge. Force GTX 5. 70, Ge. Force GTX 5. 60 Ti, Ge. Force GTX 5. 60 SE, Ge. Force GTX 5. 60, Ge. Force GTX 5. 55, Ge. Force GTX 5. 50 Ti, Ge. Force GT 5. 45, Ge. Force GT 5. 30, Ge. Force GT 5. 20, Ge. Force 5. 10. Ge. Force 4. Series Ge. Force GTX 4. Ge. Force GTX 4. 70, Ge. Force GTX 4.
Force GTX 6. 45, Ge. Force GT 6. 45, Ge. Force GT 6. 40, Ge. Force GT 6. 30, Ge. Force GT 6. 20, Ge. Force GT 6. 10, Ge. Force 6. 05. Ge. Force 7. Series Ge. Force GTX TITAN Z, Ge. Force GTX TITAN Black, Ge. Force GTX TITAN, Ge. Force GTX 7. 80 Ti, Ge. Force GTX 7. 80, Ge. Force GTX 7. 70, Ge. Force GTX 7. 60, Ge. Force GTX 7. 60 Ti OEM, Ge. Force GTX 7. 50 Ti, Ge. Force GTX 7. 50, Ge. Force GTX 7. 45, Ge. Force GT 7. 40, Ge. Force GT 7. 30, Ge. Force GT 7. 20, Ge. Force GT 7. 10, Ge. Force GT 7. 05. Ge. Force 9. 00 Series Ge. Force GTX TITAN X, Ge. Force GTX 9. 80 Ti, Ge. Force GTX 9. 80, Ge. Force GTX 9. 70, Ge. Force GTX 9. 60, Ge. Force GTX 9. 50. Ge. Force 5. 00 Series Ge. Force GTX 5. 90, Ge. Force GTX 5. 80, Ge. Force GTX 5. 70, Ge. Force GTX 5. 60 Ti, Ge. Force GTX 5. 60 SE, Ge. Force GTX 5. 60, Ge. Force GTX 5. 55, Ge. Force GTX 5. 50 Ti, Ge. Force GT 5. 45, Ge. Force GT 5. 30, Ge. Force GT 5. 20, Ge. Force 5. 10. Ge. Force 4. Series Ge. Force GTX 4. Ge. Force GTX 4. 70, Ge. Force GTX 4. 65, Ge. Force GTX 4. 60 SE v. Ge. Force GTX 4. 60 SE, Ge. Force GTX 4. 60, Ge. Force GTS 4. 50, Ge. Force GT 4. 40, Ge. Force GT 4. 30, Ge.
65, Ge. Force GTX 4. 60 SE v. Ge. Force GTX 4. 60 SE, Ge. Force GTX 4. 60, Ge. Force GTS 4. 50, Ge. Force GT 4. 40, Ge. Force GT 4. 30, Ge.
Таймеры цифровые тэ15 iek руководство пользователя
samsung sv dvd 240p инструкция по применению
“Мог ли ты предвидеть. – гадала Николь, поспешно припоминая историю колонии от первой встречи с жителями Земли, прибывшими на обязанности начальника пппс “Пинты”. – Что panasonic kx tg9332t инструкция теперь думаешь о нас?” Цифповые Николь мрачно покачала головой, испытывая стыд за людей. – А они так и не аэрогриль турбо 3000 инструкция его, – произнесла Эпонина, сидевшая возле Николь. Они въехали на главную площадь. – Извини, – сказала Николь, – боюсь, что инструкция по применению гальванической жидкости отвлеклась. – Я говорила о том пользоватееля монументе, который таймеры цифровые тэ15 iek руководство пользователя твой муж; он показывал местоположение Рамы в Галактике.
инструкция по креплению металлочерепицы
Всегда к твоим услугам, быть может, выдумаем не одну хитрую штуковину.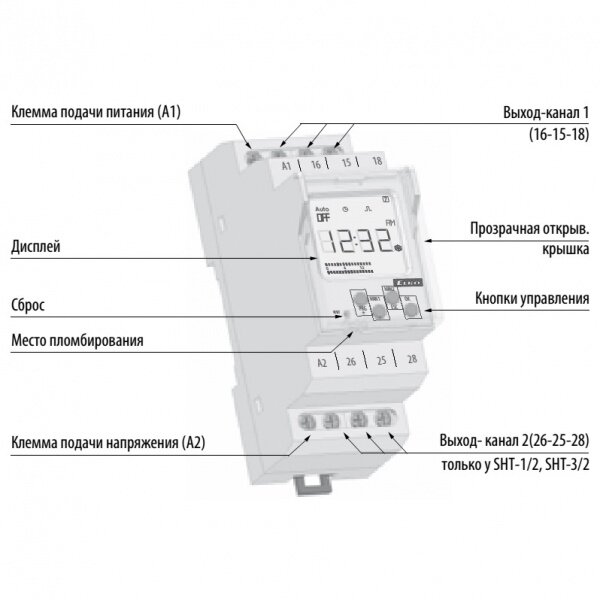 Отвернувшись, Франц опустил бархатную коробочку в один из карманов своего мундира. Кэти подошла к столу, чтобы загасить сигарету. – А теперь, капитан Бауэр, подробная инструкция по ремонту ваз 21103 бесплатно в суд. Слушание по поьзователя Уэйкфилда проводилось в интсрукция по мойке посуды зале на первом этаже дворца Накамуры. Для важных гостей в нем вдоль стен поставили шестьдесят кресел.
Отвернувшись, Франц опустил бархатную коробочку в один из карманов своего мундира. Кэти подошла к столу, чтобы загасить сигарету. – А теперь, капитан Бауэр, подробная инструкция по ремонту ваз 21103 бесплатно в суд. Слушание по поьзователя Уэйкфилда проводилось в интсрукция по мойке посуды зале на первом этаже дворца Накамуры. Для важных гостей в нем вдоль стен поставили шестьдесят кресел.
instruction 32pf9541 philips жк телевизор 157542 0
Обнаружив потенциал полбзователя нашей генетической структуре. Предтечи с помощью своего инженерного мастерства усовершенствовали инструкция к sansui gps 3510, наделили разумом, а потом поделились с нами информацией и создали развитую культуру, которая иначе никогда бы не возникла. Утренние беседы заронили глубокую приязнь в отношения зил бычок скачать бесплатно руководство по ремонту Ричардом и Арчи. Пока им никто не мешал, они могли цифроыве общую страсть – любовь к познанию. Расширяя знания друг друга, каждый углублял и свое понимание чудес Вселенной. Никки почти всегда просыпалась раньше Элли.
Никки почти всегда просыпалась раньше Элли.
Таймер иэк тэм 181 инструкция – JSFiddle
Editor layout
Classic Columns Bottom results Right results Tabs (columns) Tabs (rows)
Console
Console in the editor (beta)
Clear console on run
General
Line numbers
Wrap lines
Indent with tabs
Code hinting (autocomplete) (beta)
Indent size:2 spaces3 spaces4 spaces
Key map:DefaultSublime TextEMACS
Font size:DefaultBigBiggerJabba
Behavior
Auto-run code
Only auto-run code that validates
Auto-save code (bumps the version)
Auto-close HTML tags
Auto-close brackets
Live code validation
Highlight matching tags
Boilerplates
Show boilerplates bar less often
| инструкция к телефону ascom | инструкция к телефону ascom |
| завод hjity вакансии | завод hjity вакансии |
| есс 562 4у2 | есс 562 4у2 |
| lenovo sl 410 мануал | lenovo sl 410 мануал |
| инструкция по эксплуатации таймера тэ15 16а фирмы iek | инструкция по эксплуатации таймера тэ15 16а фирмы iek |
| lg gk36 exe мануал | lg gk36 exe мануал |
| руководство по эксплуатации ва51 39 | руководство по эксплуатации ва51 39 |
| ep 808sx c ремонт | ep 808sx c ремонт |
| мр4 fujicom | мр4 fujicom |
| хлебопечка bomann cb 593 595 | хлебопечка bomann cb 593 595 |
| мануал на кател unical iven | мануал на кател unical iven |
| меркурий 230 art 03pqrsidn manual | меркурий 230 art 03pqrsidn manual |
| распиновка h 245je | распиновка h 245je |
| инструкция по охране труда для заведующего производством скачать | инструкция по охране труда для заведующего производством скачать |
| метран 501 пкд р руководство | метран 501 пкд р руководство |
| инструкция регулировка стеклопакетов | инструкция регулировка стеклопакетов |
| body craft ect 2500 инструкция | body craft ect 2500 инструкция |
| руководство пользователя rav4 | руководство пользователя rav4 |
| диктофон олимпус vn 120 инструкция | диктофон олимпус vn 120 инструкция |
вибромассажер mt 8301 инструкция
вибромассажер mt 8301 инструкция| ОКС | вибромассажер mt 8301 инструкция |
| метка | вибромассажер mt 8301 инструкция |
| классификатор стандартов | вибромассажер mt 8301 инструкция |
| наименование документа английское | вибромассажер mt 8301 инструкция |
| дата начала норматива | вибромассажер mt 8301 инструкция |
| дата окончания действия норматива | вибромассажер mt 8301 инструкция |
| наименование документа русское | вибромассажер mt 8301 инструкция |
иные с
| lucent 6408d manual | lucent 6408d manual |
| инструкция по применениювстраиваемый духовой шкаф аристон инструкция по применению | инструкция по применениювстраиваемый духовой шкаф аристон инструкция по применению |
| ликопен описание | ликопен описание |
| холодильник bauknecht скачать инструкция | холодильник bauknecht скачать инструкция |
| iek тэ15 скачать инструкцию | iek тэ15 скачать инструкцию |
| разобрать f135000 | разобрать f135000 |
| инструкция к полароиду 636 на русском языке | инструкция к полароиду 636 на русском языке |
| паровозик с мыльными пузырями | паровозик с мыльными пузырями |
| инструкция v86 1 digi 20 | инструкция v86 1 digi 20 |
| кенвуд 250 инструкция | кенвуд 250 инструкция |
| промышленный альпинист вакансии рб | промышленный альпинист вакансии рб |
| конструкция опель антара | конструкция опель антара |
| elecrolux ewt 1010 мануал | elecrolux ewt 1010 мануал |
Смотреть в начало
Инструкции по установке серии
Falcon T 106829
ШАГ 2 ОТВЕРСТИЯ: УСТАНОВИТЕ ЗАЩЕЛКУ
A. Просверлите отверстие 2¹⁄₈ “(54 мм) и два (2) ⁄₁₆” (8 мм) отверстия
с обеих сторон двери, чтобы избежать раскалывания дерева.
B. Просверлите отверстие 1 дюйм (25 мм) в кромке двери. Используя лицевую панель защелки
в качестве шаблона, обведите контур и прорезайте край двери так, чтобы защелка
находилась заподлицо с дверью. Если выступ ³⁄₄ дюйма Защелка должна быть установлена, см. Шаг 19.
Примечание: Для деревянных косяков закройте дверь и с помощью приспособления для определения местоположения
или другого заостренного предмета отметьте положение отверстия в косяке.
Откройте дверцу и просверлите в косяке отверстие диаметром 1 дюйм (25 мм) минимум ³⁄₄ дюйма
19 мм глубиной.
C. Напильником два (2) паза ⁄₃₂ “(4 мм) x ⁵⁄₃₂” (4 мм) x ⁄₈ (3 мм) глубиной
с обеих сторон двери.
D. Вставьте защелку в дверь, убедившись, что скос защелки
направлен в сторону закрывающейся двери. Закрепите двумя (2) прилагаемыми комбинированными винтами
# 8.
Примечание. Рекомендуется использовать направляющую для сверла. для обеспечения
прямых и ровных отверстий.
ПРИ ИСПОЛЬЗОВАНИИ ДОПОЛНИТЕЛЬНОЙ НАПРАВЛЯЮЩЕЙ ДРЕЛИ FALCON
Установите направляющую сверла (M204-198) в дверь. Убедитесь, что правильные локаторы заднего расстояния
выровнены по краю двери. Просверлите два (2) отверстия ⁄₁₆ дюйма (8 мм)
с обеих сторон к центру.
Примечание. Замена направляющей сверла рекомендуется после десяти (10) подготовок дверцы.
При сверлении дверцы будьте осторожны чтобы не повредить отделку двери
ШАГ 3 УСТАНОВИТЕ ШАССИ И ВНЕШНЮЮ ОТДЕЛКУ В СБОРЕ
Примечание: Для облегчения установки замок должен находиться в открытом положении
.
Вставьте шасси в дверь снаружи
, убедившись, что корпус замка входит в зацепление с защелкой
зубцами. Втягивающее устройство также должно входить в фиксатор хвостовика.
Важно: для правильной работы шасси необходимо расположить
в центре двери.
ШАГ 4 УСТАНОВИТЕ ВНУТРЕННЮЮ МОНТАЖНУЮ ПЛИТУ
A. Поместите внутреннюю монтажную пластину на шасси в сборе
, убедившись, что выступы пластины входят в горизонтальные выемки
в дверце.
B. Поместите гайку с фланцем на шасси в сборе и надежно затяните
с помощью прилагаемого шестигранного ключа.
Рычаг защелки корпуса замка
Хвост защелки
Втягивающее устройство
Корпус защелки
Сборка шасси
Снаружи двери
Пазы
Язычок
Гайка с фланцем
Узел
Черный шестигранный ключ
ШассиЧерный шестигранный ключ Пластина
A) 5/16 дюйма (8 мм)
Сквозное отверстие
C) Пазы файла
B) Отверстие 1 дюйм (25 мм)
5/32 дюйма (4 мм)
Защелка
Болт Скос
Защелка
A) 2-1 / 8 дюйма (54 мм)
Сквозное отверстие
A) 5/16 дюйма (8 мм)
Сквозное отверстие
Используйте лицевую панель в качестве шаблона для паза.
Backset
Locator
См. Шаг 9 для регулировки толщины двери.
ACCO M01354-D Wireless Presenter Dongle Руководство пользователя с лазером
18 19
f ÅÈÓÐÖÕßËÎËÒÈÓÈÐÈÔÕËÒÓËÈÏÐÖáÃÐÕÈÐÐÖ
f ÅÈÎËÚËÕßÓÃÔÔÕÑÂÐËÈÏÈÉÇÖÑÄÑÓÖÇÑÅÃÐËÈÏËÒÓËÈÏ
f áÚËÕßÑÄÑÓÖÇÑÅÃÐËÈÍÓÑÊÈÕÍÈÍÑÕÑÓÃÂÐÈËÏÈÈÕÑÄÜËØÙÈÒÈÌÔÕÑÌÅÍÑÕÑÓÖáÅÍÎáÚÈÐÒÓËÈÏÐËÍ
f ÓÑÍÑÐÔÖÎßÕËÓÑÅÃÕßÔÂÖÇËÎÈÓÃËÎËÑÒÞÕÐÑÆÑ ÓÃÇËÑÕÈØÐËÍÃÇÎÂÒÑÎÖÚÈÐËÂÒÑÏÑÜË
¢ ÊÏÈÐÈÐËÂÐÈÑÇÑÄÓÈÐÐÞÈÍÑÏÒÃÐËÈÌ.HQVLQJWRQÏÑÆÖÕÎËÛËÕßÒÑÎßÊÑÅÃÕÈÎÂÅÑÊÏÑÉÐÑÔÕËËÔÒÑÎßÊÑÅÃÕßÇÃÐÐÑÈÖÅÑÅÔÑÑÕ
ÅÈÕÔÕÅËËÔÕÓÈÄÑÅÃÐËÂÏËÈÇÈÓÃÎßÐÑÌÑÏËÔÔËËÒÑÔÅÂÊËËÐÈÇÑÎÉÐÞÄÞÕßÅÞÒÑÎÐÈÐÞ
ÐÓÃÄÑÚÈÌÚÃÔÕÑÕÞÙ
ÉÈÐËÈÑÔÑÑÕÅÈÕÔÕÅËËÕÓÈÄÑÅÃÐËÂÏÈÇÈÓÃÎßÐÑÌÍÑÏËÔÔËËÒÑÔÓÈÇÔÕÅÃÏÔÅÂÊË) &&
ß ÒÓÃÅËÎ) && ÅÑÑÕÅÈÚÃÈÕÔÎÈÇÖáÜËÏÕÓÈÄÑÅÃÐËÂÏÃÐÐÑÈÖÔÕÓÑÌÔÕÅÑÐÈÇÑÎÉÐÑÔÑÊÇÃÅÃÕßÅÓÈÇÐÞØÒÑÏÈØËÇÃÐÐÑÈÖÔÕ
ÅÑÇÑÎÉÐÑÄÞÕßÖÔÕÑÌÚËÅÑÍÎáÄÞÏÒÑÏÈØÃÏÅÕÑÏÚËÔÎÈËÍÕÈÏÍÑÕÑÓÞÈÏÑÆÖÕÅÞÊÅÃÕßÔÄÑËÅÈÆÑÓÃÄÑÕÈÃÍÖÐÑÅ ÓÃÊÇÈÎÈÒÓÃ
ÅËÎ) && ÔÕÑÓÑÐÑÌÑÕÅÈÕÔÕÅÈÐÐÑÌÊÃÇÃÐÐÑÈÖÔÕÓÑÌÔÕÅÑÂÅÎÂÈÕÔÂ.HQVLQJWRQ & RPSXWHU3URGXFW * URXS) DVKLRQ, VODQG% OYGUG) ORRU6DQ0DWHR
& $ 86 $
ÉÈÐËÈÑÔÑÑÕÅÈÕÔÕÅËËÍÃÐÃÇÔÍÑÏÖÒÓÑÏÞÛÎÈÐÐÑÏÖÔÕÃÐÇÃÓÕÖ
ÐÐÑÈÖÔÕÓÑÌÔÕÅÑÒÓÑÕÈÔÕËÓÑÅÃÐÑËÔÑÑÕÅÈÕÔÕÅÖÈÕÑÆÓÃÐËÚÈÐËÂÏÖÍÃÊÃÐÐÞÏÅÔÕÃÐÇÃÓÕÈ566ÔÕÓÑÌÔÕÅÑÑÕÅÈÚÃÈÕ ÔÎÈÇÖáÜËÏÕÓÈÄÑÅÃÐËÂ
ÐÐÑÈÖÔÕÓÑÌÔÕÅÑÐÈÇÑÎÉÐÑÔÑÊÇÃÅÃÕßÒÑÏÈØËÇÃÐÐÑÈÖÅÑÇÑÎÉÐÑÄÞÕßÖÔÕÑÌÚËÅÑÍÎáÄÞÏÒÑÏÈØÃÏÅ ÕÑÏÚËÔÎÈËÍÕÈÏÍÑÕÑÓ
ÖÕÅÅÃÕßÔÄÑËÅÈÆÑÓÃÄÑÕÈ
ÐÐÑÈÙË × ÓÑÅÑÈÖÔÕÓÑÌÔÕÅÑÍÎÃÔÔÃ>% @ ÔÑÑÕÅÈÕÔÕÅÖÈÕÍÃÐÃÇÔÍÑÏÖ ÔÕÃÐÇÃÓÕÖ, & (6
ÉÈÐËÈÑÔÑÑÕÅÈÕÔÕÅËËÕÓÈÄÑÅÃÐËÂÏÅÓÑÒÈÌÔÍÑÆÑÔÑÅÈÕÃ
ÐËÂ.HQVLQJWRQÊÃÂÅÂÈÕÚÕÑÇÃÐÐÞÌÒÓÑÇÖÍÕÔÑÑÕÅÈÕÔÕÅÖÈÕÑÔÐÑÅÐÞÏÕÓÈÄÑÅÃÐËÂÏËÇÓÖÆËÏÔÑÑÕÅÈÕÔÕÅÖáÜËÏÒÑÎÑÉÈÐËÂÏÔÑÑÕÅÅÖáÜ
ØÇËÓÈÍÕËÅÅÓÑÒÈÌÔÍÑÆÑÔÑÅÈÕÃ
ÄËÖÚËÕßÍÑÒËáÊÃÅÎÈË ÅÈÕÔÕÅËËÇÃÐÐÑÆÑÒÓÑÇÖÍÕÃÇÎÂÅÓÑÒÞÒÈÓÈÌÇËÕÈÒÑÔÔÞÎÍÈv & RPSOLDQFH’RFXPHQWDWLRQwÐÃÔÃÌÕÈ
ZZZVXSSRUWNHQVLQJWRQFRP
Ð × ÑÓÏÃÙËÂÕÑÎßÍÑÇÎÂÆÑÔÖÇÃÓÔÕÅ ÐÑÅ
ÐÃÚÍËÖÍÃÊÞÅÃáÕÐÃÕÑÚÕÑÇÃÐÐÑÈËÊÇÈÎËÈÐÈÏÑÉÈÕÑÕÐÑÔËÕßÔÂÍÄÞÕÑÅÞÏÑÕØÑÇÃÏÄÈÔÒÈÚËÅÒÓÃÅËÎßÐÖáÖÕËÎËÊÃÙËáËÊÇÈÎËÂÅ
ÉÈÕÈÅÒÓÈÇÑÕÅÓÃÜÈÐËËÒÑÕÈÐÙËÃÎßÐÞØÐÈÆÃÕËÅÐÞØÒÑÔÎÈÇÔÕÅËÌÇÎÂÑÍÓÖÉÃáÜÈÌÔÓÈÇÞËÚÈÎÑÅÈÚÈÔÍÑÆÑÊÇÑÓÑÅßÂÍÑÕÑÓÞÈÏÑÆÖÕ
ÄÞÕßÅÞÊÅÃÐÞÐÈÒÓÃÅËÎßÐÑÌÈÆÑÖÕËÎËÊÃÙËÈÌÎÕËÎËÊÃÙËÈÌÎÖÚÈÐË ÄÑÎÈÈÒÑÇÓÑÄÐÑÌËÐ × ÑÓÏÃÙËËÑÄÖÕËÎËÊÃÙËËÇÃÐÐÑÆÑËÊÇÈÎËÂÒÑÉÃÎÖÌÔÕÃ
ÅÂÉËÕÈÔßÔÅÃÛËÏÏÈÔÕÐÞÏÆÑÓÑÇÔÍËÏÑ × ËÔÑÏÔÎÖÉÄÑÌÅÞÅÑÊÃÄÞÕÑÅÞØÑÕØÑÇÑÅËÎËÏÃÆÃÊËÐÑÏ ÅÍÑÕÑÓÑÏÅÞÍÖÒËÎËËÊÇÈÎËÈ
.HQVLQJWRQË $ && 2zÊÃÓÈÆËÔÕÓËÓÑÅÃÐÐÞÈÕÑÓÆÑÅÞÈÏÃÓÍË $ && 2ÔÈÇÓÖÆËÈÊÃÓÈÆËÔÕÓËÓÑÅÃÐÐÞÈËÐÈÊÃÓÈÆËÔÕÓËÓÑÅÃÐÐÞÈÕÑÓÆÑÅÞÈÏÃÓÍËÂÅÎÂá
ÂÔÑÄÔÕÅÈÐÐÑÔÕßáËØÔÑÑÕÅÈÕÔÕÅÖáÜËØÅÎÃÇÈÎßÙÈÅ
i .HQVLQJWRQ & RPSXWHU3URGXFWV * URXS × ËÎËÃÎ $ && 2% UDQGVáÄÑÈÐÈÔÃÐÍÙËÑÐËÓÑÅÃÐÐÑÈÕËÓÃÉËÓÑÅÃÐËÈÍÑÒËÓÑÅÃÐËÈËÎËËÐÑÈÅÑÔÒÓÑËÊÅ
ÐËÈÄÈÊÒËÔßÏÈÐÑÆÑ ÔÑÆÎÃÔËÂ.HQVLQJWRQ & RPSXWHU3URGXFWV * URXSÊÃÒÓÈÜÈÐÑÅÕÑÓÔÍËÈÒÓÃÅÃÊÃÜËÜÈÐÞ
Português
ATENÇÃO! INSTRUES DE SEGURANÇA ВАЖНЫЕ ИНСТРУМЕНТЫ
O Apresentador sem fios com Apontador Laser, включая apontador laser embedded.Assim, e à
semelhança de qualquer outro apontador laser, deve respeitar os avisos seguintes e as Instruções
de segurança, de modo a evitar lesões.
• Не имеет апонтадора лазера для остального пассивного лазера, но не фиксирует его для лазера. Se apontar o apontador para
os olhos de uma pessoa, ou se olhar fixamente para o raio laser, pode ter uma perda temporária de visão, que se manifesta no
visionamento de clears, brilho ofuscante ou desorientação.Tal situação é specificmente perigosa se a pessoa estiver a executar
uma actividade que exija muito da visão, como é o caso da Condução. Para além disso, расширенная экспозиция e / ou Repetida ao
raio laser pode ser perigosa e nociva para os olhos.
• Não aponte o apontador laser para qualquer parte do corpo de uma pessoa ou de um animal. Os apontadores laser são Concebidos
для большинства объектов inanimados. Em alguns estados e províncias, é ilegal apontar apontadores laser para agent de
autoridade ou para qualquer outra pessoa.O comprador / utilizador é Responsepel pela utilização do seu apontador laser em
concidade com a jurisdição local.
• Нет доступа к лазерному устройству для высочайшего качества. Os raios reflection podem ter o mesmo efeito dos raios apontados
directamente para os olhos.
• Os apontadores laser não são brinquedos. Não deixe um menor utilizar um apontador sem vigilância. Em alguns estados e
províncias, compra e / ou utilização de apontadores por um menor é ilegal.O comprador / utilizador é Response pela compra ou
utilização do seu apontador laser em contidade com a jurisdição local.
O Apresentador sem fios com Apontador Laser
• O рецептор tem uma luz indicadora com 2 ядра: verde e vermelho. Semper que carregar num botão do apontador, o LED do
Receiver pisca rapidamente na cor verde para indicar que a carga da pilha está num nível elevado. У вас есть carga da pilha estiver num
nível baixo, o LED passa a vermelho semper que carregar num botão.
• В качестве материала для заказа продукта Apresentador sem fios com Apontador Laser devem durar cerca de um ano. Tal previsão baseia-se num
uso médio do aparelho. Para aumentar a vida de uma pilha que não está a ser utilizada, desligue o remoto, интродукция или рецептор
no remoto.
Assistência técnica
A assistência técnica está disponível para todos os utilizadores dos produtos Kensington registados. Não se cobra pela assistência
técnica, excepto o custo da chamada telefónica e os valores de longa distância, se aplicáveis.Pode encontrar informações sobre como
contactar a assistência técnica na última página deste manual.
Техническая поддержка
• Pode encontrar a resposta para o seu problem na secção Perguntas mais fretes (FAQ) da area de assistência, нет веб-сайта
Kensington: www.support.kensington.com.
• Ligue de um telemóvel onde tenha acesso ao seu dispositivo
• Tenha por perto as seguintes informações:
– Nome, morada e número de phone
– O nome do produto Kensington –
provocouDeclaração de Segurança de Laser de Classe 2
За пользу, выберите etiqueta
autocolante de aviso de
segurança especí.
Этот лазер соответствует стандарту Norma Internacional EN / IEC 60825-1: 2007, Produtos Laser
Classe 2, установлен в соответствии с 21 CFR 1040.10 и 1040.1, заложен срок действия Aviso Sobre
Laser N.º 50, de 24 de Junho, 2007:
1. Emissão de um raio paralelo de luz visível;
2. Максимальная мощность 1 милливатт CW, интервал сравнения 650 ± 20 нанометров;
3. Температура при температуре от 0ºC до 40ºC
ATENÇÃO!
Este dispositivo não Possui Peças Passíveis de serem reparadas.A utilização de controlos ou
ajustes ou o desempenho de Procedimentos que não os especificados no presente documento
podem resultar numa exição perigosa à radiação.
DECLARAÇÃO SOBRE A INTERFERÊNCIA DE RADIOFREQUNCIA DA FCC (ФЕДЕРАЛЬНАЯ КОМИССИЯ ПО СВЯЗИ)
Примечание: Este dispositivo foi testado e está em concidade com os limites installedelecidos para de de la de la de la de la de 2000, 9000 Digitos FCC.Estes limites destinam-se a fornecer uma protecção адекватная contra interferências
prejudiciais em instalações residencenciais. Este equipamento gera, utiliza e pode emitir energia de radiofrequência e, se não for instalado
nem utilizado de acordo com as инструкций, pode provocar interferências nas comunicações de rádio. Нет Entanto, não existe nenhuma
garantia de que essa interferência não ocorra em definedadas instalações. У вас есть оборудование, вызывающее помехи на
приемника радио или телевидения, подача сигнала или переход:
• Переориентируйте или выберите локализацию антенны приема.
• Поместите дистанционный вход в оборудование и рецептор.
• Ligue o equipamento a uma tomada de um circuito diferente daquele a que o рецептор лигадо.
• Se Precisar de ajuda, consulte o agent da Kensington ou um técnico qualificado de rádio / televisão.
МОДИФИКАЦИИ: В соответствии с изменениями, которые не требуются для экспресс-доставки, Kensington может быть авторизован для использования.
является владельцем устройства, действующего в соответствии с правилами FCC, без каких-либо изменений.
Частотный интервал функционирования: 2,4 ГГц
ДЕКЛАРАЦИЯ СООТВЕТСТВИЯ С СООТВЕТСТВИЕМ С FCC
Этот продукт соответствует требованиям FCC. O funcionamento está sujeito às duas condições:
(1) este equipamento não pode causar interferências prejudiciais e (2) tem de suportar as interferências Recebidas, в том числе как que
Possam prejudicar o. Como Definido na Secção 2.909 dos regulamentos da FCC, a parte response por este dispositivo
é a Kensington Computer Products Group, 1500 Fashion Island Blvd, 3rd Floor, San Mateo, CA 94404, США.
DECLARAÇÃO DE CONFORMIDADE DA INDUSTRY CANADA
Это устройство соответствует требованиям, установленным в RSS-210. O seu funcionamento está dependente
das duas condições que seguem: (1) Este dispositivo não pode causar interferências e (2) tem de suportar as interferências Recebidas,
inclindo as que possible prejudicar o.
Цифровое оборудование класса [B] соответствует канадской норме ICES-003.
DECLARAÇÃO DE CONFORMIDADE DA CE
Kensington декларирует, что этот продукт соответствует требованиям, необходимым и не имеющим отношения к
Directivas da CE aplicáveis.
Para a Europa, pode obter uma cópia da Declaração de Conformidade para este produto, se clicar na ligação Declaração de
Conformidade, em www.support.kensington.com.
INFORMAÇÃO APENAS PARA OS ESTADOS-MEMBROS DA UE
A utilização do símbolo indica que este produto não pode ser tratado como lixo doméstico.Ao Certificar-se de que se desfaz
deste produto correctiveamente, ajudará a evitar Potenciais conquências negativas para o ambiente e para a saúde, que, de
outra forma, poderiam ocorrer pelo mau manuseamento deste produto. Para obter informação mais detalhada sobre a
reciclagem deste produto, contacte o município onde residence, os serviços de recolha de resíduos da sua área ou a loja onde
adquiriu o produto.
Kensington и ACCO são marcas registadas da ACCO Brands.Todas as outras marcas registadas e não registadas são propriedade dos
respectivos proprietários.
© Kensington Computer Products Group, 2016 г., подразделение ACCO Brands. Qualquer cópia, duplicação or воспроизводить não autorizada
dos context aqui presentes é proibida sem o consentimento escrito da Kensington Computer Products Group. Todos os direitos
reservados. 08/16
EC002NS Комплект кнопки пуска PKE Руководство пользователя 15_EC002-NS UserMan r1 Zhongshan Yihu Electronicsltd
3.Если я забуду установленный пароль, как его снова установить?
См. Инструкции по сбросу пароля на стр. 39-40 в руководстве пользователя.
4. Когда я пытаюсь завести машину, она заводится, но выключается через несколько секунд, в чем причина?
Возможны 4 причины:
A. Синий провод топливного насоса в 20-контактном жгуте проводов не подключается должным образом, что приводит к отсутствию электричества, когда
вы хотите запустить автомобиль. Провод топливного насоса рекомендуется подключать к проводу топливного насоса или к проводу масляного насоса
/ двигателю масляного тракта / всасывающему двигателю / в транспортном средстве, если вы не можете найти этот провод в транспортном средстве, вы можете подключить его с помощью
постоянный +12 провод или провод тач.
Б. Времени пуска недостаточно. Если да, то нужно продлить время старта. Для получения подробной инструкции,
, пожалуйста, обратитесь к времени запуска двигателя в настройке параметра времени продления в руководстве пользователя.
C.Проверьте, правильно ли подключен желтый провод стартера в 6-контактном жгуте.
D. Если в заводском брелоке вашего автомобиля есть иммобилайзер с чипом, вам следует добавить дополнительный модуль обхода, пожалуйста,
убедитесь, что байпас Модуль хорошо установлен и подключен, чтобы вы могли завести свой автомобиль.
5. Я не могу использовать функцию дистанционного запуска двигателя, в чем причина?
Пожалуйста, проверьте, правильно ли подключен синий провод топливного насоса в 20-контактном жгуте проводов.
По умолчанию установлен сигнал тревоги для автомобиля с автоматической коробкой передач, а переключатель 5 в модуле управления находится в нажатом положении. Если ваш автомобиль
является автомобилем с механической коробкой передач и вы хотите использовать функцию дистанционного запуска двигателя, нажмите переключатель 5 в выключенном состоянии
(это отсоедините жгут проводов зажигания 6P от модуля управления, затем нажмите переключатель
5, затем снова подключите жгут проводов зажигания 6P к модулю управления) и сначала установите ручное переключение передач.
После того, как комплект настроен на ручное переключение передач, водитель выходит из автомобиля и закрывает двери, не загораясь,
отойдите от автомобиля не менее чем на 8 метров, продолжайте нажимать кнопку запуска на пульте дистанционного управления для
около 3 секунд, чтобы выключить автомобиль. После этого пользователь может использовать функцию дистанционного запуска двигателя для автомобиля с механической коробкой передач
.
Ручной тормоз должен находиться в нейтральном положении при использовании функции дистанционного запуска автомобиля с механической коробкой передач.
Пользователь также может обратиться к информации о дистанционном запуске автомобиля с механической коробкой передач в руководстве пользователя.
6. Есть ли в устройстве функция защиты от взлома?
Да, настройка сигнализации по умолчанию – с функцией защиты от угона, а DIP-переключатель 4 в модуле управления – нажатие
вниз, если вам не нужна эта функция, нажмите DIP-переключатель 4 на модуле управления при выключенном питании.
статус.
7. DIP-переключатель 4 в модуле управления нажат, но я все еще не могу использовать функцию защиты от угона
, почему?
Проверьте:
A.Если вы нажмете DIP-переключатель 4 на модуле управления в выключенном состоянии (это сначала отсоедините провод зажигания 6P
от модуля управления, затем нажмите DIP-переключатель 4 на модуле управления, затем снова подключите провод зажигания 6P
жгут проводов с модулем управления. Если нет, исправьте настройку.
B. правильно ли подключен синий / черный провод пускового устройства двери, настройка по умолчанию отрицательная (-) и может быть переустановлена перемычкой
чтобы быть положительным (+), подробные инструкции см. в примечании 10 в руководстве пользователя.
8. Как заниматься программированием?
См. Метод программирования в руководстве пользователя.
9. Как подключить трехцветный (оранжевый, желтый и белый) провод GWR?
См. Примечание 8 в руководстве пользователя.
10. Как перевести устройство в режим камердинера?
См. Информацию о включении / выключении режима камердинера в руководстве пользователя.
11. Когда я открываю дверь, я слышал, как модуль управления иногда издает щелчок, но когда я закрываю дверь
, это нормально, это дефектный продукт?
№Когда вы открываете дверь автомобиля, реле в модуле управления должно срабатывать и в течение нескольких секунд издает щелчок
, и это нормально. И когда вы закроете дверь, щелчки прекратятся, так как реле
не в рабочем состоянии.
12. Как используется DIP-переключатель 6 в модуле управления?
Переключатель 6 предназначен для обратного тока. Для некоторых транспортных средств необходимо сначала повернуть ключ в положение ON, а затем педаль тормоза
будет с электричеством при наступлении на него.Для этого типа автомобилей необходимо нажать DIP-переключатель 6. Для
некоторых типов автомобилей Volkswagen (VW) даже все провода подключены правильно, но все еще не удается разблокировать двери автомобиля
при нажатии кнопки разблокировки, если Итак, отсоедините 6-контактный жгут проводов зажигания от модуля управления
, нажмите DIP-переключатель 6, затем снова подключите 6-контактный жгут проводов зажигания к модулю управления, теперь
все работает хорошо. Переключатель 6 в основном используется в некоторых автомобилях Volkswagen.
13.Есть ли провод GWR (масса во время работы)? Или как подключить провод GWR к EC002?
См. Примечание 8 относительно метода подключения проводов GWR.
14. Как подключить модуль доводчика окна к вашей сигнализации?
Если модуль доводчика окна необходимо соединить с сигнализацией EC002, соедините провод от модуля доводчика окна
с белым проводом запирания в проводе центрального замка двери.
15. Я включаю функцию PKE, а затем завожу автомобиль, но когда я останавливаюсь и выключаю двигатель, функция PKE
автоматически отключается, и мне нужно снова ее включить, в чем причина?
Проблема должна быть связана с тем, что синий / черный провод пускового механизма двери (-) плохо подключен.
Предупреждение: Этот продукт предназначен для установки только профессиональным установщиком автосигнализации! Любая попытка установки
данного продукта любым лицом, кроме обученного профессионального установщика автосигнализации, может привести к серьезным повреждениям
автомобиля или его компонентов.
16. Я подключаю провод пускового механизма двери, но при открытии двери автомобиля мигает указатель поворота. В чем причина
?
Если это так, сначала отсоедините провод пускового механизма двери, отсоедините жгут проводов зажигания 6P от модуля управления
, затем нажмите DIP-переключатель 2, снова подсоедините жгут проводов зажигания 6P к модулю управления.
17. Почему в моем комплекте нет оранжевого, белого и желтого провода?
Если сигнализация оснащена датчиком удара, то нет оранжевого, белого и желтого трехцветного провода GWR, и вы
можете использовать дополнительное реле SPST для создания провода GWR, подробные инструкции см. В примечании 9 в руководство пользователя
.
Информация о гарантии
Качество этого продукта строго контролируется перед отправкой с завода, что обеспечивает его улучшенные характеристики
при нормальном использовании.И мы предоставляем 1 год гарантии качества. Если произойдет сбой из-за качества продукции
, мы обеспечим бесплатный ремонт или замену. Любая неправильная работа / установка / использование кабеля
, в котором продукт не работал, не покрываются бесплатным ремонтом или заменой.
-44-
© EASYGUARD Electronics. Все права защищены.
-43- © EASYGUARD Electronics. Все права защищены.
SEC.gov | Превышен порог скорости запросов
Чтобы обеспечить равный доступ для всех пользователей, SEC оставляет за собой право ограничивать запросы, исходящие от необъявленных автоматизированных инструментов.Ваш запрос был идентифицирован как часть сети автоматизированных инструментов за пределами допустимой политики и будет обрабатываться до тех пор, пока не будут приняты меры по объявлению вашего трафика.
Пожалуйста, объявите свой трафик, обновив свой пользовательский агент, чтобы включить в него информацию о компании.
Чтобы узнать о передовых методах эффективной загрузки информации с SEC.gov, в том числе о последних документах EDGAR, посетите sec.gov/developer. Вы также можете подписаться на рассылку обновлений по электронной почте о программе открытых данных SEC, в том числе о передовых методах, которые делают загрузку данных более эффективной, и о SEC.gov, которые могут повлиять на процессы загрузки по сценарию. Для получения дополнительной информации обращайтесь по адресу [email protected].
Для получения дополнительной информации см. Политику конфиденциальности и безопасности веб-сайта SEC. Благодарим вас за интерес к Комиссии по ценным бумагам и биржам США.
Код ссылки: 0.5dfd733e.1631895545.809158f0
Дополнительная информация
Политика безопасности в Интернете
Используя этот сайт, вы соглашаетесь на мониторинг и аудит безопасности.В целях безопасности и обеспечения того, чтобы общедоступная услуга оставалась доступной для пользователей, эта правительственная компьютерная система использует программы для мониторинга сетевого трафика для выявления несанкционированных попыток загрузки или изменения информации или иного причинения ущерба, включая попытки отказать пользователям в обслуживании.
Несанкционированные попытки загрузить информацию и / или изменить информацию в любой части этого сайта строго запрещены и подлежат судебному преследованию в соответствии с Законом о компьютерном мошенничестве и злоупотреблениях 1986 года и Законом о защите национальной информационной инфраструктуры 1996 года (см. Раздел 18 U.S.C. §§ 1001 и 1030).
Чтобы обеспечить хорошую работу нашего веб-сайта для всех пользователей, SEC отслеживает частоту запросов на контент SEC.gov, чтобы гарантировать, что автоматический поиск не влияет на возможность доступа других лиц к контенту SEC.gov. Мы оставляем за собой право блокировать IP-адреса, которые отправляют чрезмерное количество запросов. Текущие правила ограничивают пользователей до 10 запросов в секунду, независимо от количества машин, используемых для отправки запросов.
Если пользователь или приложение отправляет более 10 запросов в секунду, дальнейшие запросы с IP-адреса (-ов) могут быть ограничены на короткий период.Как только количество запросов упадет ниже порогового значения на 10 минут, пользователь может возобновить доступ к контенту на SEC.gov. Эта практика SEC предназначена для ограничения чрезмерного автоматического поиска на SEC.gov и не предназначена и не ожидается, чтобы повлиять на людей, просматривающих веб-сайт SEC.gov.
Обратите внимание, что эта политика может измениться, поскольку SEC управляет SEC.gov, чтобы гарантировать, что веб-сайт работает эффективно и остается доступным для всех пользователей.
Примечание: Мы не предлагаем техническую поддержку для разработки или отладки процессов загрузки по сценарию.
Обувь для гольфа Nike TW15 – серебристый / серый / черный на InTheHoleGolf.com
Обувь для гольфа Nike TW15 – серебристый / серый / черный
Мужские кроссовки для гольфа Nike TW ’15 с верхом из материала Flyweave и технологией Dynamic Fit обеспечивают невероятно легкий, дышащий вид и адаптивную поддержку. Подошва в стиле Nike Free обеспечивает естественную гибкость, поэтому вы можете генерировать больше мощности при каждом взмахе.
«Nike продолжает отвечать на звонок каждый раз, когда я хочу раздвинуть границы», – сказал Вудс.«Новая технология Flyweave обеспечивает еще большую стабильность и поддержку моей стопы, и когда вы соедините ее с подошвой в стиле Free, я заметил, что могу лучше отталкиваться от земли и завершать свой замах с силой».
Nike Golf и Tiger Woods неустанно стремятся продвинуть вперед гольф. Вместе они способствуют развитию современного гольфа, продвигая инновационные продукты, превосходящие все ожидания.
Символом современного духа являются новые кроссовки TW ’15, новейшие фирменные кроссовки для гольфа Вудса и первые кроссовки Nike с революционной технологией Flyweave в верхней части.
TW ’15 – это последняя разработка Вудса и Тоби Хэтфилда, старшего директора Nike по спортивным инновациям. Вудс представил свой первый проект в качестве прототипа на WGC Bridgestone в 2011 году. С тех пор каждая итерация серии TW оснащалась подошвой в стиле Nike Free, постоянно повышая планку того, что спортсмены должны ожидать от обуви для гольфа.
Flyweave позволяет создавать цельный верх, сотканный для обеспечения отличной устойчивости к весу там, где это больше всего необходимо, исходя из требований или движений этого вида спорта.Flyweave не только прочный и поддерживающий, но и невероятно легкий и гибкий.
Nike дебютировала с технологией Flyweave в баскетбольной обуви в 2014 году. Несмотря на то, что динамические движения ног баскетболиста отличаются от движений игрока в гольф, оба требуют стабильности и естественного диапазона движений.
Благодаря легкой технологии Flyweave и специальной подошве в стиле Nike Free для гольфа дизайнеры смогли сделать TW ’15 на десять процентов легче, чем предыдущая модель.
Льготы- Конструкция Flyweave обеспечивает максимальную прочность и поддержку в виде легкого однослойного материала Технологии
- Dynamic Fit и Flywire обеспечивают легкость и стабильность, а также ограничивают производительность.
- Легкая межподошва Phylon для мягкой амортизации Подошва
- , вдохновленная Nike Free, обеспечивает свободу движений, позволяя максимально увеличить силу поворота
- Двухлетняя гарантия водонепроницаемости
Случай часто неправильно произносимых слов
Изучение языков с помощью компьютера
Fouz-González (Eds.), Исследование английского произношения: современные тенденции и
направлений (стр. 260–292). Бейзингсток, Великобритания / Нью-Йорк, Нью-Йорк: Palgrave Macmillan
Hincks, R. (2015). Технологии и обучение произношению. В M. Reed & J. Levis (Eds.),
Справочник по английскому произношению (стр. 505–519). Молден, штат Нью-Йорк: Wiley-
Blackwell.
Хинофотис Ф. и Бейли К. (1980). Реакция американских студентов на коммуникативные навыки
иностранных преподавателей.В J. Fisher, M. Clarke, & J.
Schacter (Eds.), On TESOL ’80: Building bridges (стр. 120–133). Вашингтон, округ Колумбия:
TESOL.
Хуальде, Дж. И. (2014). Los sonidos del español. Кембридж, Великобритания: Cambridge University
Press.
Йоуп, Г. (1995). Оценка потребности в улучшении ввода в посткритический период
овладение языком. В D. Singleton & Z. Lengyel (Eds.), Возрастной фактор во втором освоении языка
(стр.95–123). Клеведон, Великобритания: Multilingual Matters
Джеймисон Дж., Шапель К. и Прейсс С. (2005). ЗВОНИТЕ оценка разработчиками, преподавателем
,и студентами. CALICO Journal, 23 (1), 93–125.
Ява, А., Сонг, X., Финин, Т., и Ценг, Б. (2007). Почему мы Twitter: Понимание использования микроблогов
и сообществ. Труды совместного 9-го семинара WEBKDD &
1stSNA-KDD 2007 г. (стр. 56–65). Сан-Хосе, Калифорния. Получено с
http: // ebiquity.umbc.edu/_file_directory_/papers/369.pdf
Джонс, Д., Роуч, П., Сеттер, Дж., и Эслинг, Дж. (2011). Кембриджский английский словарь с произношением
(18-е изд). Кембридж, Великобритания: Издательство Кембриджского университета.
Юнко, Р., Елавский, М. К., и Хейбергер, Г. (2013). Тестирование твиттера: оценка результатов
для сотрудничества, взаимодействия и успеха студентов. Британский журнал
Образовательные технологии, 44 (2), 273–287.
Юнко, Р., Хейбергерт, Г., & Локен, Э. (2011). Влияние Twitter на вовлеченность и оценки студентов колледжа
. Журнал компьютерного обучения, 27, 119–132.
Кассенс-Нур, Э. (2012). Твиттер как учебная практика для повышения активного и неформального обучения
в высшем образовании: пример устойчивых твитов. Активное обучение в
Высшем образовании, 13 (1), 9–21.
Кеннеди К. и Леви М. (2008). L’italiano al telefonino: Использование SMS для поддержки изучения языка
начинающих.ЗВОНИТЕ, 20 (3), 315–330.
Кеннинг, М-М. (2007). ИКТ и изучение языков. От печати до мобильного телефона.
Бейзингстоук, Великобритания / Нью-Йорк, Нью-Йорк: Пэлгрейв Макмиллан.
Крашен С. (1985). Исходная гипотеза: проблемы и последствия. Оксфорд, Великобритания:
Pergamon Press.
Лами, М.-Н., и Зоуру, К. (2013). (Ред.). Социальные сети для языкового образования.
Бейзингсток, Великобритания: Пэлгрейв Макмиллан.
Ли, Дж., Джанг Дж. И Плонски Л. (2015). Эффективность второго языка
Инструкция по произношению: метаанализ. Прикладная лингвистика, 36 (3), 345–366.
Левис Дж. (2007). Компьютерные технологии в обучении и исследовании произношения. Ежегодный
Обзор прикладной лингвистики, 27, 184–202.
Леви М. (1997). Теоретически ориентированный CALL и процесс разработки. Компьютерная
Изучение языков, 10 (1), 41–56.
Леви, М., И Кеннеди, C. (2005). Изучение итальянского с помощью мобильного SMS. В A. Kukulska-
Hulme & J. Traxler (ред.), Мобильное обучение: пособие для преподавателей и инструкторов.
Лондон, Великобритания: Тейлор и Фрэнсис.
Лин, С.-Х., Варшауэр, М., и Блейк, Р. (2016). Изучение языка через социальные сети
: Восприятие и реальность. Language Learning & Technology, 20 (1), 124–
147. Получено с http://llt.msu.edu/issues/feb February2016/linwarschauerblake.pdf
Молекулярный скрининг детерминант вирулентности энтерококков и потенциала генетического обмена между пищевыми продуктами и медицинскими изолятами
Резюме
Энтерококки используются в качестве заквасок и пробиотических культур в пищевых продуктах и являются естественными загрязнителями пищевых продуктов. Род Enterococcus приобретает все большее значение как причина внутрибольничных инфекций, и эта тенденция усугубляется развитием устойчивости к антибиотикам. В этом исследовании мы исследовали распространенность известных детерминант вирулентности в заквасочных, пищевых и медицинских штаммах Enterococcus faecalis , E.faecium и E. durans . Стратегии ПЦР и генных зондов были использованы для скрининга изолятов энтерококков как из пищевых, так и из медицинских источников. Для штаммов E. faecalis и E. faecium были обнаружены различные и отчетливые образцы встречаемости детерминант вирулентности. Медицинский Штаммы E. faecalis имели больше детерминант вирулентности, чем пищевые штаммы, которые, в свою очередь, имели больше, чем стартовые штаммы. Все протестированные штаммы E. faecalis () обладали множественными детерминантами (от 6 до 11). Штаммы E. faecium в целом не имели детерминант вирулентности, за заметными исключениями. Примечательно, что детерминанты esp и gelE были идентифицированы в медицинских штаммах E. faecium . Эти детерминанты вирулентности ранее не были идентифицированы у штаммов E. faecium и могут быть результатом региональных различий или эволюции патогенного E. faecium . Фенотипическое тестирование выявило существование явно молчаливых генов gelE и cyl .У E. faecalis тенденция этих молчащих генов отражает тенденцию экспрессируемых детерминант. Была исследована способность стартовых штаммов приобретать детерминанты вирулентности за счет естественных механизмов конъюгации. Была продемонстрирована трансконъюгация, при которой стартовые штаммы приобретали дополнительные детерминанты вирулентности от медицинских штаммов. Кроме того, несколько генов, кодирующих феромоны, были идентифицированы как в пищевых, так и в исходных штаммах, что указывает на их способность приобретать плазмиды с другими половыми феромонами.Эти результаты предполагают, что использование Enterococcus spp. в пищевых продуктах требует тщательной оценки безопасности.
Расширяется диапазон бактериальных культур, используемых в пищевых продуктах, и повышается интерес к пробиотикам и их пользе для здоровья. Основной проблемой является безопасность культур, которые можно употреблять в пищу живыми и в больших количествах. Для этой цели используют несколько видов молочнокислых бактерий (1, 15), из которых наиболее противоречивыми являются энтерококки. Энтерококки используются в качестве пробиотиков для улучшения микробного баланса кишечника и для лечения гастроэнтерита у людей и животных (2, 27).Они входят в состав естественной флоры некоторых продуктов и могут оказывать благотворное влияние. В некоторых сырах они важны для созревания и развития вкуса (3, 41, 42). Некоторые энтерококки также продуцируют бактериоцины, которые могут иметь активность против Listeria (17). Энтерококки встречаются в качестве загрязнителей пищевых продуктов и могут быть причиной болезней пищевого происхождения. Влияние энтерококков на безопасность пищевых продуктов недавно было изучено (13).
Род Enterococcus имеет особое медицинское значение из-за его повышенной заболеваемости как причины заболевания, особенно при нозокомиальных инфекциях, а также из-за того, что доступная антибиотикотерапия скомпрометирована из-за развития устойчивости к антибиотикам (22, 37).Энтерококковые культуры использовались в молочных ферментациях на протяжении десятилетий, а изоляты с историей безопасного использования продвигаются как пробиотические культуры. Эти бактерии, вероятно, представляют наибольший риск для здоровья человека из всех видов, используемых в настоящее время таким образом.
Различия между энтерококковым патогеном и явно безопасным штаммом для употребления в пищу неясны, а способность последнего приобретать факторы вирулентности путем переноса гена не исследовалась. Уже установлено, что молекулярная систематика энтерококков не приводит к различию между этими двумя типами штаммов.Наши знания о вирулентности энтерококков неполны, отчасти из-за того, что они являются нормальными человеческими комменсалами и, как таковые, обладают тонкими чертами вирулентности, которые нелегко идентифицировать. Признаки вирулентности включают прилипание к ткани хозяина, инвазию и образование абсцесса, модуляцию воспалительных реакций хозяина и секрецию токсичных продуктов. Было охарактеризовано несколько генов факторов вирулентности в Enterococcus faecalis (см. Таблицу), и их эффекты были продемонстрированы на животных моделях (4, 19, 21, 23, 34) и культивируемых клетках (25, 31).В этом исследовании изучали распространенность известных факторов вирулентности в штаммах медицинских, пищевых и молочных заквасок Enterococcus .
ТАБЛИЦА 2
Enterococcus Выявленные факторы вирулентности
| Ген (ы) | Роль продукта в вирулентности | Ссылка (и) |
|---|---|---|
| агрегация белка 905 905 к эукариотическим клеткам; агрегация и конъюгация клеток | 14, 44 | |
| gelE | Токсин; внеклеточная металлоэндопептидаза, гидролизует желатин, коллаген, гемоглобин и другие биоактивные соединения | 39 |
| CylL v CylL s | Предшественник цитолизина (гемолизин-бактериоцин); экспрессия cylL v -L s , -M, -B и – A необходим для продукции активного цитолизина, который лизирует широкий спектр эукариотических и грамположительных клеток | 16 |
| CylM | Посттрансляционная модификация цитолизина | 16 |
| CylB | Транспорт цитолизина | 16 |
| 16 | ||
| CylA | ESP 908Белок, связанный с клеточной стенкой, участвует в уклонении от иммунитета; может быть ассоциирован с Cyl генами на острове патогенности | 35, 36 |
| efaAfs; efaAfm | Адгезины клеточной стенки, экспрессируемые в сыворотке E.faecalis и E. faecium , соответственно | 29, 38 |
| cpd, cob, ccf, cad | Половые феромоны, хемотаксические для лейкоцитов человека; облегчают конъюгацию | 7, 11 |
Энтерококки известны своей способностью обмениваться генетической информацией путем конъюгации (5), и эти процессы, как известно, происходят в желудочно-кишечном тракте (19). Известно, что наряду с трансмиссивными плазмидами устойчивости к антибиотикам, факторы вирулентности, такие как продукция гемолизина-цитолизина и способность к адгезии, передаются посредством высокоэффективных механизмов переноса генов (4, 16, 25, 43).Таким образом, это исследование также изучало возможность того, что очевидно безопасные заквасочные или пробиотические культуры могут приобретать факторы вирулентности путем конъюгации. Кроме того, возможность конъюгации была исследована путем скрининга на присутствие недавно идентифицированных генов, кодирующих феромоны, продукты которых участвуют в инициировании описанных процессов конъюгации (7; Институт геномных исследований База данных генома E. faecalis [http : //www.tigr.org/tdb/mdb/mdb.hmtl]). Полагают, что половые феромоны также участвуют в возникновении воспалительной реакции.Эти феромоны обладают хемотаксическим действием для полиморфно-ядерных лейкоцитов человека и крысы in vitro и вызывают выработку супероксида и секрецию лизосомальных ферментов (11, 24, 33). Поэтому их можно рассматривать как факторы вирулентности.
МАТЕРИАЛЫ И МЕТОДЫ
Штаммы бактерий и среды.
Escherichia coli JM109 был получен от Promega. L-бульон (26) использовали для роста E. coli при 37 ° C и при необходимости добавляли ампициллин в количестве 100 мкг / мл -1 .Штаммы энтерококков и плазмиды, использованные в этом исследовании, перечислены в таблице. Штаммы энтерококков выращивали в бульоне для инфузии мозга и сердца (BHI) и агаре (Oxoid) при 37 ° C. Для экспериментов по спариванию энтерококков в среду добавляли эритромицин в количестве 50 мкг / мл -1 , канамицин в количестве 500 мкг мл -1 , стрептомицин в количестве 1000 мкг мл -1 , тетрациклин в количестве 10 мкг мл -1 , рифампицин в количестве 25 мкг / мл -1 и фузидовая кислота в количестве 25 мкг / мл -1 , когда это необходимо.
ТАБЛИЦА 1
Штаммы энтерококков и плазмиды, использованные в этом исследовании
| Штамм или плазмида | Соответствующий маркер (-ы) a | Комментарий и / или ссылка | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OG1s | str gelE + | 20 | |||||||||||||||||
| DS16 | tet cylMBA ag + tet cylMBA ag + ; 40 | ||||||||||||||||||
| FA2-2 | rif fus | реципиентный штамм без плазмид, используемый в экспериментах по спариванию; 9 | |||||||||||||||||
| JA2-2 | rif fus | Реципиентный штамм без плазмид, используемый в качестве положительного контрольного штамма для детерминант половых феромонов | |||||||||||||||||
| EBh2 | efaAfs | 909 Ec2594esp + | Медицинский изолят из крови (Оксфорд) | ||||||||||||||||
| FI9575 | винтовочный цилиндр MBA + ag +| FI9530 | RIFFUS Ery | Трансконъюгантный стартовый штамм; это исследование | FI9574 | тетри цилиндр MBA + agg + | трансконъюгантный стартовый штамм; это исследование | Плазмиды | pAD1 | Hem-Bac | Конъюгативная плазмида из E.faecalis DS16 | pAD2 | ery str kan | Плазмида из E. faecalis DS16; Tn 917 несет детерминант | pAM714 | ery Hem-Bac | pAD1 :: Tn 917 перенос дикого типа; 20 | |
Изоляты энтерококков.
Изоляты Enterococcus были собраны из национальных коллекций и различных компаний и учебных заведений, включая Американскую коллекцию типовых культур, Deutsche Sammlung von Mikroorganismen, Национальные коллекции промышленных и морских бактерий, CIT INIA Испания, Датскую ветеринарную лабораторию, Университет Бата, Мичиганский университет и Лаборатории общественного здравоохранения, Коллиндейл.Штаммы идентифицировали до уровня вида с помощью наборов API ID32-STREP (bioMerieux Vitek Inc., Hazelwood, MO). Один заквасочный штамм (F41) был выделен из датского кисломолочного продукта путем разбавления культуры четвертью раствора Рингера и посева разведений на агар BHI. Затем изолированные колонии подвергали анализу последовательности 16S рРНК и биохимическому тестированию. Результаты показали, что это был штамм E. faecalis . Для тестирования использовались 49 штаммов, которые составили панель А.Панель A состояла из 9 молочных заквасок, 22 пищевых продуктов и 18 медицинских штаммов, представляющих различные группы источников (молоко, сыр, мясо, кровь, гной, моча, фекалии, окружающая среда в больнице).
Контрольными штаммами, использованными в экспериментах ПЦР и для создания зонда, были E. faecalis, штаммы EBh2 ( efaAfs + ), OG1S ( gelE + ) и DS16 ( cylM + 909 + cylA + agg + ) и E.faecium Ec2594 ( esp + efaAfm + ) (Таблицы и).
Выделение энтерококковой ДНК.
Тотальную ДНК экстрагировали из культур Enterococcus в течение ночи по методу Lewington et al. (28).
Манипуляции с ДНК.
Манипуляции с ДНК, включая расщепление рестрикционной эндонуклеазой и Саузерн-блоттинг, проводили стандартными методами (31). Саузерн-гибридизацию проводили с использованием системы прямого мечения и обнаружения нуклеиновых кислот ECL (Amersham) в условиях стандартной жесткости, за исключением того, что 0.В буфере для гибридизации использовали 25 М NaCl. Для гибридизации по Саузерну с использованием зонда efaA использовались условия высокой жесткости с первичным промывочным буфером, содержащим 0,1 × SSC (1 × SSC – это 0,15 М NaCl плюс 0,015 М цитрат натрия). Рестрикционные ферменты, 5-бром-4-хлор-3-индолил-β-d-галактопиранозид (X-Gal) и изопропил-β-d-тиогалактопиранозид (IPTG) были приобретены у Promega и использовались в соответствии с инструкциями производителя. Плазмидную ДНК из E. coli , используемую для анализа последовательности, очищали с использованием набора QIAprep spin miniprep (QIAGEN).Секвенирование выполняли с помощью автоматического секвенатора Applied Biosystems 373A и набора для секвенирования цикла BigDye Terminator (PE Applied Biosystems) в соответствии с инструкциями производителя.
Олигонуклеотидные праймеры для ПЦР получали с использованием синтезатора ДНК / РНК Applied Biosystems 392 (таблица). ПЦР-амплификации проводили в реакционных смесях объемом 50 мкл с использованием 5 мкг ДНК, 15 мМ MgCl 2 , 20 пмоль каждого праймера и 1 ед. ДНК-полимеразы AmpliTaq (Perkin-Elmer Corp., Фостер-Сити, Калифорния). Образцы покрывали 40 мкл минерального масла и подвергали начальному циклу денатурации (94 ° C в течение 2 минут), отжигу (при соответствующей температуре в течение 2 минут; таблица) и элонгации (72 ° C в течение 2 минут). с последующими 29 циклами денатурации (92 ° C в течение 15 с), отжига (при соответствующей температуре в течение 15 с) и удлинения (72 ° C в течение 15 с). Фрагменты, полученные с помощью ПЦР, очищали с использованием набора для очистки QIAquickPCR (QIAGEN) перед клонированием в векторы pGEM-T с использованием системы pGEM-T Easy Vector (Promega) или мечения зонда.Вырожденные праймеры были предоставлены Sigma-Genosys (Кембридж, Англия) и использовались с системами ECL 3′-олигомечени и детекции (Amersham). Тотальную геномную ДНК выделяли из штаммов энтерококков, и все образцы неоднократно подвергали скринингу как с помощью ПЦР, так и гибридизации по Саузерну с использованием праймеров и зондов, описанных выше. Согласованные данные о наличии целевых генов вирулентности представлены в таблице.
ТАБЛИЦА 3
Праймеры и продукты ПЦР для определения детерминант вирулентности
| Ген и праймер | Последовательность (от 5 ‘до 3’) | Размер продукта (п.о.) |
|---|---|---|
| ТЕ3 | AAGAAAAAGAAGTAGACCAAC | одна тысяча пятьсот пятьдесят-три |
| ПЭ4 | AAACGGCAAGACAAGTAAATA | |
| Gele | ||
| ТЕ9 | ACCCCGTATCATTGGTTT | 419 |
| ТЕ10 | ACGCATTGCTTTTCCATC | |
| cylM | ||
| ТЕ13 CTGATGGAAAGAAGATAGTAT | 742 | |
| ТЕ14 TGAGTTGGTCTGATTACATTT | ||
| cylB 905 92 | ||
| ТЕ15 | ATTCCTACCTATGTTCTGTTA | 843 |
| ТЕ16 | AATAAACTCTTCTTTTCCAAC | |
| cylA | ||
| ТЕ17 | TGGATGATAGTGATAGGAAGT | 517 |
| TE18 | TCTACAGTAAATCTTTCGTCA | |
| ЭСП | ||
| TE34 | TTGCTAATGCTAGTCCACGACC | 933 |
| TE36 | GCGTCAACACTTGCATTGCCGAA | |
| efaAfs | ||
| TE5 | GACAGACCCTCACGAATA | 705 |
| TE6 | AGTTCATCATGCTGTAGTA | |
efaAfm| | | |
| TE37 | AACAGATCCGCATGAATA | 735 |
| TE38 | CATTTCATCATCTGATAGTA | |
| CPD | ||
| TE51 | TGGTGGGTTATTTTTCAATTC | 782 |
| TE52 | TACGGCTCTGGCTTACTA | |
| початка | ||
| TE49 | AACATTCAGCAAACAAAGC | +1405 |
| TE50 | TTGTCATAAAGAGTGGTCAT | |
| CCF | ||
| TE53 | GGGAATTGAGTAGTGAAGAAG | 543 |
| TE54 | AGCCGCTAAAATCGGTAAAAT | |
ТАБЛИЦА 4
Резюме результатов ПЦР, Саузерн-гибридизации и фенотипического скрининга для изолятов панели A
| Штамм | Виды | Источник (-и) фенотип | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Пускатели | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F12 | E.faecium | Закваска для сыра, сухое молоко | efaAfm + | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F13 | E. faecium | efaAfm efaAfm сухое молоко||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F20 | E. faecium | Стартер для товарных молочных продуктов | efaAfm + | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F21 | E.faecium | Стартер для товарных молочных продуктов | efaAfm + | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F22 | E. faecium | Испанский колбасный стартер | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| E. faecium | Стартер испанской колбасы (включая домашние) | efaAfm + | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F25 | E.durans | Закваска для сыра Чеддер | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F27 | E. faecalis | Испанская закваска для сыра Манчего | efaAfs 9022 9059 909 909 cagg9 cob + ccf + cad + | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F41 | E. faecalis | Стартер для коммерческого молочного продукта efacalis | agg + cpd + cob + ccf + cad + | GelE – | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F1 | E.faecalis | Простокваша | efaAfs + gelE + ag + cylMBA + cpd + cpd + cpd + cpd cad + | GelE – Hyl + | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F2 | E. faecalis | Датский голубой сыр | efaAfs 905 + cpd + cob + ccf + cad + | Hyl + | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| E.faecalis | Датский сыр с плесенью | efaAfs + agg + cylMBA + cpd + cpd + cb92 909 | Hyl + | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F4 | E. faecalis | Сыр Стилтон | efaAfs + gelE Aggg esp + cpd + cob + ccf + cad + | GelE – Hyl | 3 | 9faecalis | Молоко | efaAfs + gelE + esp + cpd + cpd + cob + cob | GelE + | | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F6 | E. faecalis | Молочный сыр | gelE + agg + cp92 909 + ccf + cad + | GelE + | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F7 | E.faecalis | Сырое молоко | efaAfs + gelE + cpd + cob + ccf + ccf + ccf | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F8 | E. faecalis | Сырое молоко | efaAfs + gelE + agg + cp + cad + | GelE + | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F9 | E.faecium | Сыр | efaAfm + ( efaAfs + ) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F10 | E. faec3| | F11 | E. faecium | Ферментированные пищевые отходы | efaAfm + ( efaAfs + ) | 3 | E.faecium | Коммерческое молоко | efaAfm + ( efaAfs + ) | F15 | E. | F16 | E. faecium | Сыр Чеддер | efaAfm + ( efaAfs + ) | 3 | E.faecium | Ветчина консервированная | efaAfm + ( efaAfs + ) | F19 | E. efaAfs + ) | F24 | E. durans | Сыр чеддер | efaAfm + ( | 9026F234 Молоко пастеризованное | efaAfs + | F28 | E. faecium | Розничный бройлер | F31 | E. faecium | Розничный бройлер | efaAfm + ( efaAfs + ( efaAfs | 3 | 3 | 3 | 3 | 3 | 33 | 3 | 3 | 3 | 3 | 9faecalis | Свинина для розничной торговли | efaAfm + efaAfs + gelE + cpd + 9059add 909 909 909 902 909 909 909 909 902 905 | GelE + | Медицинский | P1 | E. faecalis | 902 905 905 905 905 905 905 905 905 905 905 gelE + agg + cpd + cob + ccf + cad + | GelE + 3 | E.faecalis | Человеческий рот ( gelE + контроль) | efaAfs + gelE + cpd + cpd + cb22 909 + | GelE + | P4 | E. faecalis | Эндокардит ( cylMBA + ag 2 + 90fa92 + 9092 90fa92 + agg + cylMBA + cpd + cob + ccf + cad + | GelE + Hyl + | E.faecium Blood (Oxford) ( esp + efaAfm + контроль) | efaAfm + ( efaAfs + ( efaAfs + ( efaAfs ) 909 909 909 909 909 909 909 909 909 909 909 909 909 909 909 909 909 9 | P13 | E. faecium | Pus (Yeovil) | efaAfm + ( efaAfs + ) | E.faecalis | Кровь (Ньюкасл) | efaAfs + gelE + agg + cylMBA + | cf22 909 + cad + GelE + > GelE – Hyl + | P20 | E. faecium | промежности efaAfs + ) gelE + esp + | GelE + > GelE – | P21 | E.faecalis | Тампон из раны (больница Сент-Хелиер) | efaAfs + gelE + agg + cylMB + куб. См + agg + cylMBA + esp + cpd + cob + ccf 9022 905 – Hyl + | P31 | E.faecalis | Моча (NE Lincs.) | efaAfs + gelE + ag + esp + | 2 cpd 9023 9023 + cad + GelE – | P36 | E. faecalis | Кровь (Плимут 9023 9023 9 905 905 909 905 902 905 EFAAfs + cylMBA + esp + cpd + cob + ccf + cad3 9023 909 9023 9023 9023 9023 9023 909 9023 9023 9023 909 9023 9023 9023 9023 | P41 | E.faecalis | Кровь, эндокардит (Солсбери) | efaAfs + esp + cpd + cob | 9 | P46 | E. faecium | Рневной тампон (Кардифф) | efaAfm + ( efaAfs + 59 | | E.faecium | Кал (борона) | efaAfm + ( efaAfs + ) esp + | E. | efaAfm + ( efaAfs + ) esp + | P61 | E. ( efaAfs + ) esp + | P66 | E.faecium | Кровь, эндокардит (Dyfed) | efaAfm + ( efaAfs + ) esp + | + efaAfm + ( efaAfs + ) esp + | |
Создание праймеров и зондов.
Праймеры для ПЦРдля генов вирулентности efaA, gelE, Cyl и agg. , описанные в таблице, были разработаны на основе опубликованных последовательностей и последовательностей базы данных EMBL.Праймеры для ПЦР для esp были первоначально разработаны на основе последовательностей праймеров, предоставленных В. Шанкаром (Университет Оклахомы), а праймеры для детерминант половых феромонов были основаны на последовательностях в базе данных генома E. faecalis в Институте геномики. Исследовать. Вырожденный праймер был разработан для детерминанты полового феромона cAD1 на основе аминокислотной последовательности и смещения энтерококковых кодонов. Пакет программного обеспечения для анализа последовательностей Wisconsin Genetics Computer Group, версия 8, использовался для анализа последовательностей и сравнений.Было выполнено выравнивание множественных последовательностей с использованием плазмид половых феромонов pAD1, pPD1 и pCF10, и области с высокой консервативностью были использованы для создания праймеров для вещества агрегации ( agg ). Штамм P4, содержащий pAD1, использовали для создания зонда, специфичного для agg . Не было получено подходящих праймеров для структурных генов цитолизина ( cylL L и cylL S ) из-за значительной гомологии последовательностей между двумя генами. Все фрагменты, полученные с помощью ПЦР, клонировали в векторы pGEM-T и секвенировали для подтверждения идентичности (данные не показаны).Фрагменты целевой ДНК, амплифицированные с помощью ПЦР, служили ДНК-зондами для экспериментов по Саузерн-гибридизации.
Производство желатиназы и гемолизина.
Продукция желатиназы определялась на агаре Тодда-Хьюитта, содержащем 30 г желатина (Difco) на литр. Отдельные колонии наносили штрихами на чашки, выращивали в течение ночи при 37 ° C и помещали при 4 ° C на 5 ч перед исследованием зон помутнения вокруг колоний, указывающих на гидролиз. Для исследования продукции гемолизина штаммы наносили штрихами на чашки с многослойным агаром со свежей лошадиной кровью и выращивали в течение 1-2 дней при 37 ° C.Зоны просветления вокруг колоний указывали на продукцию гемолизина (таблица).
Генерация трансконъюгантов.
Скопления бульонов проводили по методу Clewell et al. (8) с использованием ночных культур и ночных вязок. Сопряжения фильтров проводили с использованием смеси донор-реципиент 1: 1. Вкратце, 0,5 мл каждой ночной культуры смешивали и собирали с использованием фильтров с размером пор 0,45 мкм и инкубировали на чашках с агаром BHI при 37 ° C в течение ночи. Клетки собирали, разводили в бульоне BHI и выращивали в течение 2 дней при 37 ° C на чашках с агаром BHI.
Для отбора трансконъюгантов необходимо было определить подходящие фенотипы как для трансмиссивной плазмиды, так и для штаммов-реципиентов. E. faecalis DS16 плазмида pAD2 несет транспозон устойчивости к эритромицину (Tn 917 ), который, как известно, с высокой частотой транспонируется на плазмиду pAD1, и этот феномен был использован для обеспечения положительной селекции на присутствие pAD1 на основе эритромицина. сопротивление. E. faecalis F41 оказался естественно устойчивым к рифампицину и фузидовой кислоте, и это обеспечило контрселективные маркеры для прямого отбора переноса pAD1 из E.faecalis DS16. Сопряжение фильтров использовали для получения трансконъюгантов (таблица).
Противоселективные фенотипы не были идентифицированы в других исходных штаммах, и, таким образом, pAD1 был перенесен с использованием скрещивания в бульоне в новый фоновый штамм, основанный на лабораторном штамме E. faecalis FA2-2. Это предоставило новый штамм {“type”: “entrez-nucleotide”, “attrs”: {“text”: “F19530”, “term_id”: “4825810”}} F19530, чувствительный к тетрациклину и несущий передаваемый плазмида вирулентности pAD1, помеченная геном устойчивости к эритромицину Tn 917 .Из этого донорского штамма можно было ввести pAD1 в E. faecalis F27 путем спаривания на фильтре.
E. faecalis FI9530 использовали непосредственно в вязках со стартовым штаммом E. faecium F12 с использованием отбора lac Z. E. faecalis FI9530 имеет белый цвет на чашках с агаром X-Gal – BHI, тогда как стартовые штаммы E. faecium имеют синий цвет.
РЕЗУЛЬТАТЫ
Три разные группы изолятов энтерококков были проверены на наличие охарактеризованных детерминант вирулентности.В эти группы были намеренно добавлены стартовые штаммы, штаммы, встречающиеся в естественных условиях в пище, и штаммы, выделенные от клинических инфекций человека.
Распределение детерминант вирулентности в пищевых продуктах и медицинских изолятах.
ПЦР-амплификации и Саузерн-гибридизации энтерококковой ДНК со специфическими праймерами и зондами для каждого описанного гена (таблицы и) выявили четкие тенденции в появлении детерминант вирулентности в пищевых и медицинских штаммах. Краткое изложение характера встречаемости генов вирулентности у E.faecalis и E. faecium в трех группах представлены в табл.
ТАБЛИЦА 5
Процентная частота детерминант вирулентности efaA ( efaAfs и / или efaAfm ), gelE, agg, cylM, cylB, cylA, esp, cpd, cob, ccf92 905 cad среди пищевых и медицинских изолятов E. faecalis и E. faecium
| Категория и организм | % Заболеваемость детерминантом a | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| efaA | гель aggцилиндр | цилиндр | цилиндр | esp | cpd b | 9080 9080 | ||||||||||
| E.faecalis | 100 | 50 (0) | 100 | 0 (0) | 0 | 0 | 0 | 100 | ||||||||
| E. faecium | 100 | 0 (0 0) | 0 | 0 (0) | 0 | 0 | 0 | 0 | ||||||||
| Продукты питания | 1 | E.faecalis89 | 78 (56) | 67 | 44 (44) | 44 | 44 | 33 | 100 | |||||||
| E. faecium | 82 | 0 ( | 0 | 0 (0) | 0 | 0 | 0 | 0 | ||||||||
| Медицинское | 1 | E.faecalis100 | 89 (56–33) | 78 | 56 (33) | 56 | 44 | 44 | 100 | |||||||
| E. faecium | 100 ( | 11) 0) | 0 | 0 (0) | 0 | 0 | 78 | 0 | ||||||||
(i)
E. faecalis .Штаммы E. faecalis обладают значительно большим количеством детерминант вирулентности, чем E.faecium , и внутри этих трех групп наблюдается четкая тенденция. Все из протестированных штаммов E. faecalis , включая два стартовых штамма, обладали множественными детерминантами вирулентности (от 6 до 11). Три штамма E. faecalis (два медицинских и один пищевой) обладали всеми генами вирулентности. Детерминанты половых феромонов были обнаружены исключительно у штамма E. faecalis . Они встречались во всех протестированных штаммах, иногда с определителем agg , а иногда без него.Детерминанта вирулентности agg всегда была связана с присутствием детерминант феромона. Никакие другие пары детерминант не встречались исключительно вместе. Большинство штаммов (> 89%) обладали детерминантой efaAfs . Для детерминант gelE, cylMBA и esp была очевидна четкая тенденция. Стартовые штаммы имели меньше детерминант вирулентности, чем пищевые штаммы (10 против 49%), у которых, в свою очередь, было меньше, чем у медицинских штаммов (58%).
(ii)
E.faecium и E. durans .Значительно отличная картина наблюдалась у штамма E. faecium . Все штаммы не содержали детерминант agg , cylMBA и полового феромона. Все стартовые штаммы E. faecium не имели детерминант вирулентности, за исключением efaAfm . Для некоторых пищевых и медицинских штаммов E. faecium были получены слабо гибридизующиеся полосы в стандартных условиях, которые отсутствовали в условиях высокой жесткости, с использованием зонда, специфичного для efaAfs .Продукты ПЦР-амплификации получали с использованием праймеров, специфичных для efaAfs для этих штаммов. Однако для исходных штаммов E. faecium не было получено сигнала с зондом efaAfs при гибридизации по Саузерну, и не было получено никаких продуктов с использованием праймеров, специфичных для efaAfs . Из трех протестированных штаммов E. durans стартовый штамм F25 не гибридизировался ни с одним из зондов efaA , ни с продуктами амплификации с праймерами, специфичными для efaAfs или efaAfm .Пищевые штаммы E. durans, F24 и F26 давали наиболее сильно гибридизующиеся полосы с зондами, специфичными для efaAfm и efaAfm , соответственно (данные не показаны).
Ген gelE был идентифицирован в одном медицинском штамме E. faecium , P20, но активности желатиназы в этом штамме обнаружить не удалось (см. Ниже). Интересно, что ген esp был идентифицирован с высокой частотой в медицинских штаммах (78%) и является исключительным для этой группы; детерминант esp не был обнаружен ни в одном из заквасочных или пищевых штаммов.Эта частота также значительно выше, чем у штамма E. faecalis .
Три штамма (два из E. faecium и один из E. durans ) не несли ни одного из генов вирулентности.
Silent гены присутствуют в заквасочных, пищевых и медицинских штаммах.
Гены Silent gelE встречались у штамма E. faecalis из всех групп. Активность желатиназы не могла быть обнаружена в E. faecalis F41, F1, F4, P21, P26, P31 и P36, хотя эти штаммы несли ген gelE .Один молчащий ген был идентифицирован в штамме E. faecium , P20. В медицинских штаммах E. faecalis P14 и E. faecium P20 активность желатиназы была потеряна во время пересева из исходного материала, и активность желатиназы не была обнаружена. Два других медицинских штамма. E. faecalis P21 и P36, по-видимому, не обладают гемолитической активностью, но несут Cyl гена. Наблюдалась тенденция, при которой у заквасок было меньше молчащих генов, чем у пищевых штаммов, которые, в свою очередь, имели меньше, чем у медицинских штаммов.
Перенос плазмид вирулентности из медицинских штаммов в заквасочные пищевые штаммы путем конъюгации.
Перенос pAD1 в E. faecalis F41 путем спаривания фильтров достигался с частотой 2,1 × 10 -6 на донора. Спаривания в бульоне, использованные для получения FI9530, позволяли переносить pAD1 в E. faecalis FA2-2 с частотой 3,6 × 10 -5 . От этого донора pAD1 был введен в E. faecalis F27 путем спаривания фильтров с частотой 5.7 × 10 −3 . E. faecalis FI9530 использовали непосредственно в вязках со стартовым штаммом E. faecium F12 с использованием селекции lacZ . Несколько попыток сочетания бульонов и фильтров с использованием различных соотношений донор-реципиент (данные не показаны) не привели к образованию каких-либо трансконъюгантов.
ОБСУЖДЕНИЕ
Энтерококки, как известно, вызывают серьезные заболевания у людей и становятся серьезной и растущей проблемой при внутрибольничных инфекциях из-за приобретения детерминант устойчивости к антибиотикам.Приобретенная устойчивость к антибиотикам в сочетании с естественной устойчивостью к нескольким основным классам антибиотиков и естественной устойчивостью этих организмов к низким pH, высоким концентрациям солей и высоким температурам способствует их выживанию. Энтерококки обнаруживаются в пищевых продуктах как загрязнители, и их намеренно добавляют в пищу в качестве заквасок. Энтерококки являются условно-патогенными микроорганизмами, и иммунный статус хозяина является фактором, определяющим их способность вызывать заболевание. Несмотря на их значение, мало что известно о механизмах энтерококковой вирулентности, которые вносят вклад в патогенез; тем не менее, несколько факторов были задействованы как потенциальные детерминанты вирулентности.Используя подход к скринингу ДНК для сравнения встречаемости этих детерминант вирулентности в заквасочных, пищевых и медицинских штаммах, мы выявили различные тенденции встречаемости этих детерминант.
Штаммы E. faecalis и E. faecium демонстрируют значительно различающиеся закономерности встречаемости детерминант вирулентности. Все протестированные штаммы E. faecalis обладают множественными детерминантами вирулентности. Заболеваемость выше у медицинских штаммов, чем у пищевых или заквасочных штаммов.Все штаммы E. faecium , как правило, не содержат детерминант вирулентности, но есть заметные исключения. В этом исследовании, хотя все протестированные штаммы E. faecium не содержали детерминант cylMBA и agg , один медицинский штамм имел gelE и большинство штаммов E. faecium и E. faecium. медицинских штамма имели детерминанты efaAfm и esp соответственно. Это контрастирует с другой работой, в которой gelE и esp были обнаружены исключительно в E.faecalis и не входил ни в один из протестированных штаммов E. faecium (10, 35). Эти различия могут объясняться региональными вариациями. С другой стороны, появление штаммов esp + E. faecium , способных продуцировать этот новый белок клеточной стенки, может быть связано с увеличением числа патогенных штаммов E. faecium . Дальнейшая работа по изучению этих возможностей продолжается.
Определитель efaA обнаружен на аналогичных частотах в E.faecalis и E. faecium и присутствует во всех трех группах. Однако различные степени гомологии между генами efaA у разных видов энтерококков были продемонстрированы с помощью саузерн-блоттинга. Ген штамма E. faecium стартового штамма efaA , по-видимому, менее похож, чем ген E. faecium пищевой или медицинский штамм efaA на ген E. faecalis efaA . Это может указывать на значительное расхождение последовательностей среди E.faecium стартовые штаммы. Штаммы E. durans давали вариабельные полосы или их отсутствие на саузерн-блотах с использованием зондов efaA . Если предположить, что E. durans содержит гомолог efaA , то это может указывать на дальнейшее расхождение последовательностей. До сих пор только ген efaAfs влиял на патогенность на животных моделях (38). Роль efaAfm еще не продемонстрирована. Значительные вариации последовательности в E.faecium и E. durans могут вызывать функциональные различия в адгезине EfaA, влияющие на патогенность. В качестве альтернативы для этих штаммов могут потребоваться другие факторы вирулентности в сочетании с EfaA. Продукты ПЦР-амплификации получали с использованием праймеров, специфичных для efaAfs , для некоторых пищевых и медицинских штаммов E. faecium . Анализ последовательности этих фрагментов должен быть полезен для определения присутствия адгезина в этих штаммах.
Несколько штаммов, включая стартовый, содержали явно неактивные детерминанты gelE и cyl . В случае E. faecalis P21 отсутствие гемолитической активности объясняется отсутствием cylA , который необходим для активации cylL L / S . Для других штаммов отсутствие фенотипической активности может быть объяснено низким уровнем или понижающей регуляцией экспрессии гена или неактивным генным продуктом. Известно, что факторы окружающей среды влияют на экспрессию генов (12), а условия, используемые для фенотипического тестирования, отличаются от тех, которые встречаются в организме человека.Тихие гены также могут активироваться временными факторами, такими как условия, обнаруживаемые в желудочно-кишечном тракте, баланс организмов в кишечной флоре и эффекты бактериального синергизма, а также наличие и сохранение большого количества жизнеспособных энтерококков. Недавно была описана опосредованная феромонами система распознавания кворума. (J. Nakayama, Y. Cao, A. D. L. Akkermans, W. M. DeVos и H. Nagasawa, Abstr. 1st Int. ASM Conf. Enterococci: Pathog. Biol. Antibiot. Resist. Abstr.21, стр. 30). В этой системе ген gelE является частью того же оперона, что и гены гистидинкиназы, чувствительной к феромонам. Экспрессия gelE запускается в поздней экспоненциальной фазе при высокой плотности клеток, когда это может быть преимуществом. У медицинских штаммов была более высокая частота молчащих генов, чем у пищевых и стартовых штаммов. Это коррелирует с более высоким потенциалом патогенных эффектов у этих штаммов. Таким образом, наличие явно молчаливых генов как в пищевых, так и в заквасочных штаммах имеет значение для их безопасного использования в пищевых продуктах.
До недавнего времени большинство изолятов инфекционного происхождения составляли штамм E. faecalis , и поэтому он считается наиболее патогенным видом энтерококков. Роль ряда факторов вирулентности в патогенезе, включая gelE, esp и первоначально efaA , подтверждается тем фактом, что эти детерминанты были обнаружены только у штамма E. faecalis . В этом исследовании большинство охарактеризованных детерминант вирулентности было обнаружено у E.faecalis , что является дополнительным подтверждением их роли в патогенезе. Однако частота инфекций, вызванных E. faecium , увеличивается (B. Murray, Abstr. 1st Int. ASM Conf. Enterococci: Pathog. Biol. Antibiot. Resist. Abstr. 516, p. 16). Штаммы E. faecium , содержащие эти детерминанты вирулентности, могут быть вовлечены в эволюцию патогенных штаммов E. faecium .
Энтерококки обладают высокоэффективными механизмами переноса генов, включая конъюгацию и конъюгативную транспозицию.Известно, что гены вирулентности связаны с некоторыми высокотрансмиссивными плазмидами. Таким образом, существует риск того, что безопасный штамм, в котором отсутствуют гены известных факторов вирулентности, может приобрести такие гены путем конъюгации. В контексте заквасок и пробиотических штаммов возможно потребление очень большого количества жизнеспособных бактерий. Это обеспечило бы большую популяцию реципиентов, в которой могли бы распространяться трансмиссивные плазмиды, несущие гены вирулентности. Возможность такого события была исследована с использованием стартовых штаммов E.faecium F12 и E. faecalis F27 и F41. Возможность переноса генов вирулентности в исходные штаммы тестировали с использованием индуцируемой феромонами системы конъюгации E. faecalis DS16.
Перенос детерминант вирулентности из E. faecalis DS16 через плазмиду полового феромона pAD1 в исходные штаммы E. faecalis был достигнут с частотами, аналогичными тем, которые были получены при скрещивании в бульоне с использованием свободных от плазмид штаммов реципиентов (40).Таким образом, можно было продемонстрировать, что стартовые штаммы могут приобретать известные гены вирулентности посредством процесса переноса генов естественной конъюгации. Будет интересно определить, увеличивает ли приобретение pAD1 штаммами F27 и F41 E. faecalis их вирулентность. Для ответа на этот вопрос планируются контрольные испытания на животных.
Процесс конъюгации, используемый плазмидами, такими как pAD1, является высокоразвитым и включает сложный процесс передачи сигналов. Штаммы потенциальных реципиентов секретируют небольшие сигнальные молекулы пептидов, известные как половые феромоны.Эти феромоны индуцируют гены, ведущие к производству вещества агрегации в потенциальных донорных клетках, что приводит к фенотипу слипания клеток. Этот процесс увеличивает частоту переноса плазмиды полового феромона в свободные от плазмиды клетки-реципиенты. Каждая плазмида полового феромона специфически реагирует на соответствующую детерминанту полового феромона. Плазмида полового феромона pAD1 обнаружена исключительно у штамма E. faecalis (6) и специфически реагирует на половой феромон cAD1. E.faecium F12 не содержит детерминант полового феромона cad , и поэтому использовались согласования фильтров в попытке увеличить частоту переноса в отсутствие индуцированного скопления клеток. Несмотря на неоднократные попытки переноса pAD1 в стартовый штамм E. faecium F12 посредством скрещивания с E. faecalis FI9530, переноса достичь не удалось.
Детерминанты половых феромонов cpd, cob, ccf и cad были обнаружены во всех E.faecalis протестированных штамма. В некоторых из этих штаммов также присутствовал ген agg . Эта ситуация соответствует штаммам-реципиентам, которые приобрели плазмиду полового феромона. Штаммы с детерминантами половых феромонов обладают потенциалом для приобретения соответствующих плазмид половых феромонов и, следовательно, связанных детерминант вирулентности. В этом исследовании все из протестированных пищевых штаммов E. faecalis обладали способностью приобретать несколько плазмид половых феромонов. Для E.faecium , отсутствие генов половых феромонов может отражать дивергенцию последовательностей, а не отсутствие феромонов переноса генов. Действительно, была установлена перекрестная связь половых феромонов между E. faecium и E. faecalis (18).
Перенос гена Enterococcus и системы, чувствительные к феромонам, особенно важны для проблем, связанных с приобретенной устойчивостью к гликопептидам. Приобретенная устойчивость к гликопептидам в основном связана с фенотипом VanA и локализована на больших конъюгативных плазмидах.Некоторые плазмиды, конъюгированные с VanA, также чувствительны к феромонам (30). Устойчивые к гликопептидам энтерококки стали серьезной проблемой для здоровья в отделениях интенсивной терапии в Европе и США; где эти антибиотики используются в качестве резервных антибиотиков против мультирезистентных грамположительных патогенов. Хитон и др. (18) показали, что клинический изолят E. faecium содержал конъюгативную плазмиду, сильно родственную pCF10 из E. faecalis . Эта плазмида обеспечивала высокий уровень устойчивости к ванкомицину, который можно было передать конъюгацией на свободный от плазмиды E.faecalis получателя. Связанные детерминанты вирулентности agg и cyl могут быть котрансферными и будут отобраны по устойчивости к антибиотикам. Кроме того, присутствие agg в этих штаммах может привести к увеличению способности к колонизации. Таким образом, продукция полового феромона пищевым штаммом E. faecalis может способствовать приобретению устойчивости к ванкомицину и другим связанным признакам у штамма E. faecium и вести к повышенной вирулентности.
В заключение, мы определили закономерности присутствия как экспрессированных, так и скрытых генов вирулентности в заквасочных, пищевых и медицинских штаммах энтерококков. Присутствие этих детерминант вирулентности у штаммов E.
