
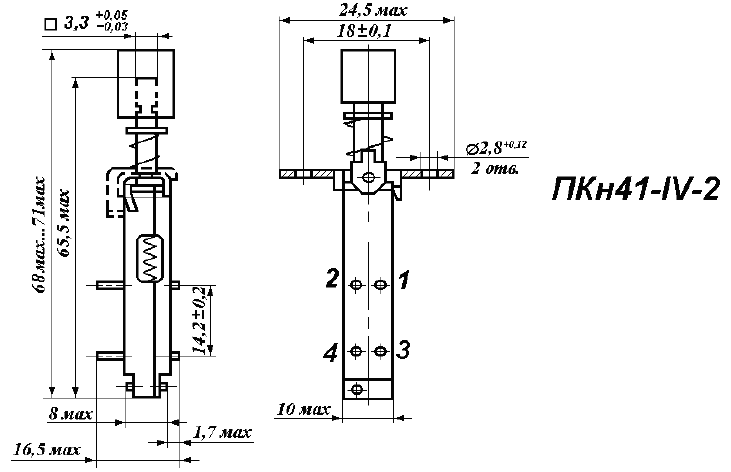
Переключатель ПКН41-1-2 –
Переключатель ПКН41-1-2
Масса и наименование драгоценных металлов которые содержатся в переключателе ПКН41-1-2 на основании документации и справочных формуляров и описаний. Справочник содержания драгметаллов в переключателе электрическом с указанием его веса которые используются (или использовались) при производстве переключателей.
Содержание драгоценных металлов в переключателе ПКН41-1-2.
Золото: 0 грамм.
Серебро: 0.023 грамм.
Платина: 0 грамм.
Палладий: 0 грамм.
Источник справочной информации: .
Если у вас есть информация о переключателе ПКН41-1-2 и его способам использования сообщите ее нам мы бесплатно разместим ее на сайте.
Фото переключатель ПКН41-1-2:
Характеристики переключатель ПКН41-1-2:
Купить или продать а также цены на ПКН41-1-2:
Оставьте отзыв или бесплатное объявление о покупке или продаже переключателя марки ПКН41-1-2:
Переключатель ПР2-5П2Н
Масса и наименование драгоценных металлов которые содержатся в переключателе ПР2-5П2Н на основании документации и справочных . ..
..
Переключатель ПТ6-12Т
Масса и наименование драгоценных металлов которые содержатся в переключателе ПТ6-12Т на основании документации и справочных …
Переключатель ПП10
Масса и наименование драгоценных металлов которые содержатся в переключателе ПП10 на основании документации и справочных …
Переключатель ПКУ-2-11
Масса и наименование драгоценных металлов которые содержатся в переключателе ПКУ-2-11 на основании документации и справочных …
Сайт по Драгметаллам в различных устройствах посвящен только основным драгоценным металлам: золоту, серебру, платине и палладию.
Новини
|
 СЕРЕБРО. Вильгельм 2. Пруссия. Германия. Редкие! Состояние! №14Б-41-1.2
СЕРЕБРО. Вильгельм 2. Пруссия. Германия. Редкие! Состояние! №14Б-41-1.2 00 р. Торг уместен
00 р. Торг уместен 00 р.
00 р.
 00 р
00 р В1 + 2 рубля 2019 ММД шт.В. Монограмма приподнята на обеих монетах.(5)
В1 + 2 рубля 2019 ММД шт.В. Монограмма приподнята на обеих монетах.(5) ..
.. ,MiNr.56 F y,зубцовка 10 1/2
,MiNr.56 F y,зубцовка 10 1/2 № 1,2,3,4,6,7,8,9 – 1984 г. – 8 штук
№ 1,2,3,4,6,7,8,9 – 1984 г. – 8 штук 00 р. 0 ставок
00 р. 0 ставок 00 р. 0 ставок
00 р. 0 ставок 1224-35 1 год Правительствa национального единства и примирения,MNH[8.81]
1224-35 1 год Правительствa национального единства и примирения,MNH[8.81] 00 р. блиц-цена
00 р. блиц-цена 00 р.
00 р. истребитель-штурмовик F/A-18.HORNET (Шершень).2
истребитель-штурмовик F/A-18.HORNET (Шершень).2Перекл. МИ3А ; ПТИ-М; ПБР-2м ; Д701; МП1-1 ; ПКН41-1-2 ;ПМ-3П3Н
Поделиться
Пожаловаться
1) Д701 Микровыключатель НО.360.011 ( 7Ш3.603.000ЭТ ) (90-91-92г) зав.уп.,паспорт (ОАО *Электроавтомат* Алатырь) — 301шт х 80руб
Д701 Микровыключатель НО. 360.011 ( 7Ш3.603.000ЭТ ) (87г) зав.уп.,паспорт (ОАО *Электроавтомат* Алатырь) – 108 шт х 50руб
360.011 ( 7Ш3.603.000ЭТ ) (87г) зав.уп.,паспорт (ОАО *Электроавтомат* Алатырь) – 108 шт х 50руб
2) МИ3А Микропереключатель ( с лепестком) 220В ; 2А ТУ-25-7135.0001-87 зав.уп. (з-д Сигма Литва) (88-91г) –199шт х 120руб
3) М-405 Микропереключатель ( запаяны в пленке ) 10шт х 20руб
4) МП1107Л УХЛ3 Микропереключатель 660V-440V-10А (знак кач-ва) (89г) 1шт х250руб
МП2101 ЛУХЛ4 Микропереключатель 660V-440V-16А (83г) 1шт х 100р
5) МП1-1 Микропереключатель УСО.360.074ТУ (89-90г) Кузбассрадио зав.уп. 55 х 105руб
6) ПМ3-1В Микропереключатель двухполюсный (ОЮО.360.072ТУ )«НПП «КРИПТОН» зав.уп.(2001г) — 30 шт
7) ПМ-3П3Н *5* Галетный малогабаритный с клювиком , карболит ( ЕЫО.360.001 ТУ) (91г )зав.уп.–128шт х 70руб
8) ПЩ-12П3Н2 Переключатель щеточный ЕЩ0360600ТУ зав.уп.—12шт х 550руб
9) ПЩ-3П1Н2 Переключатель щеточный ТУУСО.360.059 зав.уп.–13шт х 150руб
10) ПКН41-1-2 Переключатель кнопочный (Ю 60 360006) ( 2002г) Смоленский з-д радиодеталей зав.уп. –96шт х 15руб
11) П2К-Н-10-15-2 Переключатель кнопочный ( независимая фиксация ) (ТУ -ЕЩ0. 360.037 ) АО ЛТАВА ( 94г) зав.уп. 8 х 15р
360.037 ) АО ЛТАВА ( 94г) зав.уп. 8 х 15р
12) ПТИ-М Переключатель выбора точек ( АЛД 3.602.022.) ГОСТ15150-069 // ТУ25.08.116-77 зав.уп. -172 шт х170руб
13) П3ПН-20 (20А, 27В) Переключатель ТУ16-526.016-73 (87г ) зав.уп. 9 х 150руб
14) 2ППН-45 ( 20А, 27В ) Тумблер ,переключатель (80г) 6шт х 290р
15) Пускатель ПБР-2м (220В: 50Гц)( 86г) зав.уп. -1шт х 1000руб
16) Выключатель пакетный ПВ2-10 (10А; 220В) СССР -8шт х 70р
17) Выключатель пакетный ПВ2-16 (16А; 220В) Электротехник -8шт х 70р
Контакты
| Ф.И.О. | Светлана Светлана ★★★★★ 5.00 |
|---|---|
| Компания: | техавто |
| Страна: | Россия |
| Телефон: | 8-919-642-2191 Сообщите, что нашли информацию на сайте «Элек.ру» |
| Дата регистрации: | 19 марта 2012 г. Последний вход вчера |
| Отправить сообщение |
 elec.ru/doska/elektrogidrorele-keg-rele-potoka-rpi-1238390053/”>ВКонтакте
elec.ru/doska/elektrogidrorele-keg-rele-potoka-rpi-1238390053/”>ВКонтакте
ННБ37-60-090-I (Эра) | Старый Свет
«Суровая настольная лампа» времён развитого социализма. Подобный «робототехнический» дизайн начал входить в моду в конце первой половины 80-х, одновременно с активным применением пластмасс и подчёркнуто яркими «дизайнерскими» цветами. Можно предположить, что это была попытка снять кальку с современных на тот момент тенденций буржуазной моды. Ну а так как снимали её наши тяжеловесные предприятия, это добавляло такой продукции свой неповторимый отпечаток
Данная настольная лампа у основания снабжена чем-то вроде пепельницы, предназначенной очевидно для хранения канцелярской мелочёвки. Основание в итоге получилось непропорционально маленьким, а главное тонким, в результате чего основной вес лампы рассредоточен между основанием и вертикальной стойкой. Конструкция, соответственно, получилась неуклюжей и склонной к заваливанию.
Основание в итоге получилось непропорционально маленьким, а главное тонким, в результате чего основной вес лампы рассредоточен между основанием и вертикальной стойкой. Конструкция, соответственно, получилась неуклюжей и склонной к заваливанию.
В качестве выключателя питания также применён «суровый», позаимствованный у электронных приборов, переключатель П2К (аналогичные использовались также в советских телевизорах 3 поколения). Весьма приличные габариты этого монстра удачно скрыты в утолщённой горловине лампы и прикрыты квадратной кнопочкой (фото 2). Жаль только, что надёжность сей конструкции получилась не очень высокая – срабатывает она не всегда с первого нажатия.
Новаторский характер этого светильника подчёркивает его полностью пластмассовая внешняя облицовка, все металлические детали скрыты внутри. Одной из таких деталей служит белый отражатель (фото 3), который не позволяет теплу лампы проплавить красивые вентиляционные отверстия на корпусе. К чести этого светильника надо сказать, что несмотря на заявленную максимальную мощность лампы в 60 ватт, домашние по незнанию ставили в неё и 75-, и 100-ваттные лампочки, и каких-либо видимых повреждений это не вызвало.
Дату выпуска на корпусе обнаружить не удалось, равно как и указания госцены (на предполагаемом месте которой имеется выштампованное на пластмассе окошко). Зато моделей указано целых две: ННБ37-60-090 и то же самое с -I в конце. С учётом того, что приобретена эта лампа была не позже 1992 г., делаем предположение, что она выпущена во второй половине 1991 г. или в самом начале 1992 г. В любом случае, модель ещё 100% советская, поэтому страна происхождения пусть будет СССР…
Пара железо-чувствительных генов, кодирующих протеинкиназы с Ser / Thr-киназным доменом и гист-киназным доменом, регулируется NtcA в Cyanobacterium Anabaena sp. Штамм PCC 7120
СПИСОК ЛИТЕРАТУРЫ
1. Альфонсо, М., И. Перевоска, Д. Кириловский. 2001. Редокс-контроль экспрессии гена ntcA
у Synechocystis sp. PCC 6803. Доступность азота и транспорт электронов
регулируют уровни белка NtcA. Plant Physiol.
125: 969–981.
2. Байчу, Н. , Т. Ван, Р. Йе и Дж. Д. Хельманн. 2002. Глобальный анализ регулона
, Т. Ван, Р. Йе и Дж. Д. Хельманн. 2002. Глобальный анализ регулона
Bacillus subtilis Fur и стимулятора железного голодания. Мол. Microbiol.
45: 1613–1629.
3. Бауэр, К. К., Л. Скаппино и Р. Хазелкорн. 1993. Рост cyanobac-
terium Anabaena на молекулярном азоте: NifJ требуется, когда железо ограничено. Proc. Natl. Акад. Sci. США 90: 8812–8816.
4. Бурнап Р.Л., Троян Т., Л.А.Шерман. 1993. Обильный хлорофилл-белковый комплекс
железодефицитных Synechococcus sp. PCC 7942
(CP43⬘) кодируется геном isiA. Plant Physiol. 103: 893–902.
5. Cai, Y., and C.P. Wolk. 1990. Использование условно летального гена у Anabaena
sp. штамм PCC 7120 для отбора двойных рекомбинантов и включения последовательностей вставки
. J. Bacteriol. 172: 3138–3145.
6. Cai, Y., and C.P. Wolk. 1997. Азотная депривация Anabaena sp.штамм
PCC 7120 вызывает быструю активацию кластера генов, который необходим для поглощения
и утилизации нитрата. J. Bacteriol. 179: 258–266.
J. Bacteriol. 179: 258–266.
7. Коццоне, А. Дж., К. Гранжасс, П. Дублет и Б. Дюкло. 2004. Фосфорилирование белка
по тирозину в бактериях. Arch. Microbiol. 181: 171–181.
8. Эльхай Дж. И К. П. Волк. 1988. Конъюгальный перенос ДНК цианобактериям.
Методы Enzymol. 167: 747–754.
9. Escolar, L., Дж. Перес-Мартин и В. Лоренцо. 1999. Открытие железного ящика:
Металлорегуляциятранскрипции с помощью белка Fur. J. Bacteriol. 181: 6223–
6229.
10. Флорес, Э. и А. Эрреро. 1994. Ассимиляционный азотный обмен и его регуляция
, с. 487–517. В Д. А. Брайанта (ред.), Молекулярная биология
цианобактерий. Kluwer Academic Publishers, Дордрехт, Нидерланды.
11. Frı´as, J. E., E. Flores, and A. Herrero. 1994. Требование регуляторного белка NtcA
для экспрессии генов ассимиляции азота и развития гетероцист
у цианобактерий Anabaena sp.PCC 7120. Mol.
Microbiol. 14: 823–832.
12. Frı´as, J. E., E. Flores, and A. Herrero. 1997. Кластер генов ассимиляции нитратов
E., E. Flores, and A. Herrero. 1997. Кластер генов ассимиляции нитратов
из гетероцистообразующей цианобактерии Anabaena sp. штамм PCC 7120.
J. Bacteriol. 179: 477–486.
13. Frı´as, J. E., E. Flores, and A. Herrero. 2000. Для активации промотора оперона Anabaena nir
требуются факторы транскрипции как NtcA (семейство CAP), так и NtcB (семейство LysR)
. Мол. Microbiol.383: 613–625.
14. Geiss, U., J. Vinnemeier, A. Kunert, B. Lindner, B. Gemmer, M. Lorenz, M.
Hagemann, and A. Schoor. 2001. Обнаружение гена isiA в териальных штаммах цианобактерии
: потенциал для исследования дефицита железа. Прил. Environ. Microbiol.
67: 5247–5253.
15. Хэнкс, С. К., и Т. Хантер. 1995. Суперсемейство эукариотических протеинкиназ
ily: структура и классификация каталитических доменов киназ. FASEB J. 9: 576–596.
16.Харди, Л. П., Д. Л. Балквилл и С. Е. Стивенс, мл. 1983. Влияние голодания на железо
на физиологию цианобактерий Agmenellum quad-
ruplicatum. Прил. Environ. Microbiol. 45: 999–1006.
Прил. Environ. Microbiol. 45: 999–1006.
17. Харди, Л. П., Д. Л. Балквилл, и С. Е. Стивенс, мл. 1983. Влияние голодания на железо
на ультраструктуру цианобактерий Agmenellum quad-
ruplicatum. Прил. Environ. Microbiol. 45: 1007–1017.
18. Эрреро А., Муро-Пастор А.М., Э.Флорес. 2001. Контроль азота в
цианобактериях. J. Bacteriol. 183: 411–425.
19. Эрреро, А., А. М. Муро-Пастор, А. Валладарес и Э. Флорес. 2004. Клеточная дифференцировка
и фактор транскрипции NtcA в филаментных цианобактериях
терий. FEMS Microbiol. Откр. 28: 469–487.
20. Цзян Ф., Б. Маннервик и Б. Бергман. 1997. Доказательства редокс-регуляции –
фактора транскрипции NtcA, действующего как активатор, так и репрессор
в цианобактериях Anabaena PCC 7120.Biochem. J. 327: 513–
517.
21. Цзян Ф., С. Визен, М. Вилдерстен, Б. Бергман и Б. Маннервик. 2000.
Исследование мотива связывания фактора транскрипции NtcA с помощью in vitro выборки последовательностей ДНК из случайной библиотеки. J. Mol. Биол. 301: 783–793.
J. Mol. Биол. 301: 783–793.
22. Лаббе Д., Дж. Гарнон и П. К. Лау. 1997. Характеристика генов
, кодирующих рецептор-подобную His-киназу и родственный регулятор ответа из
бактерии, разлагающей бифенил / полихлорбифенил, Rhodococcus sp.штамм
М5. J. Bacteriol. 179: 2772–2776.
23. Лоран, С., Х. Чен, С. Беду, Ф. Зиарелли, Л. Пэн и К.-К. Чжан. 2005.
Неметаболизируемый аналог 2-оксоглутарата вызывает дифференцировку гетероцист-
в репрессивных условиях у Anabaena sp. PCC 7120. Proc. Natl.
Акад. Sci. США 102: 9907–9912.
24. Леонхардт К. и Н. А. Штраус. 1994. Гены фотосистемы II isiA, psbDI и
psbC у Anabaena sp. PCC 7120: клонирование, секвенирование и регуляция транскрипции
в стрессированных железом и избыточных железом клетках.Завод Мол. Биол. 24: 63–73.
25. Луке И., Э. Флорес и А. Эрреро. 1993. Ген нитритредуктазы из
Synechococcus sp. PCC 7942: гомология между цианобактериальными и высшими нитритредуктазами
растений. Завод Мол. Биол. 21: 1201–1205.
Завод Мол. Биол. 21: 1201–1205.
26. Луке И., Э. Флорес и А. Эрреро. 1994. Молекулярный механизм операции
контроля азота у цианобактерий. EMBO J. 13: 2862–2869.
27. Муро-Пастер, А. М., А. Валладарес, Э. Флорес и А.Herreo. 1999. Ген hetC
является прямой мишенью регулятора транскрипции NtcA в развитии гетероцист цианобактерий
. J. Bacteriol. 181: 6664–6669.
28. Омори, М., М. Икеучи, Н. Сато, П. Волк, Т. Канеко, Т. Огава, М. Канехиса,
С. Гото, С. Кавасима, С. Окамото, Х. Йошимура , H. Katoh, T. Fujisawa, S.
Ehira, A. Kamei, S. Yoshihara, R. Narikawa, and S. Tabata. 2001. Характеристика
генов, кодирующих мультидоменные белки в геноме филамен-
азотфиксирующих цианобактерий Anabaena sp.штамм PCC 7120. DNA Res.
8: 271–284.
29. Outten, F. W., O. Djaman, and G. Storz. 2004. Требуется суфоперон для сборки кластеров
Fe-S во время железного голодания у Escherichia coli. Мол. Микро-
биол. 52: 861–872.
52: 861–872.
30. Phalip, V., J.-H. Ли и Ч.-К. Чжан. 2001. HstK, цианобактериальный белок
как с доменом серин / треонинкиназы, так и с доменом гистидинкиназы:
Значениедля механизма передачи сигнала.Biochem. J. 360: 639–
644.
31. Prentki, P., and H. M. Krisch. 1984. Инсерционный мутагенез in vitro с выборочным фрагментом ДНК
. Ген 29: 303–313.
32. Рамасубраманян, Т.С., Т.-Ф. Wei и J. W. Golden. 1994. Две Анабаена
сп. ДНК-связывающие факторы штамма PCC 7120 взаимодействуют с генами, специфичными для вегетативных клеток и
гетероцист. J. Bacteriol. 176: 1214–1223.
33. Rippka, R., J. Deruelles, J. B. Waterbury, M. Herdman, R.Ю. Станье.
1979. Родовые задания: истории штаммов и свойства чистых культур цианобактерий
. J. Gen. Microbiol. 111: 1–61.
34. Сайер, М. Х., младший, 1998. Множественные механизмы, контролирующие метаболизм углерода
в бактериях. Biotechnol. Bioeng. 582: 170–174.
35. Schmitz, O., T. Kentemich, W. Zimmer, B. Hundeshagen и H. Bothe.
1993. Идентификация гена nifJ, кодирующего пируват: ферредоксинокси-
доредуктазу в диазотфиксирующих цианобактериях.Arch. Microbiol. 160: 62–67.
36. Schmitz, O., J. Gurke, and H. Bothe. 2001. Молекулярные доказательства аэробной экспрессии
nifJ, кодирующего пируват: ферредоксин оксидоредуктазу, у
цианобактерий. FEMS Microbiol. Lett. 195: 97–102.
37. Шерман, Д. М., и Л. А. Шерман. 1983. Влияние дефицита железа и восстановления железа
на ультраструктуру Anacystis nidulans. J. Bacteriol. 156: 393–401.
38. Stanier, R. Y., and G. Cohen-Bazire.1977 г. Фототрофные прокариоты: цианобактерии
. Анну. Rev. Microbiol. 31: 225–274.
39. Сток, А. М., В. Л. Робинсон, и П. Н. Гудро. 2000. Двухкомпонентное преобразование сигнала
. Анну. Rev. Biochem. 69: 183–215.
40. Штраус, Н. А. 1994. Депривация железа: физиология и регуляция генов, с.
731–750. В Д. А. Брайанте (ред.), Молекулярная биология цианобактерий.
Kluwer Academic Publishers, Дордрехт, Нидерланды.
41.Су, З.-К., В. Ольман, Ф. Мао, Ю. Сюй. 2005. Сравнительный анализ геномики
регулонов NtcA у цианобактерий: регуляция ассимиляции азота
и ее связь с фотосинтезом. Nucleic Acids Res. 33: 5156–5171.
42. Валладарес, А., А. М. Муро-Пастор, А. Эрреро и Э. Флорес. 2004.
NtcA-зависимый промотор P1 используется для экспрессии glnA в N
2
, фиксируя
гетероцист Anabaena sp. штамм PCC 7120.J. Bacteriol. 186: 7337–7343.
43. Вега-Палас, М.А., Э. Флорес и А. Эрреро. 1992. NtcA, глобальный регулятор азота
из цианобактерии Synechococcus, которая принадлежит к семейству бактериальных регуляторов Crp
. Мол. Microbiol. 6: 1853–1859.
44. Виннемайер, Дж., А. Кунерт и М. Хагеманн. 1998. Транскрипционный анализ
оперона isiAB в подвергнутых солевому стрессу клетках цианобактерии Synechocystis
sp. PCC 6803. FEMS Microbiol.Lett. 169: 323–330.
PCC 6803. FEMS Microbiol.Lett. 169: 323–330.
45. Ван, Л., Ю.-П. Солнце, W.-L. Чен, Ж.-Х. Ли и Ч.-К. Чжан. 2002. Genomic
анализ протеинкиназ, протеинфосфатаз и двухкомпонентных регуляторных систем цианобактерий Anabaena sp. штамм PCC 7120. FEMS
Microbiol. Lett. 217: 155–165.
46. Wei, T.-F., T. S. Ramasubramanian, and J. W. Golden. 1994. Anabaena sp.
Штамм, ген ntcA PCC 7120, необходимый для роста на нитратах и гетероцистах
.J. Bacteriol. 176: 4473–4482.
47. Волк, К. П., А. Эрнст и Дж. Эльхай. 1994. Метаболизм и развитие гетероцист-
, с. 769–823. В Д. А. Брайанта (ред.), Молекулярная биология циано-
бактерий. Kluwer Academic Publishers, Дордрехт, Нидерланды.
48. Xu, W.-L., R. Jeanjean, Y.-D. Лю и К.-К. Чжан. 2003. pkn22 (alr2502)
, кодирующий предполагаемую киназу Ser / Thr в цианобактериях Anabaena sp. PCC
7120 индуцируется как железным голоданием, так и окислительным стрессом и регулирует экспрессию
isiA. FEBS Lett. 553: 179–182.
FEBS Lett. 553: 179–182.
49. Zhang, C.-C. 1993. Ген, кодирующий белок, относящийся к киназе эукариотического белка
из филаментной гетероцистной цианобактерии Anabaena PCC
7120. Proc. Natl. Акад. Sci. USA 90: 11840–11844.
50. Zhang, C.-C., and L. Libs. 1998. Клонирование и характеристика гена pknD
, кодирующего протеинкиназу эукариотического типа в цианобактериях
Anabaena sp. PCC7120. Мол. Genet Genet. 258: 26–33.
51.Zhang, C.-C., J. Jang, S. Sakr, and L. Wang. 2005. Фосфорилирование белка
по остаткам Ser, Thr и Tyr у цианобактерий. J. Mol. Microbiol. Biotechnol.
9: 154–166.
52. Zhang, Z., G. Gosset, R. Barabote, CS Gonzalez, WA Cuevas, и MH
Saier, Jr. 2005. Функциональные взаимодействия между углеродом и железом. , в кишечной палочке. J. Bacteriol. 187: 980–990.
ТОМ. 188, 2006 ntcA РЕГУЛИРУЕТ pkn41 И pkn42 В ЦИАНОБАКТЕРИИ 4829
Life | Бесплатный полнотекстовый | Количественное определение 2-оксоглутарата в цианобактериях и протеобактериях на основе биосенсоров
1.
 Введение Внутри клетки координация различных метаболических путей опосредуется колебаниями концентраций конкретных метаболитов, что позволяет аллостерически передавать эту информацию сенсорным белкам, тем самым генерируя соответствующий метаболический ответ. Метаболиты, которые играют ключевую роль в передаче микробных сигналов, включают глутамин, 2-оксоглутарат (α-кетоглутарат, 2-OG), цАМФ и ppGpp [1]. Каждый из этих метаболитов предназначен для передачи информации о наличии определенного класса питательных веществ.2-OG является промежуточным метаболитом цикла трикарбоновой кислоты (TCA или Krebs) и используется в качестве углеродного скелета для ассимиляции азота. Таким образом, он координирует метаболизм углерода и азота. Цикл TCA является важным метаболическим путем, сохраняющимся во всех живых организмах, и обеспечивает не только прекурсоры для биосинтеза большого количества биомолекул, но также снижает мощность производства биологической формы энергии, АТФ, посредством дыхания.
Введение Внутри клетки координация различных метаболических путей опосредуется колебаниями концентраций конкретных метаболитов, что позволяет аллостерически передавать эту информацию сенсорным белкам, тем самым генерируя соответствующий метаболический ответ. Метаболиты, которые играют ключевую роль в передаче микробных сигналов, включают глутамин, 2-оксоглутарат (α-кетоглутарат, 2-OG), цАМФ и ppGpp [1]. Каждый из этих метаболитов предназначен для передачи информации о наличии определенного класса питательных веществ.2-OG является промежуточным метаболитом цикла трикарбоновой кислоты (TCA или Krebs) и используется в качестве углеродного скелета для ассимиляции азота. Таким образом, он координирует метаболизм углерода и азота. Цикл TCA является важным метаболическим путем, сохраняющимся во всех живых организмах, и обеспечивает не только прекурсоры для биосинтеза большого количества биомолекул, но также снижает мощность производства биологической формы энергии, АТФ, посредством дыхания. Уровень 2-OG отражает также углеродный и азотный баланс, сигнальную характеристику, которая сохраняется во всех организмах.Обратная корреляция между доступностью аммония и накоплением 2-OG была продемонстрирована у различных микроорганизмов, таких как Escherichia coli, архей, цианобактерий и почкующихся дрожжей [2,3,4]. Сигнальная роль 2-OG хорошо описана у цианобактерий и особенно у нитчатых гетероцистозных цианобактерий Anabaena sp. штамм PCC 7120 (далее Anabaena). У Anabaena истощение комбинированного азота ведет к накоплению 2-OG, который действует как триггер, вызывающий дифференцировку гетероцист [3,5,6].Гетероцисты – это клетки, специализирующиеся на азотфиксации, которые дифференцируются от вегетативных клеток. Гетероцисты и вегетативные клетки взаимозависимы: гетероцисты поставляют фиксированный азот из атмосферного N 2 в форме аминокислот вегетативным клеткам, в то время как последние используют фотосинтез для фиксации углерода из атмосферного CO 2 и обеспечения сахарозы гетероцистам [5,7 , 8].
Уровень 2-OG отражает также углеродный и азотный баланс, сигнальную характеристику, которая сохраняется во всех организмах.Обратная корреляция между доступностью аммония и накоплением 2-OG была продемонстрирована у различных микроорганизмов, таких как Escherichia coli, архей, цианобактерий и почкующихся дрожжей [2,3,4]. Сигнальная роль 2-OG хорошо описана у цианобактерий и особенно у нитчатых гетероцистозных цианобактерий Anabaena sp. штамм PCC 7120 (далее Anabaena). У Anabaena истощение комбинированного азота ведет к накоплению 2-OG, который действует как триггер, вызывающий дифференцировку гетероцист [3,5,6].Гетероцисты – это клетки, специализирующиеся на азотфиксации, которые дифференцируются от вегетативных клеток. Гетероцисты и вегетативные клетки взаимозависимы: гетероцисты поставляют фиксированный азот из атмосферного N 2 в форме аминокислот вегетативным клеткам, в то время как последние используют фотосинтез для фиксации углерода из атмосферного CO 2 и обеспечения сахарозы гетероцистам [5,7 , 8]. Благодаря своей функции сигнальной молекулы 2-OG связывается с регуляторными белками PII и NtcA. Кристаллические структуры этих двух регуляторов, по отдельности или в комплексе с 2-OG, показали, что они претерпевают значительные конформационные изменения при связывании со своими лигандами [9,10,11,12,13].PII, кодируемый геном glnB, консервативен у бактерий и растений. Он собирается как гомотример и выполняет свою регуляторную функцию посредством межбелковых взаимодействий [14]. У цианобактерий активность PII регулируется фосфорилированием и связыванием эффекторов, таких как АТФ и 2-OG, что позволяет PII принимать разные конформации и взаимодействовать с разными белками-партнерами в разных условиях [15]. В Anabaena тример PII связывается с 3 молекулами PipX в условиях комбинированной депривации азота.Интересно, что PipX и 2-OG имеют один и тот же сайт связывания на PII и поэтому являются взаимоисключающими. Когда концентрация аммиачных клеток уменьшается, уровень 2-OG увеличивается и вытесняет PipX из PII.
Благодаря своей функции сигнальной молекулы 2-OG связывается с регуляторными белками PII и NtcA. Кристаллические структуры этих двух регуляторов, по отдельности или в комплексе с 2-OG, показали, что они претерпевают значительные конформационные изменения при связывании со своими лигандами [9,10,11,12,13].PII, кодируемый геном glnB, консервативен у бактерий и растений. Он собирается как гомотример и выполняет свою регуляторную функцию посредством межбелковых взаимодействий [14]. У цианобактерий активность PII регулируется фосфорилированием и связыванием эффекторов, таких как АТФ и 2-OG, что позволяет PII принимать разные конформации и взаимодействовать с разными белками-партнерами в разных условиях [15]. В Anabaena тример PII связывается с 3 молекулами PipX в условиях комбинированной депривации азота.Интересно, что PipX и 2-OG имеют один и тот же сайт связывания на PII и поэтому являются взаимоисключающими. Когда концентрация аммиачных клеток уменьшается, уровень 2-OG увеличивается и вытесняет PipX из PII. Кроме того, PipX является соактиватором специфичного для цианобактерий транскрипционного фактора NtcA, который принадлежит к семейству гомодимерных регуляторов транскрипции CRP / CAP и использует 2-OG в качестве молекулы-индуктора [16,17]. Одна молекула 2-OG связывается с каждым мономером димера NtcA в кармане, расположенном в N-концевой области.Связывание 2-OG с NtcA уменьшает расстояние между двумя ДНК-связывающими F-спиралями димера NtcA с 37 Å в апо-форме до 34 Å в комплексе 2-OG-NtcA, оптимальное расстояние для связывания ДНК [3, 9]. Активность NtcA также контролируется его взаимодействием с PipX. Каждая субъединица NtcA связывает одну субъединицу PipX в комплексе, и это взаимодействие необходимо для экспрессии гена, опосредованной NtcA. Было высказано предположение, что PipX стабилизирует активную форму NtcA и помогает NtcA рекрутировать РНК-полимеразу [4,10,16,17,18].У Anabaena мутант ntcA неспособен инициировать дифференцировку гетероцист [5,19]. На основании наблюдения, что 2-OG связывается с NtcA и что повышение уровней NtcA и 2-OG запускает дифференцировку гетероцист, было высказано предположение, что взаимодействие между 2-OG и NtcA составляет самый первый шаг в развитии гетероцист [ 3,5].
Кроме того, PipX является соактиватором специфичного для цианобактерий транскрипционного фактора NtcA, который принадлежит к семейству гомодимерных регуляторов транскрипции CRP / CAP и использует 2-OG в качестве молекулы-индуктора [16,17]. Одна молекула 2-OG связывается с каждым мономером димера NtcA в кармане, расположенном в N-концевой области.Связывание 2-OG с NtcA уменьшает расстояние между двумя ДНК-связывающими F-спиралями димера NtcA с 37 Å в апо-форме до 34 Å в комплексе 2-OG-NtcA, оптимальное расстояние для связывания ДНК [3, 9]. Активность NtcA также контролируется его взаимодействием с PipX. Каждая субъединица NtcA связывает одну субъединицу PipX в комплексе, и это взаимодействие необходимо для экспрессии гена, опосредованной NtcA. Было высказано предположение, что PipX стабилизирует активную форму NtcA и помогает NtcA рекрутировать РНК-полимеразу [4,10,16,17,18].У Anabaena мутант ntcA неспособен инициировать дифференцировку гетероцист [5,19]. На основании наблюдения, что 2-OG связывается с NtcA и что повышение уровней NtcA и 2-OG запускает дифференцировку гетероцист, было высказано предположение, что взаимодействие между 2-OG и NtcA составляет самый первый шаг в развитии гетероцист [ 3,5]. Однако роль NtcA не ограничивается инициацией развития гетероцист, поскольку было показано, что он выполняет широкую регуляторную функцию. Реглон NtcA включает несколько сотен генов, включая мишени, участвующие как в углеродном, так и в азотном метаболизме, стрессовой реакции и на различных этапах дифференцировки и функционирования гетероцист [5,20].В дополнение к PII и NtcA, уровни 2-OG также модулируют активность многих регуляторных белков и путей, преимущественно связанных с азотным или углеродным метаболизмом у бактерий, архей, водорослей, растений и животных [1,11,15,21,22, 23]. Несмотря на центральную роль 2-OG и его обширный регуляторный репертуар, нам все еще не хватает инструментов для надлежащего измерения его концентраций и динамики in vivo. Для оценки уровней 2-OG в клеточных экстрактах были разработаны методы ферментативного анализа, масс-спектрометрии или жидкостной хроматографии высокого давления (ВЭЖХ) [4,24,25,26,27,28,29,30].Тем не менее, 2-OG, такой как многие метаболиты, очень быстро превращается и может быть потерян или разложен во время экстракции.
Однако роль NtcA не ограничивается инициацией развития гетероцист, поскольку было показано, что он выполняет широкую регуляторную функцию. Реглон NtcA включает несколько сотен генов, включая мишени, участвующие как в углеродном, так и в азотном метаболизме, стрессовой реакции и на различных этапах дифференцировки и функционирования гетероцист [5,20].В дополнение к PII и NtcA, уровни 2-OG также модулируют активность многих регуляторных белков и путей, преимущественно связанных с азотным или углеродным метаболизмом у бактерий, архей, водорослей, растений и животных [1,11,15,21,22, 23]. Несмотря на центральную роль 2-OG и его обширный регуляторный репертуар, нам все еще не хватает инструментов для надлежащего измерения его концентраций и динамики in vivo. Для оценки уровней 2-OG в клеточных экстрактах были разработаны методы ферментативного анализа, масс-спектрометрии или жидкостной хроматографии высокого давления (ВЭЖХ) [4,24,25,26,27,28,29,30].Тем не менее, 2-OG, такой как многие метаболиты, очень быстро превращается и может быть потерян или разложен во время экстракции. Как следствие, измерения концентрации 2-OG невоспроизводимы и недооцениваются [30,31,32]. Кроме того, ни один из этих подходов не позволяет проследить динамику 2-OG. Чтобы обойти эти недостатки, недавно были разработаны биосенсоры 2-OG на основе флуоресцентного резонансного переноса энергии (FRET). FRET основывается на зависимом от расстояния взаимодействии между двумя флуоресцентными молекулами и на передаче возбужденного состояния донорной молекулы акцептору, когда эти две молекулы находятся в непосредственной близости [33,34].Мы сконструировали биосенсор 2-OG, основанный на взаимодействии между белками Anabaena PII и PipX, взаимодействие которых нарушается связыванием 2-OG с PII. Было показано, что соотношение FRET (флуоресценция акцептора YFP / флуоресценция донорного CFP) обратно пропорционально уровням 2-OG in vitro в диапазоне 0,01–10 мМ [35]. Второй биосенсор, основанный на 2-OG-зависимом взаимодействии между Synechococcus elongatus PII и белками N-ацетил-глутаматкиназы (NAGK), измеряет уровни 2-OG в 0.
Как следствие, измерения концентрации 2-OG невоспроизводимы и недооцениваются [30,31,32]. Кроме того, ни один из этих подходов не позволяет проследить динамику 2-OG. Чтобы обойти эти недостатки, недавно были разработаны биосенсоры 2-OG на основе флуоресцентного резонансного переноса энергии (FRET). FRET основывается на зависимом от расстояния взаимодействии между двумя флуоресцентными молекулами и на передаче возбужденного состояния донорной молекулы акцептору, когда эти две молекулы находятся в непосредственной близости [33,34].Мы сконструировали биосенсор 2-OG, основанный на взаимодействии между белками Anabaena PII и PipX, взаимодействие которых нарушается связыванием 2-OG с PII. Было показано, что соотношение FRET (флуоресценция акцептора YFP / флуоресценция донорного CFP) обратно пропорционально уровням 2-OG in vitro в диапазоне 0,01–10 мМ [35]. Второй биосенсор, основанный на 2-OG-зависимом взаимодействии между Synechococcus elongatus PII и белками N-ацетил-глутаматкиназы (NAGK), измеряет уровни 2-OG в 0. Диапазон 01–1 мМ [36]. Хотя такие биосенсоры доказали свою высокую эффективность in vitro или ex vivo, они не смогли измерить вариации 2-OG в живых клетках [35,36,37]. Одним из ограничений является то, что было трудно достичь аналогичных уровней продукции двух слитых белков в E. coli и Anabaena. По этой причине необходимо разработать биосенсоры 2-OG на основе отдельных белков для измерения уровней 2-OG in vitro и in vivo. В этом исследовании мы сообщаем о разработке двух новых биосенсоров, основанных на 2-OG-зависимых конформационных изменениях PII или NtcA.Мы показываем, что они оба могут сообщать об изменениях 2-OG с различным диапазоном концентраций in vitro и in vivo у двух бактерий, принадлежащих к разным типам.
Диапазон 01–1 мМ [36]. Хотя такие биосенсоры доказали свою высокую эффективность in vitro или ex vivo, они не смогли измерить вариации 2-OG в живых клетках [35,36,37]. Одним из ограничений является то, что было трудно достичь аналогичных уровней продукции двух слитых белков в E. coli и Anabaena. По этой причине необходимо разработать биосенсоры 2-OG на основе отдельных белков для измерения уровней 2-OG in vitro и in vivo. В этом исследовании мы сообщаем о разработке двух новых биосенсоров, основанных на 2-OG-зависимых конформационных изменениях PII или NtcA.Мы показываем, что они оба могут сообщать об изменениях 2-OG с различным диапазоном концентраций in vitro и in vivo у двух бактерий, принадлежащих к разным типам.4. Обсуждение
В этой работе мы показываем, что белок PII или домен NtcA, содержащий область связывания 2-OG, может быть использован в качестве монобелковых биосенсоров 2-OG FRET при слиянии как с YFP, так и с CFP / mTurquoise. Две конструкции, названные F-mT_PII и F-mT_NtcA (с mTurquoise на С-конце и YFP на полпути через Т-петлю PII или близко к N-концу в NtcA) были основаны на конформационных изменениях, зависящих от связывания 2-OG. Т-образной петли и С-спирали соответственно.Различные биосенсоры, несущие мутации, отменяющие связывание 2-OG с PII и NtcA, также были разработаны для контроля специфичности.
Т-образной петли и С-спирали соответственно.Различные биосенсоры, несущие мутации, отменяющие связывание 2-OG с PII и NtcA, также были разработаны для контроля специфичности.
 2A, B). Такое изменение аффинности может быть связано с большой разницей между карманами связывания 2-OG и остатками, участвующими в связывании 2-OG в PII и NtcA [10,12].Связывание 2-OG с PII происходит в микромолярном диапазоне для точной настройки способности клетки к азотфиксации в зависимости от баланса углерода / азота [48], тогда как 2-OG связывает NtcA на высоких уровнях во время комбинированной клеточной депривации азота [3]. Эта разница в аффинности и кармане связывания может объяснить, почему 2-метиленпентандиовая кислота (2-MPA), неметаболизируемый аналог 2-OG, способна изменять соотношение FRET F-mT_PII, но не F-mT_NtcA (рис. 2G) [46]. Мы также отметили, что концентрации биосенсора не оказали никакого влияния на измерения уровня 2-OG (рис. 2A, B).Это наблюдение, а также тот факт, что два биосенсора реагируют на разные диапазоны уровней 2-OG, представляют собой преимущество при измерении колебаний 2-OG in vivo у широкого круга организмов. Таким образом, разработанные здесь инструменты могут быть применены ко многим биологическим системам, таким как цианобактерии, бактерии, грибы, растения и животные.
2A, B). Такое изменение аффинности может быть связано с большой разницей между карманами связывания 2-OG и остатками, участвующими в связывании 2-OG в PII и NtcA [10,12].Связывание 2-OG с PII происходит в микромолярном диапазоне для точной настройки способности клетки к азотфиксации в зависимости от баланса углерода / азота [48], тогда как 2-OG связывает NtcA на высоких уровнях во время комбинированной клеточной депривации азота [3]. Эта разница в аффинности и кармане связывания может объяснить, почему 2-метиленпентандиовая кислота (2-MPA), неметаболизируемый аналог 2-OG, способна изменять соотношение FRET F-mT_PII, но не F-mT_NtcA (рис. 2G) [46]. Мы также отметили, что концентрации биосенсора не оказали никакого влияния на измерения уровня 2-OG (рис. 2A, B).Это наблюдение, а также тот факт, что два биосенсора реагируют на разные диапазоны уровней 2-OG, представляют собой преимущество при измерении колебаний 2-OG in vivo у широкого круга организмов. Таким образом, разработанные здесь инструменты могут быть применены ко многим биологическим системам, таким как цианобактерии, бактерии, грибы, растения и животные. Важно отметить, что мы показали, что два биосенсора способны обнаруживать вариации уровней 2-OG в течение нескольких минут (рис. 3A, B). ), что особенно важно при исследовании флуктуаций 2-OG во время покадровых экспериментов или после изменения условий эксперимента.Модулируя концентрации глюкозы, 2-OG и аммония в среде, мы наблюдали, что биосенсор на основе NtcA F-mT_NtcA может обнаруживать колебания 2-OG в популяциях E. coli в зависимости от наличия углерода или азота. Даже если нашей целью является определение вариаций 2-OG in vivo, а не количественная оценка абсолютных концентраций 2-OG, полученные данные можно рассматривать как согласующиеся с предыдущими результатами, доступными в литературе. Измерения ВЭЖХ внутриклеточной концентрации 2-OG показали, что она увеличивается от 0.От 35 до 2,6 мМ после добавления 10 мМ глюкозы в безуглеродную среду M9 [37]. Также было показано, что уровни 2-OG значительно снижаются после добавления 10–200 мМ NH 4 Cl к культуре клеток E. coli, культивируемых в условиях азотного голодания [27].
Важно отметить, что мы показали, что два биосенсора способны обнаруживать вариации уровней 2-OG в течение нескольких минут (рис. 3A, B). ), что особенно важно при исследовании флуктуаций 2-OG во время покадровых экспериментов или после изменения условий эксперимента.Модулируя концентрации глюкозы, 2-OG и аммония в среде, мы наблюдали, что биосенсор на основе NtcA F-mT_NtcA может обнаруживать колебания 2-OG в популяциях E. coli в зависимости от наличия углерода или азота. Даже если нашей целью является определение вариаций 2-OG in vivo, а не количественная оценка абсолютных концентраций 2-OG, полученные данные можно рассматривать как согласующиеся с предыдущими результатами, доступными в литературе. Измерения ВЭЖХ внутриклеточной концентрации 2-OG показали, что она увеличивается от 0.От 35 до 2,6 мМ после добавления 10 мМ глюкозы в безуглеродную среду M9 [37]. Также было показано, что уровни 2-OG значительно снижаются после добавления 10–200 мМ NH 4 Cl к культуре клеток E. coli, культивируемых в условиях азотного голодания [27]. Напротив, E. coli, культивированная в присутствии высокого уровня аммония (10 мМ NH 4 Cl), показывала увеличение внутриклеточного 2-OG с 0,5 до 2,5 мМ в условиях азотного голодания [30]. Что касается биосенсора на основе PII, было обнаружено, что он обнаруживает уровни 2-OG в физиологическом диапазоне (10 -2 -10 -5 M) in vitro (рис. 2A).Мы также показали, что эффективно измерять уровни 2-OG in vivo в клетках Anabaena (рис. 3C, D). И здесь наши данные согласуются с ранее опубликованными результатами с использованием клеточных экстрактов [3]. Этот биосенсор, вероятно, будет мощным инструментом для точного измерения вариаций 2-OG на разных стадиях дифференцировки гетероцист. Однако PII является высококонсервативным белком в бактериях и растениях, который контролирует экспрессию и активность многих ферментов, таких как глутаминсинтетаза, ацетил-CoA-карбоксилаза или нитрогеназа [14,49,50,51,52,53,54, 55], биосенсор 2-OG на основе PII, вероятно, не самая подходящая конструкция для измерения колебаний 2-OG у большинства живых организмов, поскольку его избыточное производство может влиять на регуляторные каскады и клеточную активность, не влияя на уровни 2-OG в клетках скорее путем титрования партнеров GlnB / PII.
Напротив, E. coli, культивированная в присутствии высокого уровня аммония (10 мМ NH 4 Cl), показывала увеличение внутриклеточного 2-OG с 0,5 до 2,5 мМ в условиях азотного голодания [30]. Что касается биосенсора на основе PII, было обнаружено, что он обнаруживает уровни 2-OG в физиологическом диапазоне (10 -2 -10 -5 M) in vitro (рис. 2A).Мы также показали, что эффективно измерять уровни 2-OG in vivo в клетках Anabaena (рис. 3C, D). И здесь наши данные согласуются с ранее опубликованными результатами с использованием клеточных экстрактов [3]. Этот биосенсор, вероятно, будет мощным инструментом для точного измерения вариаций 2-OG на разных стадиях дифференцировки гетероцист. Однако PII является высококонсервативным белком в бактериях и растениях, который контролирует экспрессию и активность многих ферментов, таких как глутаминсинтетаза, ацетил-CoA-карбоксилаза или нитрогеназа [14,49,50,51,52,53,54, 55], биосенсор 2-OG на основе PII, вероятно, не самая подходящая конструкция для измерения колебаний 2-OG у большинства живых организмов, поскольку его избыточное производство может влиять на регуляторные каскады и клеточную активность, не влияя на уровни 2-OG в клетках скорее путем титрования партнеров GlnB / PII. Напротив, регулятор NtcA обнаруживается только в пределах типа цианобактерий. Кроме того, чтобы избежать перекрестных помех со стороны других организмов, сконструированный нами биосенсор на основе NtcA лишен ДНК-связывающего домена и, следовательно, потерял способность контролировать экспрессию генов в Anabaena. Таким образом, этот биосенсор наиболее подходит для изучения колебаний 2-OG у большинства организмов. Эти биосенсоры представляют собой первый шаг к разработке флуоресцентных сенсорных систем для 2-OG, способных измерять разницу концентраций этой универсальной сигнальной молекулы в отдельных клетках.Два других биосенсора 2-OG (PII-TC3 и PII-TC3-R9P), основанные на скелете PII Synechococcus elongatus PCC 7942, были недавно описаны [56]. Эти биосенсоры 2-OG также были сконструированы на основе конформационного изменения этой петли для измерения изменения FRET. Белки PII из Anabaena и S. elongatus имеют одинаковый размер (112 аминокислот) и высокое сходство последовательностей. Однако структуры этих биосенсоров (PII-TC3 и PII-TC3-R9P) и биосенсоры, описанные в нашем исследовании, не идентичны.
Напротив, регулятор NtcA обнаруживается только в пределах типа цианобактерий. Кроме того, чтобы избежать перекрестных помех со стороны других организмов, сконструированный нами биосенсор на основе NtcA лишен ДНК-связывающего домена и, следовательно, потерял способность контролировать экспрессию генов в Anabaena. Таким образом, этот биосенсор наиболее подходит для изучения колебаний 2-OG у большинства организмов. Эти биосенсоры представляют собой первый шаг к разработке флуоресцентных сенсорных систем для 2-OG, способных измерять разницу концентраций этой универсальной сигнальной молекулы в отдельных клетках.Два других биосенсора 2-OG (PII-TC3 и PII-TC3-R9P), основанные на скелете PII Synechococcus elongatus PCC 7942, были недавно описаны [56]. Эти биосенсоры 2-OG также были сконструированы на основе конформационного изменения этой петли для измерения изменения FRET. Белки PII из Anabaena и S. elongatus имеют одинаковый размер (112 аминокислот) и высокое сходство последовательностей. Однако структуры этих биосенсоров (PII-TC3 и PII-TC3-R9P) и биосенсоры, описанные в нашем исследовании, не идентичны. Действительно, в случае биосенсоров Synechococcus донор флуорофора вставлен в Т-петлю, тогда как в случае биосенсора Anabaena он является флуорофором-акцептором. Более того, акцептор сливается с С-концом PII через аффинную метку к стрептавидину из 12 аминокислот, и используемые флуорофоры различны. Lüddecke et al. Объединили PII с циркулярно переставленными mCerulean и Venus FP вместо mTurquoise и YFP для F-mT_PII [56]. Наиболее разительное различие между двумя системами заключается в том, что флуорофор не вставляется в точно такое же положение в Т-образной петле.В случае биосенсоров на основе Synechococcus PII последовательность дикого типа дает слишком чувствительный ответ. Следовательно, в последовательность PII была введена замена, чтобы получить соответствующий биосенсор. В случае описанного здесь биосенсора использованная последовательность PII дикого типа оказалась совместимой с измерением физиологических концентраций. Эти структурные различия между двумя биосенсорами могут объяснить тот факт, что две системы реагируют на разные концентрации 2-OG.
Действительно, в случае биосенсоров Synechococcus донор флуорофора вставлен в Т-петлю, тогда как в случае биосенсора Anabaena он является флуорофором-акцептором. Более того, акцептор сливается с С-концом PII через аффинную метку к стрептавидину из 12 аминокислот, и используемые флуорофоры различны. Lüddecke et al. Объединили PII с циркулярно переставленными mCerulean и Venus FP вместо mTurquoise и YFP для F-mT_PII [56]. Наиболее разительное различие между двумя системами заключается в том, что флуорофор не вставляется в точно такое же положение в Т-образной петле.В случае биосенсоров на основе Synechococcus PII последовательность дикого типа дает слишком чувствительный ответ. Следовательно, в последовательность PII была введена замена, чтобы получить соответствующий биосенсор. В случае описанного здесь биосенсора использованная последовательность PII дикого типа оказалась совместимой с измерением физиологических концентраций. Эти структурные различия между двумя биосенсорами могут объяснить тот факт, что две системы реагируют на разные концентрации 2-OG.
Тем не менее, биосенсоры F-mT_PII и F-mT_NtcA должны быть протестированы, чтобы определить наилучшую конструкцию для исследуемого организма. Для дальнейшего улучшения способности биосенсоров измерять динамику изменений 2-OG in vivo в течение продолжительных периодов времени следует учитывать несколько параметров. Во-первых, как и для многих флуорофоров, повторяющееся возбуждение флуоресценции приводит к фотообесцвечиванию, которое задерживает или затрудняет любое покадровое измерение. Мы уже заменили донорный флуорофор CFP на mTurquoise, который, как было показано, обеспечивает более высокую яркость, но менее подвержен фотообесцвечиванию.Будущие улучшения могут включать переключение YFP на более подходящий флуорофор. Во-вторых, измерения FRET in vivo должны мобилизовать больший набор контролей в виде скелетов биосенсоров, слитых с одним флуорофором, чтобы модулировать данные в зависимости от утечки флуоресценции и гетерогенных уровней продукции от одной клетки к другой. Тем не менее, измерение FRET также можно анализировать с помощью альтернативных методов. Здесь мы определили соотношение FRET, измеряя интенсивность флуоресценции донора и акцептора.Технология FRET на основе FLIM (флуоресцентная микроскопия с визуализацией на протяжении всей жизни) также может сообщать о FRET. Эти измерения сосредоточены на изменении срока службы донора, что предотвращает некоторые недостатки измерения FRET на основе интенсивности, такие как перекрестное загрязнение сигнала и фотообесцвечивание. Каждый флуорофор имеет собственное характерное время жизни, которое зависит от его молекулярного окружения, но не зависит от его клеточной концентрации. MTurquoise – это флуорофор, который имеет лучшую производительность по яркости, но, прежде всего, два срока службы, близкие к 3.7 нс и 3,8 нс, которые можно рассматривать как одно время жизни, в отличие от CFP, который имеет два более отдаленных времени жизни (2,3 и 3,0 нс). Переключение донорского флуорофора CFP на mTurquoise позволяет обойти сложную проблему обеспечения стабильной и однородной продукции биосенсоров и позволяет измерять уровни 2-OG in vivo на клеточном уровне.
Здесь мы определили соотношение FRET, измеряя интенсивность флуоресценции донора и акцептора.Технология FRET на основе FLIM (флуоресцентная микроскопия с визуализацией на протяжении всей жизни) также может сообщать о FRET. Эти измерения сосредоточены на изменении срока службы донора, что предотвращает некоторые недостатки измерения FRET на основе интенсивности, такие как перекрестное загрязнение сигнала и фотообесцвечивание. Каждый флуорофор имеет собственное характерное время жизни, которое зависит от его молекулярного окружения, но не зависит от его клеточной концентрации. MTurquoise – это флуорофор, который имеет лучшую производительность по яркости, но, прежде всего, два срока службы, близкие к 3.7 нс и 3,8 нс, которые можно рассматривать как одно время жизни, в отличие от CFP, который имеет два более отдаленных времени жизни (2,3 и 3,0 нс). Переключение донорского флуорофора CFP на mTurquoise позволяет обойти сложную проблему обеспечения стабильной и однородной продукции биосенсоров и позволяет измерять уровни 2-OG in vivo на клеточном уровне. Однако подход FLIM может быть альтернативным или дополнительным методом измерения FRET.
Однако подход FLIM может быть альтернативным или дополнительным методом измерения FRET.
Избыточное производство флаводиирона Flv3B увеличивает фотобиологическое производство водорода азотфиксирующими цианобактериями Nostoc PCC 7120 | Фабрики микробных клеток

Houchins JP, Burris RH. Светлые и темные реакции поглощения гидрогеназы в anabaena 7120. Plant Physiol. 1981; 68: 712–6.
CAS Статья Google Scholar
Пугиони В., Темпель С., Латифи А. Распределение гидрогеназ в цианобактериях: геномный обзор всего филума.Фронт Жене. 2016; 7: 223.
Артикул Google Scholar
Таманьини П., Лейтао Е., Оливейра П., Феррейра Д., Пинто Ф., Харрис Д. Д., Хейдорн Т., Линдблад П. Цианобактериальные гидрогеназы: разнообразие, регулирование и применение. FEMS Microbiol Rev.2007; 31: 692-720.
CAS Статья Google Scholar
Cournac L, Guedeney G, Peltier G, Vignais PM. Устойчивая фотоэволюция молекулярного водорода у мутанта Synechocystis sp.штамм PCC 6803, дефицитный по комплексу НАДФН-дегидрогеназа I типа. J Bacteriol. 2004; 186: 1737–46.
CAS Статья Google Scholar
Kiss E, Kos PB, Vass I. Регуляция транскрипции двунаправленной гидрогеназы в цианобактериях Synechocystis 6803. J. Biotechnol. 2009; 142: 31–7.
CAS Статья Google Scholar
Dutheil J, Saenkham P, Sakr S, Leplat C, Ortega-Ramos M, Bottin H, Cournac L, Cassier-Chauvat C, Chauvat F.Авторепрессор AbrB2, экспрессируемый атипичным промотором, репрессирует оперон гидрогеназы, регулируя выработку водорода в штамме Synechocystis PCC6803. J Bacteriol. 2012; 194: 5423–33.
CAS Статья Google Scholar
Gutekunst K, Phunpruch S, Schwarz C, Schuchardt S, Schulz-Friedrich R., Appel J. LexA регулирует двунаправленную гидрогеназу в цианобактериях Synechocystis sp.PCC 6803 как активатор транскрипции. Mol Microbiol. 2005. 58: 810–23.
CAS Статья Google Scholar
Oliveira P, Lindblad P. LexA, регулятор транскрипции, связывающийся в промоторной области двунаправленной гидрогеназы в цианобактериях Synechocystis sp. PCC 6803. FEMS Microbiol Lett. 2005. 251: 59–66.
CAS Статья Google Scholar
Ханна Н., Линдблад П. Кианобактериальные гидрогеназы и водородный метаболизм: недавний прогресс и перспективы на будущее. Int J Mol Sci. 2015; 16: 10537–61.
CAS Статья Google Scholar
Аллахвердиева Ю., Суорса М., Тикканен М., Аро Э.М. Фотозащита фотосистем при колебаниях интенсивности света. J Exp Bot. 2015; 66: 2427–36.
CAS Статья Google Scholar
Хельман Ю., Чернов Д., Рейнхольд Л., Шибата М., Огава Т., Шварц Р., Охад И., Каплан А. Гены, кодирующие флавопротеины A-типа, необходимы для фотовосстановления O 2 в цианобактериях. Curr Biol. 2003. 13: 230–5.
Гены, кодирующие флавопротеины A-типа, необходимы для фотовосстановления O 2 в цианобактериях. Curr Biol. 2003. 13: 230–5.
CAS Статья Google Scholar
Аллахвердиева Ю., Мустила Н., Ермакова М., Берсанини Л., Ришо П., Айлани Г., Батчикова Н., Курнак Л., Аро Е.М. Белки Flavodiiron Flv1 и Flv3 способствуют росту цианобактерий и фотосинтезу при колебаниях света.Proc Natl Acad Sci USA. 2013; 110: 4111–6.
CAS Статья Google Scholar
Ермакова М., Батчикова Н., Аллахвердиева Ю., Аро Э.М. Новые гетероцист-специфичные белки флаводиирона в Anabaena sp. PCC 7120. FEBS Lett. 2013; 587: 82–7.
CAS Статья Google Scholar
Ермакова М., Батчикова Н., Ришо П., Лейно Х., Косоуров С., Исоярви Дж., Пельтье Г., Флорес Э., Курнак Л., Аллахвердиева Ю., Аро Е.М.Гетероцист-специфичный белок Flv3B флаводиирон обеспечивает кислородный диазотрофный рост нитчатой цианобактерии Anabaena sp. PCC 7120. Proc Natl Acad Sci USA. 2014; 111: 11205–10.
PCC 7120. Proc Natl Acad Sci USA. 2014; 111: 11205–10.
CAS Статья Google Scholar
Avilan L, Roumezi B, Risoul V, Bernard CS, Kpebe A, Belhadjhassine M, Rousset M, Brugna M, Latifi A. Фототрофное производство водорода из клостридиальной гидрогеназы [FeFe], экспрессируемой в гетероцитах цианобактерий. Nostoc PCC 7120.Appl Microbiol Biotechnol. 2018; 102: 5775–83.
CAS Статья Google Scholar
Митчке Дж., Виоке А., Хаас Ф., Хесс В. Р., Муро-Пастор А. М.. Динамика выбора сайта старта транскрипции при дифференцировке клеток под действием азотного стресса у Anabaena sp. PCC7120. Proc Natl Acad Sci USA. 2011; 108: 20130–5.
CAS Статья Google Scholar
Jones KM, Buikema WJ, Haselkorn R. Гетероцит-специфическая экспрессия patB , гена, необходимого для фиксации азота в Anabaena sp. штамм PCC 7120. J Bacteriol. 2003; 185: 2306–14.
штамм PCC 7120. J Bacteriol. 2003; 185: 2306–14.
CAS Статья Google Scholar
Chaux F, Burlacot A, Mekhalfi M, Auroy P, Blangy S, Richaud P, Peltier G. Белки флаводиирона способствуют быстрому и временному фотовосстановлению O 2 при хламидомонаде.Plant Physiol. 2017; 174: 1825–36.
CAS Статья Google Scholar
Nierzwicki-Bauer SA, Curtis SE, Haselkorn R. Котранскрипция генов, кодирующих малые и большие субъединицы рибулозо-1,5-бисфосфаткарбоксилазы в цианобактериях Anabaena 7120. Proc Natl Acad Sci. 1984. 81: 5961–5.
CAS Статья Google Scholar
Масукава Х., Мочимару М., Сакураи Х. Нарушение гена поглощения гидрогеназы, но не гена двунаправленной гидрогеназы, приводит к усиленному фотобиологическому производству водорода азотфиксирующими цианобактериями Anabaena sp. PCC 7120. Appl Microbiol Biotechnol. 2002; 58: 618–24.
PCC 7120. Appl Microbiol Biotechnol. 2002; 58: 618–24.
CAS Статья Google Scholar
Шохольм Дж., Оливейра П., Линдблад П. Транскрипция и регуляция двунаправленной гидрогеназы в цианобактериях Nostoc sp.штамм PCC 7120. Appl Environ Microbiol. 2007. 73: 5435–46.
Артикул Google Scholar
Нюберг М., Хейдорн Т., Линдблад П. Производство водорода сконструированным штаммом цианобактерий Nostoc PCC 7120 DeltahupW исследовали в системе фотобиореактора с плоской панелью. J Biotechnol. 2015; 215: 35–43.
CAS Статья Google Scholar
Лиран О., Семятич Р., Милрад И., Эйленберг Х., Вайнер И., Якоби И.Микрооксичные ниши в тилакоидной строме выращенных на воздухе Chlamydomonas reinhardtii Защищают [FeFe] -гидрогеназу и поддерживают производство водорода в полностью аэробной среде. Plant Physiol. 2016; 172: 264–71.
Plant Physiol. 2016; 172: 264–71.
CAS Статья Google Scholar
Паттерсон-Фортин Л.М., Колвин К.Р., Отттрим Г.В. Белок, родственный LexA, регулирует чувствительную к окислению-восстановлению экспрессию цианобактериальной РНК-геликазы crhR. Nucleic Acids Res.2006; 34: 3446–54.
CAS Статья Google Scholar
Houchins JP, Burris RH. Сравнительная характеристика двух различных гидрогеназ из Anabaena sp. штамм 7120. J Bacteriol. 1981; 146: 215–21.
CAS Статья Google Scholar
Ян И, Хуанг XZ, Ван Л., Ризул В, Чжан С.К., Чен В.Л. Фенотипические вариации, вызванные вариациями относительного числа копий плазмид на основе pDU1, экспрессирующих домен GAF Pkn41 или Pkn42 у Anabaena sp.PCC 7120. Res Microbiol. 2013. 164: 127–35.
CAS Статья Google Scholar

Цай Ю.П., Волк С.П. Использование условно летального гена в Anabaena sp. штамм PCC 7120 для отбора двойных рекомбинантов и улавливания инсерционных последовательностей. J Bacteriol. 1990; 172: 3138–45.
CAS Статья Google Scholar
Ричи Р.Дж. Согласованные наборы спектрофотометрических уравнений хлорофилла для растворителей ацетона, метанола и этанола.Photosynth Res. 2006; 89: 27–41.
CAS Статья Google Scholar
Dron A, Rabouille S, Claquin P, Talec A, Raimbault V, Sciandra A. Продолжительность фотопериода определяет временную оркестровку клеточного цикла и углеродно-азотного метаболизма у Crocosphaera watsonii . Environ Microbiol. 2013; 15: 3292–304.
CAS Статья Google Scholar
Wolk CP, Cai Y, Cardemil L, Flores E, Hohn B, Murry M, Schmetterer G, Schrautemeier B, Wilson R. Выделение и комплементация мутантов Anabaena sp. штамм PCC 7120 не может расти в аэробных условиях на диазоте. J Bacteriol. 1988; 170: 1239–44.
Выделение и комплементация мутантов Anabaena sp. штамм PCC 7120 не может расти в аэробных условиях на диазоте. J Bacteriol. 1988; 170: 1239–44.
CAS Статья Google Scholar
Тканевая культура-специфическая экспрессия естественно встречающейся табачной нечувствительной к обратной связи антранилатсинтазы
- Copyright © 1998 Американское общество физиологов растений
Abstract
кДНК и соответствующая промоторная область для природного, нечувствительного к обратной связи гена α-субъединицы антранилатсинтазы (AS), ASA2 , была выделена из невыделенного, но устойчивого к 5-метилтриптофану (5MT r ) , линия клеток табака ( Nicotiana tabacum ) (AB15–12-1).КДНК ASA2 содержит предполагаемую последовательность транзитного пептида, и гибридизация по Саузерну показывает, что в геноме табака присутствует более одной близкородственной последовательности. КДНК ASA2 дополняла нонсенс-мутант trpE штамм Escherichia coli , обеспечивая рост на 300 мкм 5MT-содержащей минимальной среде без триптофана, и клеточные экстракты содержали нечувствительную к обратной связи активность AS. 5MT r был утерян, когда штамм E. coli трансформировали сайт-направленным мутантом ASA2 (фенилаланин-107-аргинин-108 → серин-107-глутамин-108).Идентичные нуклеотидные последовательности, кодирующие область фенилаланин-107-аргинин-108, были обнаружены в амплифицированных полимеразной цепной реакцией геномных фрагментах 326 пар оснований ASA2 дикого типа (5-метилтриптофан-чувствительный [5MT s ]) табака и вид-прародитель. Высокий уровень транскрипционной экспрессии ASA2 был обнаружен только в культивируемых клетках 5MT r , но не в клетках 5MT s или в растениях. Исследования промоторов показывают, что тканевая специфичность ASA2 контролируется областью промотора между -2252 и -607.
КДНК ASA2 дополняла нонсенс-мутант trpE штамм Escherichia coli , обеспечивая рост на 300 мкм 5MT-содержащей минимальной среде без триптофана, и клеточные экстракты содержали нечувствительную к обратной связи активность AS. 5MT r был утерян, когда штамм E. coli трансформировали сайт-направленным мутантом ASA2 (фенилаланин-107-аргинин-108 → серин-107-глутамин-108).Идентичные нуклеотидные последовательности, кодирующие область фенилаланин-107-аргинин-108, были обнаружены в амплифицированных полимеразной цепной реакцией геномных фрагментах 326 пар оснований ASA2 дикого типа (5-метилтриптофан-чувствительный [5MT s ]) табака и вид-прародитель. Высокий уровень транскрипционной экспрессии ASA2 был обнаружен только в культивируемых клетках 5MT r , но не в клетках 5MT s или в растениях. Исследования промоторов показывают, что тканевая специфичность ASA2 контролируется областью промотора между -2252 и -607. Поскольку промоторные последовательности ASA2 существенно не отличаются в линиях 5MT r и 5MT s , повышенные уровни мРНК ASA2 в линиях 5MT r , скорее всего, связаны с изменениями в регуляторном гене, влияющем на Экспрессия ASA2 .
Поскольку промоторные последовательности ASA2 существенно не отличаются в линиях 5MT r и 5MT s , повышенные уровни мРНК ASA2 в линиях 5MT r , скорее всего, связаны с изменениями в регуляторном гене, влияющем на Экспрессия ASA2 .
Многие ароматические соединения синтезируются шикиматным путем у высших растений (Haslam, 1993; Herrmann, 1995). Хоризмат находится в точке ветвления этого пути и является последним общим предшественником многих из этих соединений.AS катализирует первую детерминированную реакцию в ветви биосинтеза Trp, превращая хоризмат в антранилат, и по обратной связи ингибируется конечным продуктом, Trp (Haslam, 1993; Romero et al., 1995). На то, что AS является контрольной точкой в ветви Trp в растительных клетках, указывают (а) исследования питания промежуточного звена (Widholm, 1974), (b) уровни активности ферментов, (c) ингибирование активности соответствующих ферментов по принципу обратной связи (Singh and Widholm, 1974), и (d) отбор на устойчивость к 5MT, в результате которого были получены линии с измененным AS, подавленным по обратной связи, и более высоким свободным Trp (Widholm, 1972a, 1972b).
Системы культивирования растительных клеток были полезны для изучения регуляторных механизмов биосинтеза аминокислот (Widholm, 1972a, 1972b). Отбор клеток, устойчивых к аналогам аминокислот, таким как 5MT, может дать линии с повышенным количеством конечного продукта, Trp, из-за изменения аллостерического регуляторного фермента, вызывающего меньшее ингибирование фермента аналогом и Trp (Widholm, 1972a, 1972b ). Нечувствительный к обратной связи AS был обнаружен в отобранной 5MT моркови ( Daucus carota ; Brotherton et al., 1986), Datura innoxia (Ranch et al., 1983), картофель ( Solanum tuberosum ; Carlson and Widholm, 1978) и табак ( Nicotiana tabacum ; Brotherton et al., 1986). Клеточная линия табака 5MT r (TX2-4) содержала Trp-нечувствительный AS и более высокие уровни свободного Trp, чем клетки дикого типа. Аналогичные результаты были получены с 5MT-отобранными культивированными клетками картофеля (Carlson and Widholm, 1978). Эти характеристики не были обнаружены у регенерированных растений табака, но были снова восстановлены в культивируемых клетках, индуцированных из листьев регенерантов (Brotherton et al. , 1986). Эти результаты предполагают, что экспрессия этого измененного AS регулируется тканеспецифическим образом и что отбор на 5MT r дает линии с повышенным количеством нечувствительной к обратной связи формы AS, которая экспрессируется в культивируемых клетках, но не в растениях.
, 1986). Эти результаты предполагают, что экспрессия этого измененного AS регулируется тканеспецифическим образом и что отбор на 5MT r дает линии с повышенным количеством нечувствительной к обратной связи формы AS, которая экспрессируется в культивируемых клетках, но не в растениях.
Недавно AS был очищен из культивируемых растительных клеток или растительных тканей (Poulsen et al., 1993; Bohlmann et al., 1995; Romero and Roberts, 1996), и было обнаружено, что он имеет состав субъединиц, подобный AS некоторых микробов, т.е. .е. неидентичные большие (α, компонент I) и малые (β, компонент II) субъединицы (Zalkin et al., 1984; Yanofsky, Crawford, 1987; Crawford, 1989). Гены AS, кодирующие α-субъединицу (Niyogi and Fink, 1992; Bohlmann et al., 1995) и β-субъединицу (Niyogi et al., 1993), были клонированы из Arabidopsis и Ruta graveolens . Два гена AS, кодирующие α-субъединицу фермента, клонированного из Arabidopsis и R. graveolens , были обозначены ASA1 / ASA2 и AS α1 / AS α2 соответственно (Niyogi and Fink, 1992; Bohlmann et al. , 1995). Эти гены AS имеют сходные паттерны экспрессии генов, поскольку экспрессия α1 ASA1 и AS индуцируется раной и / или элиситорной обработкой, тогда как гены ASA2 и AS α2 экспрессируются конститутивно на низких уровнях (Niyogi и Fink, 1992; Bohlmann et al., 1995).
, 1995). Эти гены AS имеют сходные паттерны экспрессии генов, поскольку экспрессия α1 ASA1 и AS индуцируется раной и / или элиситорной обработкой, тогда как гены ASA2 и AS α2 экспрессируются конститутивно на низких уровнях (Niyogi и Fink, 1992; Bohlmann et al., 1995).
Две группы описали мутантный ген AS Arabidopsis, кодирующий нечувствительную к обратной связи α-субъединицу фермента (Kreps et al., 1996; Li and Last, 1996). Секвенирование этих мутантных генов AS показало, что одна аминокислота, Asp-341, была заменена на Asn в обоих случаях.Эти мутантные гены AS экспрессируются как в культивируемых клетках, так и в листьях, с немного более высокой экспрессией в культивируемых клетках (Kreps et al., 1996). В отличие от этих мутантов Arabidopsis, ген AS α1 из R. graveolens , который кодирует нечувствительную к обратной связи α-субъединицу, был выделен из молодых побегов дикого типа (Bohlmann et al., 1996). На основе сравнения последовательностей было сделано предположение, что простая аминокислотная замена в гене AS α1, Arg-138 на Gln-138, может вызвать нечувствительность к обратной связи. Остатки, потенциально влияющие на ингибирование обратной связи у высших растений, не совпадают с остатками бактерий (Matsui et al., 1987; Caliguri and Bauerle, 1991) и дрожжей (Graf et al., 1993), но измененные аминокислоты расположены внутри или очень близки к консервативным мотивам Leu-Leu-Glu-Ser-X 10 -Ser и Asn-Pro-Ser-Pro-Tyr-Met, которые важны для ингибирования обратной связи (Matsui et al., 1987; Caliguri and Bauerle , 1991; Граф и др., 1993).
Остатки, потенциально влияющие на ингибирование обратной связи у высших растений, не совпадают с остатками бактерий (Matsui et al., 1987; Caliguri and Bauerle, 1991) и дрожжей (Graf et al., 1993), но измененные аминокислоты расположены внутри или очень близки к консервативным мотивам Leu-Leu-Glu-Ser-X 10 -Ser и Asn-Pro-Ser-Pro-Tyr-Met, которые важны для ингибирования обратной связи (Matsui et al., 1987; Caliguri and Bauerle , 1991; Граф и др., 1993).
Растения табака, регенерированные из суспензионных культур 5MT r , описанных Brotherton et al.(1986) не были плодородными; поэтому отбор повторяли с новыми культурами для получения фертильных растений (S. Schechter и J.M. Widholm, неопубликованные данные). Одно из девяти растений, регенерированных из неотобранной контрольной культуры, продуцировало суспензионную культуру из листа, который был 5MT r . Эта устойчивость была унаследована потомством, и изучаемая здесь клеточная линия AB15–12-1 была получена от растения пятого поколения.
МАТЕРИАЛЫ И МЕТОДЫ
Суспензионные культуры поддерживали еженедельным переносом в 50 мл жидкой среды MX (базальная среда MS [Murashige and Skoog, 1962] с 1.8 мкм 2,4-Д). Клеточная линия 5MT r AB15–12-1 произошла от растения табак ( Nicotiana tabacum cv Xanthi), регенерированного из невыбранных культивируемых в суспензии клеток. Первоначально девять растений были регенерированы путем помещения неотобранных клеток на отвержденную агаром среду MS с 5,7 мкм IAA и 2 мкм кинетином. Каллус и последующие суспензионные культуры были инициированы с листьев этих растений. Одно растение продуцировало суспензионную культуру, которая показала устойчивость к 5MT, когда 0,5 г свежей массы клеток инкубировали в течение 9–12 дней в жидкой среде MX, содержащей 46 и 137 мкм5MT; другие клетки дикого типа не росли в той же среде.Поскольку это растение обладало мужской стерильностью, его опыляли пыльцой дикого типа, и полученное потомство самоопылялось четыре раза, чтобы получить AB15-12-1. Дополнительные суспензионные культуры клеток табака были впоследствии инициированы из проростков AB15-12-1 и поддерживались на среде, содержащей MX или 5MT, как указано.
Каллус и последующие суспензионные культуры были инициированы с листьев этих растений. Одно растение продуцировало суспензионную культуру, которая показала устойчивость к 5MT, когда 0,5 г свежей массы клеток инкубировали в течение 9–12 дней в жидкой среде MX, содержащей 46 и 137 мкм5MT; другие клетки дикого типа не росли в той же среде.Поскольку это растение обладало мужской стерильностью, его опыляли пыльцой дикого типа, и полученное потомство самоопылялось четыре раза, чтобы получить AB15-12-1. Дополнительные суспензионные культуры клеток табака были впоследствии инициированы из проростков AB15-12-1 и поддерживались на среде, содержащей MX или 5MT, как указано.
Анализ нуклеиновых кислот
Геномную ДНК выделяли из клеток, культивируемых в суспензии в возрасте 1 недели, с использованием процедур Dellaporta et al. (1983) и очищены центрифугированием в градиенте CsCl (Ausubel et al., 1989; Sambrook et al., 1989).
Суммарная РНК была получена с использованием метода экстракции фенолом (Wang et al. , 1994) из однонедельных культивируемых в суспензии клеток, зрелых семян и листьев, корней и стеблей, собранных из 3-недельных культур побегов. на базальной среде MS, затвердевшей с 0,8% агаром.
, 1994) из однонедельных культивируемых в суспензии клеток, зрелых семян и листьев, корней и стеблей, собранных из 3-недельных культур побегов. на базальной среде MS, затвердевшей с 0,8% агаром.
ДНК и РНК наносили на нейлоновую мембрану (Hybond N + , Amersham) в соответствии с общим методом капиллярного переноса (Sambrook et al., 1989). Клон кДНК Arabidopsis thaliana ASA1 (pKN41) (полученный от K.Niyogi [Niyogi and Fink, 1992]) или другие клоны кДНК AS табака, полученные в этой работе, использовали в качестве зонда после мечения с помощью системы мечения ДНК Megaprime (Amersham) с [α- 32 P] dCTP (3000 Ки / ммоль). Южную и северную гибридизацию проводили при 42 ° C с раствором для гибридизации (50% формамид, 5 × SSPE, 5 × раствор Денхардта [Sambrook et al., 1989], 0,1% SDS и 100 мкг / мл ДНК спермы лосося). Мембраны дважды промывали при высокой жесткости при комнатной температуре 2 × SSC и 0.5% SDS в течение 20 минут каждый раз и при 65 ° C с 0,1 × SSC и 0,1% SDS до исчезновения фонового сигнала, а затем подвергали рентгеновской пленке в течение ночи с усиливающим экраном.
Клонирование кДНК AS
кДНК AS табака выделяли с использованием 5 ‘и 3′ RACE (GIBCO-BRL). Праймеры 1 и 2 для клонирования 5’-конца кДНК ASA2 были сконструированы на основе последовательности геномного клона Arabidopsis ASA1 (номер доступа в GenBank M92353). Последовательности праймеров, используемых для клонирования, перечислены в таблице I.Для 5′-RACE кДНК первой цепи синтезировали с антисмысловым праймером 1 в положениях нуклеотидов с 5403 по 5382 геномного клона Arabidopsis ASA1 . 5′-конец кДНК первой цепи был олиго-dA-хвостом (200 мкм dATP) с использованием концевой дезоксинуклеотидилтрансферазы (0,4 единицы / мкл).
Таблица I.Нуклеотидные последовательности праймеров, используемых для клонирования
Вложенную ПЦР проводили с адаптерным праймером 5′-GGCCACGCGTCGACTAGTAC (T) 17 -3 ‘(GIBCO-BRL) и антисмысловым праймером 2 на нуклеотидах 4430-4409 геномного клона Arabidopsis ASA1 .КДНК первой цепи использовали в качестве матрицы для вложенной ПЦР. Фрагмент (приблизительно 1,1 т.п.н.) амплифицировали, клонировали в вектор pGEM-T и секвенировали. Процедуры, использованные для выделения 3′-конца кДНК ASA2 , были такими же, как и для 5′-RACE, за исключением праймеров и хвоста dATP на 5′-конце кДНК. Праймер 3 был сконструирован на основе последовательности клона кДНК 5 ‘ ASA2 с нуклеотидами от +148 до +168 (аденин ATG считали как +1, поскольку сайт начала транскрипции в данной работе не определялся).Фрагмент размером приблизительно 1,9 т.п.н. амплифицировали и сильно гибридизовали с клоном кДНК 5 ‘ ASA2 в качестве зонда. Этот фрагмент был клонирован в вектор pGEM-T и секвенирован.
Фрагмент (приблизительно 1,1 т.п.н.) амплифицировали, клонировали в вектор pGEM-T и секвенировали. Процедуры, использованные для выделения 3′-конца кДНК ASA2 , были такими же, как и для 5′-RACE, за исключением праймеров и хвоста dATP на 5′-конце кДНК. Праймер 3 был сконструирован на основе последовательности клона кДНК 5 ‘ ASA2 с нуклеотидами от +148 до +168 (аденин ATG считали как +1, поскольку сайт начала транскрипции в данной работе не определялся).Фрагмент размером приблизительно 1,9 т.п.н. амплифицировали и сильно гибридизовали с клоном кДНК 5 ‘ ASA2 в качестве зонда. Этот фрагмент был клонирован в вектор pGEM-T и секвенирован.
Анализ выравнивания последовательностей двух фрагментов с использованием Clustal (Higgins and Sharp, 1989) показывает, что перекрывающаяся область длиной 828 п.н. идентична. Существует только один сайт Xba I в перекрывающейся области длиной 828 п.н., только один сайт Nsi I в векторе pGEM-T и ни один сайт Nsi I ни в 5′-, ни в 3′-фрагментах. Эти два сайта рестрикционных ферментов использовали для конструирования полноразмерной кДНК ASA2 табака. Секвенирование клонов кДНК было выполнено Лабораторией генной инженерии Иллинойского университета (Урбана-Шампейн) с использованием метода секвенирования дидезоксинуклеотидов.
Эти два сайта рестрикционных ферментов использовали для конструирования полноразмерной кДНК ASA2 табака. Секвенирование клонов кДНК было выполнено Лабораторией генной инженерии Иллинойского университета (Урбана-Шампейн) с использованием метода секвенирования дидезоксинуклеотидов.
Тесты комплементации и сайт-направленный мутагенез
кДНК ASA2 без предполагаемой последовательности транзитного пептида (аминокислотная последовательность 1-60) была амплифицирована с использованием ДНК-полимеразы Pfu (Stratagene) с праймерами 4 и 5 (таблица I), содержащими Bam HI и Kpn I, соответственно, нависает и лигируется в вектор экспрессии (pQE30, содержащий последовательность, кодирующую 6-His-метку [6xHis, Qiagen, Санта-Кларита, Калифорния]) с последующим перевариванием рестрикционным ферментом.
Сайт-направленный мутагенез выполняли с помощью ПЦР с использованием праймера, содержащего несовпадающие нуклеотидные последовательности, путем замены четырех нуклеотидов (CCT̅GGT̅TT̅TCG̅A → CCC̅GGG̅TC̅TCA̅A) на аминокислотных остатках 105-108. Первые два несовпадающих нуклеотида, Pro-105 и Gly-106, не изменяются. аминокислотный кодон, но создает сайт Sma I. Последние два несовпадающих нуклеотида изменяют Phe-107 и Arg-108 на Ser-107 и Gln-108. Два продукта ПЦР были получены с использованием праймеров 6 и 7 и праймеров 8 и 5 (таблица I).Эти два ПЦР-фрагмента лигировали, расщепляли Sma I и лигировали в рамке считывания в вектор pQE30.
Первые два несовпадающих нуклеотида, Pro-105 и Gly-106, не изменяются. аминокислотный кодон, но создает сайт Sma I. Последние два несовпадающих нуклеотида изменяют Phe-107 и Arg-108 на Ser-107 и Gln-108. Два продукта ПЦР были получены с использованием праймеров 6 и 7 и праймеров 8 и 5 (таблица I).Эти два ПЦР-фрагмента лигировали, расщепляли Sma I и лигировали в рамке считывания в вектор pQE30.
Химерные конструкции трансформировали в мутант trpE Escherichia coli ( trpE5972 , бессмысленный мутант) с использованием трансформации CaCl 2 (Sambrook et al., 1989). Комплементарные штаммы высевали на минимальную среду M9, содержащую 100 мкг / мл ампициллина и 0,1 мМ IPTG, но без Trp. Для теста ингибирования 300 мкМ 5MT добавляли к минимальной среде, описанной ранее.
Экспрессия 6xHis-меченного белка ASA2 в
E. coli Мутант E. coli trpE ( trpE5972 ), трансформированный химерной конструкцией ASA2 с 6xHis-меткой, выращивали в 1 л среды Лурия-Бертани с добавлением 100 мкг мл -1 ампициллина и 0,1 мМ Trp. IPTG (0,1 мм) и PMSF (0,1 мм) добавляли к культуре со средним логарифмом. После 3 ч дальнейшей инкубации при 30 ° C и 150 об / мин клетки собирали центрифугированием, ресуспендировали в 20 мл буфера для экстракции (Bernasconi et al., 1994) с добавлением 0,1 мм PMSF, разрушили с помощью французского пресса (два прохода при 20000 p.s.i.) и центрифугировали для удаления остатков клеток. Супернатант выдерживали в течение ночи на льду, а затем объединяли с 2 объемами насыщенного, комнатной температуры (NH 4 ) 2 SO 4 . Полученный белковый осадок собирали центрифугированием при 4 ° C и ресуспендировали в 10 мл 50 мМ NaH 2 PO 4 , 300 мМ NaCl и 2 мМ DTT, pH 8,0. Меченый 6xHis белок ASA2 был частично очищен после связывания со смолой Ni-нитрилотриуксусной кислоты (Qiagen) и элюирования 100 мМ имидазола в 50 мМ NaH 2 PO 4 , 300 мМ NaCl, 2 мМ DTT и 10% глицерина. , pH 6.0.
IPTG (0,1 мм) и PMSF (0,1 мм) добавляли к культуре со средним логарифмом. После 3 ч дальнейшей инкубации при 30 ° C и 150 об / мин клетки собирали центрифугированием, ресуспендировали в 20 мл буфера для экстракции (Bernasconi et al., 1994) с добавлением 0,1 мм PMSF, разрушили с помощью французского пресса (два прохода при 20000 p.s.i.) и центрифугировали для удаления остатков клеток. Супернатант выдерживали в течение ночи на льду, а затем объединяли с 2 объемами насыщенного, комнатной температуры (NH 4 ) 2 SO 4 . Полученный белковый осадок собирали центрифугированием при 4 ° C и ресуспендировали в 10 мл 50 мМ NaH 2 PO 4 , 300 мМ NaCl и 2 мМ DTT, pH 8,0. Меченый 6xHis белок ASA2 был частично очищен после связывания со смолой Ni-нитрилотриуксусной кислоты (Qiagen) и элюирования 100 мМ имидазола в 50 мМ NaH 2 PO 4 , 300 мМ NaCl, 2 мМ DTT и 10% глицерина. , pH 6.0.
Белок ASA1 Arabidopsis, слитый с глутатионом S -трансферазой, экспрессировали с использованием конструкции pSCI1674, клонированной Bernasconi et al. В E. coli MC1061. (1994). Процедура была такой же, как описано выше, за исключением того, что ресуспендированный осадок (NH 4 ) 2 SO 4 использовали без дополнительной очистки.
В E. coli MC1061. (1994). Процедура была такой же, как описано выше, за исключением того, что ресуспендированный осадок (NH 4 ) 2 SO 4 использовали без дополнительной очистки.
AS Enzyme Assay
Экстракты клетокполучали, как описано Brotherton et al. (1986) из культур клеток в средней логарифмической фазе и использовались в тот же день.Ресуспендированная фракция (NH 4 ) 2 SO 4 была обессолена с использованием Сефадекса G-25.
Активность AS измеряли с использованием метода этилацетатной экстракции, использовавшегося ранее для экстрактов клеток табака (Widholm, 1971), и включения модификаций, описанных Li и Last (1996), за исключением того, что буфер для анализа был 50 мМ Hepes, pH 7,5, 10 ммGln, 2,0 мм MgCl 2 , 0,05 мм Na 2 EDTA, 2,0 мм DTT и 5% глицерин. Реакцию начинали добавлением 50 мкл клеточного экстракта.Концентрации хоризмата и Trp были такими, как указано в отдельных экспериментах. Продукция антранилата была линейной по времени в течение двойного стандартного времени инкубации и концентрации фермента (данные не показаны). Превращение хоризма в антранилат не превышало 1% ни в одном эксперименте. Хоризмат был получен с использованием метода ферментации Гибсона (1970).
Продукция антранилата была линейной по времени в течение двойного стандартного времени инкубации и концентрации фермента (данные не показаны). Превращение хоризма в антранилат не превышало 1% ни в одном эксперименте. Хоризмат был получен с использованием метода ферментации Гибсона (1970).
ASA2 геномных фрагментов, содержащих область Phe-108-Arg-108, были амплифицированы с помощью ПЦР и выделены из трех культивируемых клеточных линий дикого типа (5MT s ): N. tabacum (TXD), Nicotiana tomentosiformis (Nto ) и Nicotiana sylvestris (Ns) из листьев, собранных из культур побегов AB15-12-1, а также из двух клеточных линий табака 5MT s (h25-6, NRMX) и одной 5MT r (NR5MT). Область от -110 до +392 сначала была амплифицирована с использованием двух наборов праймеров на основе кДНК ASA2 и промоторных последовательностей 9 и 10 (таблица I). Затем праймеры 4 и 10 использовали для вложенной ПЦР-амплификации с использованием первого продукта ПЦР в качестве матрицы для получения геномных фрагментов длиной 326 пар оснований (от +181 до +392), которые затем были клонированы в pBluescriptSK-, расщепленные Eco RV, и упорядочены.
Затем праймеры 4 и 10 использовали для вложенной ПЦР-амплификации с использованием первого продукта ПЦР в качестве матрицы для получения геномных фрагментов длиной 326 пар оснований (от +181 до +392), которые затем были клонированы в pBluescriptSK-, расщепленные Eco RV, и упорядочены.
Клонирование промотора AS и конструирование химерных конструкций промотор AS-GUS
Промотор ASA2 был выделен с помощью обратной ПЦР (Ochman et al., 1989). Геномную ДНК AB15–12-1 расщепляли Hin dIII, подвергали циркуляризации ДНК-лигазой Т4 после разбавления до 100 нг / мл дистиллированной деионизированной водой и использовали в качестве матрицы для обратной ПЦР с праймером 3 и антисмысловым праймером 11 на основе последовательность кДНК ASA2 с нуклеотидом от +10 до -11.ПЦР-амплификацию проводили в течение 30 циклов (95 ° C, 1 мин; 50 ° C, 40 с; 72 ° C, 2 мин). Фрагмент ПЦР (приблизительно 2,3 т.п.н.) был обнаружен гибридизацией по Саузерну с кДНК ASA2 в качестве зонда и клонирован в вектор pGEM-T. Секвенирование промотора ASA2 (приблизительно 2,3 т.п.н.) было выполнено в Секции молекулярного анализа и синтеза Фонда Сэмюэля Робертса Нобла (Ардмор, Оклахома) с использованием метода дидезоксинуклеотидного секвенирования.
Секвенирование промотора ASA2 (приблизительно 2,3 т.п.н.) было выполнено в Секции молекулярного анализа и синтеза Фонда Сэмюэля Робертса Нобла (Ардмор, Оклахома) с использованием метода дидезоксинуклеотидного секвенирования.
На основании результатов поиска в базе данных делеции были выполнены с использованием ПЦР-амплификации с семью наборами праймеров следующим образом: антисмысловой праймер 12 со смысловыми праймерами 13, 14, 15 или 16 (таблица I), который амплифицировал 2252-, 1356-, 606- , или фрагменты промотора ASA2 длиной 370 п.н. (включая 5′-UTR) соответственно.Эти продукты ПЦР были слиты с репортерным геном GUS с терминатором NOS. Химерные конструкции были обозначены как 2252, 1356, 606 и 370. Каждый праймер содержит выступающий участок сайта рестрикционного фермента для клонирования. Праймер 12 содержит сайт Sma I, а праймеры 13, 14, 15 и 16 содержат сайт Pst I. Каждый сайт рестрикционного фермента был подчеркнут в последовательностях праймеров (Таблица I). Эти пять фрагментов были клонированы в pBI221, заменив промотор 35S CaMV.
Эти пять фрагментов были клонированы в pBI221, заменив промотор 35S CaMV.
Анализ GUS
Сконструированные плазмидные ДНК были выделены с использованием набора Plasmid Maxi Kit (Qiagen) и биологически перенесены в культивируемые в суспензии табака клетки (AB15-12-1) и листья с использованием пистолета для притока частиц (5 мкг ДНК и 0.5 мг золотых частиц диаметром 1,0 мкм [Bio-Rad] выстрелил при 80 фунтов на квадратный дюйм; Vain et al., 1993). Листья (180–350 мг сырой массы), собранные с культур 3-недельных побегов, помещали на стерильный ватман диаметром 5 см. 1 бумажный фильтровальный диск. Пять миллилитров культивированных в суспензии клеток AB15-12-1 возрастом 2 дня (приблизительно 200 мг свежего веса) собирали на фильтровальной бумаге с использованием вакуума перед бомбардировкой. Образцы переносили с фильтровальной бумагой на твердую среду MX после бомбардировки и инкубировали при 24 ° C под флуоресцентным светом (60 мкЕ м -2 с -1 ) в течение 3 дней. Активность промотора определяли с помощью флуорометрического анализа MUG (Jefferson, 1987; Jefferson et al., 1987). GUS-специфическая активность определялась как пикомоль 4-метилумбеллиферона в час на миллиграмм белка. Концентрацию белка определяли с использованием набора для анализа связывания белкового красителя (Bio-Rad).
Активность промотора определяли с помощью флуорометрического анализа MUG (Jefferson, 1987; Jefferson et al., 1987). GUS-специфическая активность определялась как пикомоль 4-метилумбеллиферона в час на миллиграмм белка. Концентрацию белка определяли с использованием набора для анализа связывания белкового красителя (Bio-Rad).
РЕЗУЛЬТАТЫ
ASA2 Клонирование кДНК и анализ последовательности Мы клонировали полноразмерную кДНК ASA2 (2,16 т.п.н., включая 5 ‘и 3’ UTR) с использованием 5 ‘и 3’ RACE.Нуклеотидная и выведенная аминокислотная последовательности показаны на фиг. 1. Этот ген AS обозначен как ASA2 , поскольку его аминокислотная последовательность на 72% идентична последовательности AS α2 из R. graveolens . Очевидная 5′-UTR была обнаружена на 89 нуклеотидов выше стартового кодона трансляции (ATG) на 5′-конце фрагмента кДНК. Результаты 3 ‘RACE показывают, что полиаденилирование начинается через 205 нуклеотидов ниже стоп-кодона трансляции. Не было обнаружено идеального совпадения с консенсусной эукариотической сигнальной последовательностью поли (A + ) AAUAAA в 3 ‘UTR.
Не было обнаружено идеального совпадения с консенсусной эукариотической сигнальной последовательностью поли (A + ) AAUAAA в 3 ‘UTR.
нуклеотид и предсказанная аминокислотная последовательность кДНК ASA2 . Кодирующая область соответствует аминокислотной последовательности от 1 (ATG) до 616 (TAG). Предполагаемые стартовый и стоп-кодоны трансляции выделены жирным шрифтом. 89 и 205 п.о. области кодирования в восходящем и нисходящем направлениях представляют 5 ‘и 3’ UTR, соответственно.
Клон кДНК ASA2 гибридизовался в условиях высокой жесткости с несколькими фрагментами геномной ДНК, расщепленными девятью рестрикционными ферментами (рис.2А). Три основные полосы примерно 3,3, 4,0 и 24 т.п.н. были обнаружены в Kpn I-переваренной геномной ДНК амфидиплоидных листьев N. tabacum и двух культивируемых клеточных линий, тогда как диплоидные предшественники табака, N. sylvestris и N. tomentosiformis показали разные картины (фиг. 2B). Картина гибридизации N. tabacum была больше похожа на N. tomentosiformis , чем на N. sylvestris , поскольку оба имели полосы примерно на 3.3 и 4,0 кб. Две слабые полосы гибридизуются с клоном кДНК ASA2 в N. sylvestris . Фрагмент размером 24 т.п.н. в N. tabacum демонстрирует очень слабую гибридизацию и не обнаруживается ни в N. sylvestris , ни в N. tomentosiformis .
2B). Картина гибридизации N. tabacum была больше похожа на N. tomentosiformis , чем на N. sylvestris , поскольку оба имели полосы примерно на 3.3 и 4,0 кб. Две слабые полосы гибридизуются с клоном кДНК ASA2 в N. sylvestris . Фрагмент размером 24 т.п.н. в N. tabacum демонстрирует очень слабую гибридизацию и не обнаруживается ни в N. sylvestris , ни в N. tomentosiformis .
Геномный ДНК-блот-анализ. A. Двадцать микрограммов геномной ДНК AB15-12-1 расщепляли девятью рестрикционными ферментами. Дорожки слева направо соответствуют Bam HI, Eco RI, Eco RV, Hin cII, Hin dIII, Kpn I, Pst I, Sca I, и Sca I и И.В. Десять микрограммов каждой геномной ДНК, полученных из листьев N. sylvestris , N. tomentosiformis , N. tabacum и культивированных в суспензии клеток линии клеток табака 5MT s и 5MT r ( AB15-12-1) расщепляли Kpn I. Полноразмерный клон кДНК ASA2 использовали в качестве зонда как в A, так и в B. Размеры фрагментов определяли с помощью лестницы размером 1 т.п.н. (GIBCO-BRL).
Полноразмерный клон кДНК ASA2 использовали в качестве зонда как в A, так и в B. Размеры фрагментов определяли с помощью лестницы размером 1 т.п.н. (GIBCO-BRL).
Сравнение аминокислотных последовательностей генов AS
На основе анализа BLAST (Altschul et al., 1990), пять наилучших совпадений с предсказанной аминокислотной последовательностью гена ASA2 табака выровнены на рисунке 3. Область, кодирующая 1851 нуклеотид ASA2 , будет кодировать полипептид из 616 аминокислот с расчетной молекулярной массой 69 035 D. Ген табака ASA2 кодирует 60 аминокислот за пределами аминоконца выровненных микробных гомологов. Последовательность Ile-58-Glu-59-Ala-60-Ser-61 подобна консенсусному мотиву сайта расщепления ([Val / Ile] X [Ala / Cys] ↓ Ala) для транзитных пептидов хлоропластов растений (von Heijne et al. al., 1989; Молоток и фон Хейне, 1990). Последовательности транзитных пептидов пяти генов AS растений мало гомологичны друг другу. Ген табака ASA2 показывает 72 и 70, 67 и 66 и 32% аминокислотную идентичность с R. graveolens AS α2 и AS α1, A. thaliana ASA1 и ASA2 и Clostridium thermocellum. trpE соответственно (Sato et al., 1989; Niyogi, Fink, 1992; Niyogi et al., 1993; Bohlmann et al., 1995).
graveolens AS α2 и AS α1, A. thaliana ASA1 и ASA2 и Clostridium thermocellum. trpE соответственно (Sato et al., 1989; Niyogi, Fink, 1992; Niyogi et al., 1993; Bohlmann et al., 1995).
Выравнивание аминокислотных последовательностей генов AS растений и прокариот. TASA2, RASA1, RASA2, AASA1, AASA2 и CTRPE соответствуют N. tabacum ASA2 , R. graveolens AS α1 и AS α2, A. thaliana ASA1 и ASA2 и клонов кДНК C. thermocellum trpE соответственно. Прочерки внутри последовательностей указывают на пропуски. Звездочки под последовательностью представляют собой идентичные аминокислоты среди этих шести различных последовательностей AS.Точки под последовательностью указывают на аналогичные аминокислоты. Два консенсусных мотива, Leu-Leu-Glu-Phe-X 10 -Ser и Asn-Pro-Ser-Pro-Tyr-Met, влияющие на ингибирование обратной связи на основе микроорганизмов и дрожжей, выделены жирным шрифтом. Замены аминокислот Phe-Arg и Arg, которые вызвали нечувствительность к обратной связи у N. tabacum и R. graveolens , выделены жирным шрифтом с ромбовидной меткой в положениях 107 и 108 соответственно. Одно изменение аминокислоты в мутанте Arabidopsis (Asp на Asn) выделено жирным шрифтом с крестиком в положении 326.Мотив консенсусной последовательности для транзитных пептидов хлоропластов растений в ASA2 табака подчеркнут в положениях 58-61. Указанные номера аминокислот основаны на аминокислотной последовательности ASA2 табака.
Замены аминокислот Phe-Arg и Arg, которые вызвали нечувствительность к обратной связи у N. tabacum и R. graveolens , выделены жирным шрифтом с ромбовидной меткой в положениях 107 и 108 соответственно. Одно изменение аминокислоты в мутанте Arabidopsis (Asp на Asn) выделено жирным шрифтом с крестиком в положении 326.Мотив консенсусной последовательности для транзитных пептидов хлоропластов растений в ASA2 табака подчеркнут в положениях 58-61. Указанные номера аминокислот основаны на аминокислотной последовательности ASA2 табака.
В бактериях, грибах и дрожжах два консервативных аминокислотных мотива, Leu-Leu-Glu-Ser-X 10 -Ser и Asn-Pro-Ser-Pro-Tyr-Met, связаны с ингибированием Trp-обратной связи (Matsui et al. др., 1987; Калигури, Бауэрле, 1991; Граф и др., 1993). Шесть генов AS на Фигуре 3 не показали аминокислотных изменений в этих консервативных областях, за исключением мотива Leu-Leu-Glu-Ser-X 10 -Ser, где остатки Ser и Gln, обнаруженные в чувствительных к обратной связи AS высших растений, были изменен на Phe-107 и Arg-108 (на основе аминокислотной последовательности ASA2 табака) в табаке ASA2 и на Arg-108 в гене AS α1 R. Гравеоленс . Эти изменения, по-видимому, коррелируют с нечувствительностью к обратной связи, обнаруженной у последних двух ферментов.
Гравеоленс . Эти изменения, по-видимому, коррелируют с нечувствительностью к обратной связи, обнаруженной у последних двух ферментов.
Экспрессия гена AS
Зонд кДНК табака ASA2 обнаружил высокие уровни транскрипта 2,2 т.п.н. на нозерн-блотах только в культивируемых клеточных линиях табака 5MT r , включая 5MT r N. sylvestris , но не в 5MT s -культивированных клеточные линии, листья, корни, стебли и семена после ночного воздействия (рис. 4).
Рис.4.Северный анализ. Двадцать микрограммов общей РНК на дорожку использовали для северной гибридизации. Полноразмерный клон кДНК ASA2 (ASA2) и рРНК (рРНК) использовали в качестве зондов для гибридизации. A. Суммарные РНК получали из однонедельных клеток N. tabacum , культивируемых в суспензии (SC) четырех 5MT s (s) и четырех 5MT r (r) клеточных линий, 5MT s Листья N. tabacum (L), собранные из 3-недельных культур побегов AB15-12-1, и 1-недельные 5MT r N.sylvestris суспензионно культивируемые клетки. B. Различные органы растений были использованы для определения тканевой специфичности. Листья, корни и стебли AB15-12-1, собранные из 3-недельных культур побегов и высушенных зрелых семян и суспензионных культивированных клеток 5MT s (TXD) и клеток 5MT r (AB15-12-1), были используется для приготовления общей РНК.
Тесты комплементации и ингибирования
Нонсенс-мутант E. coli trpE5972 , трансформированный кДНК ASA2 табака и сайт-направленной мутантной формой ASA2 (Phe-107-Arg-108 заменен на Ser-107-Gln-108), оба росли на минимальной среде. содержащие ампициллин и IPTG, но без Trp (рис.5А). Комбинированный штамм, трансформированный сайт-направленным мутантом, однако, не рос на 300 мкм 5MT, содержащем минимальную среду без Trp (фиг. 5B), тогда как рост штамма, трансформированного кДНК ASA2 , не подавлялся на 300 мкм 5МТ.
Рис.5.Комплементация E.coli trpE5972 табаком ASA2 и сайт-направленным мутантом. Мутанты pQE30 / ASA2 и pQE30 / ASA2 были трансформированы в нонсенс-мутант trpE E.coli ( trpE5972 ) и высевали на минимальную среду M9, содержащую 100 мкг / мл ампициллина (A) и 0,1 мм IPTG без и с 300 мкм 5MT (B). Снимок был сделан через 2 дня после появления полос.
Кинетические константы AS
ИнгибированиеTrp частично очищенного продукта генов E. coli -экспрессированного табака ASA2 и Arabidopsis ASA1 показано на Фигуре 6A. Продукт гена ASA2 по-прежнему на 50% активен при 100 мкм Trp, аналогично AS в 5MT-отобранных клетках табака и очищенному R.graveolens AS α1, который был активен на 80% при этой концентрации Trp (Bohlmann et al., 1996).
Рис.6.Кинетика AS из клеток табака дикого типа (□) и 5MT-отобранных (▪) и из E. coli , трансформированных геном ASA1 Arabidopsis () или геном ASA2 табака (♦). A. Относительную активность AS в присутствии Trp измеряли, как описано в Методиках (с хоризматом 100 мкм), и выражали как процент активности без добавления Trp.Удельная активность без Trp для AS из каждой линии табака составляла 73 и 169 нмоль антранилата мин -1 мг белка -1 соответственно. Б. График активности АС Лайнуивера-Берка. Скорость выражается в наномолях в минуту на миллиграмм белка.
При измерении активности AS в экстрактах из суспензионных культур клеток табака дикого типа и 5MT r устойчивые культуры имели часть активности, более нечувствительную к обратной связи, чем у клеток дикого типа (рис.6А). Кажущийся K Значения i для Trp составляют 2 и 300 мкм, соответственно, для AS из клеток табака дикого типа и 5MT-отобранных клеток табака при оценке по концентрации Trp, что приводит к 50% ингибированию, как показано на Фигуре 6A. Об аналогичных значениях сообщили Bohlmann et al. (1996) для R. graveolens Trp-чувствительных и нечувствительных АС (2,8 и> 100 мкм соответственно). AS от Arabidopsis дикого типа имел очевидный вид K i размером 3 мкм, тогда как AS из мутанта, отобранного 6-метил-Trp, имел K i толщиной 8 мкм (Li and Last, 1996).
Явный хоризмат K m Значение было ниже для AS из 5MT-отобранных клеток (6 мкм), чем для клеток дикого типа (22 мкм), при измерении с использованием 10 мм Gln в качестве второго субстрата (фиг. 6B). Эти значения сопоставимы со значениями, полученными для AS из Arabidopsis (10 и 21 мкм с использованием Gln [Li and Last, 1996]) и R. graveolens (17 и 29 мкм с использованием NH 4 + [Bohlmann et al. , 1996]). У всех трех видов хоризмат K м Значение (первое значение в списке) ниже для AS, нечувствительного к обратной связи.
Сравнение последовательностей
ASA2 и последовательностей генов дикого типаФрагмент ASA2 между +181 и +392, который содержит остатки Phe-107-Arg-108, амплифицировали с помощью ПЦР и клонировали из геномных ДНК 5MT s и 5MT r . Когда нуклеотидная последовательность фрагмента из AB15-12-1 была выровнена с нуклеотидной последовательностью из трех Nicotiana sp. клеточные линии (фиг. 7), и по сравнению с кДНК ASA2 было обнаружено, что геномные фрагменты содержат интрон длиной 115 пар оснований; следовательно, клонированные фрагменты имели длину 326 п.н.Нуклеотидные последовательности этого фрагмента были идентичны на 100% во всех случаях, за исключением восьми нуклеотидов в N. sylvestris . Эта разница в восемь нуклеотидов не изменила кодируемые аминокислоты. Аминокислотные последовательности в этой области показали 66% идентичность с четырьмя другими генами растений AS, из Arabidopsis и R. graveolens .
Рис.7.Выравнивание нуклеотидных последовательностей фрагментов геномной ДНК ASA2 . Фрагменты геномной ДНК ASA2 длиной 326 пар оснований, соответствующие последовательности кДНК ASA2 от +181 до +392, были амплифицированы из геномных ДНК листьев табака (ASA2), собранных из культур побегов AB15–12-1 и трех дикорастущих растений. -типа (5MT s ) Nicotiana sp.культивированные клеточные линии, N. tabacum (TXD), N. tomentosiformis (Nto) и N. sylvestris (Ns) с использованием праймеров 4 (смысловой) и 10 (антисмысловой), которые показаны жирным шрифтом. Эти фрагменты геномной ДНК содержат интрон длиной 115 п.н., который подчеркнут. Восемь нуклеотидов, обозначенных (+) над последовательностью, отличаются в геномном фрагменте N. sylvestris по сравнению с нуклеотидами в N. tabacum и N. tomentosiformis . Нуклеотидные последовательности, соответствующие Phe-107-Arg-108, выделены жирным шрифтом и подчеркнуты однобуквенными сокращениями кодонов аминокислот F и R над последовательностью.
Анализ секвенирования промотора
99 нуклеотидов на 3′-конце промоторной области и 5′-конце кДНК ASA2 , включая 5′-UTR, были идентичны (фиг.8). Первый возможный блок TATA (TATAAA) расположен на 121 п.н. выше по течению от сайта начала трансляции (ATG). Анализ BLAST показывает, что область от -1187 до -769 демонстрирует от 81 до 83% идентичности нуклеотидной последовательности с промоторной областью N. tabacum гена Str246C , связанного с защитой растений (Froissard et al., 1994).
Рис.8.Нуклеотидная последовательность области промотора ASA2. Первый возможный блок TATA (TATAAA), стартовый кодон трансляции (ATG), выделен жирным шрифтом. Подчеркнуты 98 п.н. на 3′-конце промоторной области ASA2, которая идентична нуклеотидной последовательности на 5′-конце кДНК ASA2 .
Экспрессия химерного гена GUS
При тестировании различных удаленных фрагментов промоторной области ASA2 временная экспрессия GUS, контролируемая промоторной областью, обозначенной 606, была сильнее как в культивируемых в суспензии клетках, так и в листьях, чем экспрессия, управляемая промотором 35S CaMV (рис.9). Области 1356 и 2252 все еще демонстрировали высокую активность GUS в культивируемых в суспензии клетках, но очень низкую активность в листьях. Область 1356 продемонстрировала примерно в 5 раз более высокую экспрессию GUS, чем экспрессия, управляемая промотором 35S CaMV в клетках, культивируемых в суспензии 5MT r .
Рис.9.Химерные конструкции промотора ASA2 -GUS и временные анализы GUS с листьями табака и культивируемыми в суспензии клетками. Четыре удаленных фрагмента промотора были слиты с репортерным геном GUS с терминатором NOS в pUC19 и обозначены как 2252, 1356, 606 и 370.Эти химерные конструкции и конструкция CaMV 35S-GUS в качестве контроля подвергали бомбардировке двухдневным культивированным клеткам AB15-12-1 (черные столбцы) и листьям (белые столбцы), собранным из культур побегов 3-недельного возраста. Временную экспрессию GUS измеряли с помощью анализа MUG через 3 дня после бомбардировки. Представлены средние значения после вычитания фона из четырех отдельных экспериментов с двумя повторностями. GUS-специфическая активность определялась как пикомоль 4-метилумбеллиферона в час на миллиграмм белка.
Чтобы определить, вызвана ли специфическая для культуры ткани экспрессия, контролируемая промотором ASA2, фактором (ами) транскрипции в клеточных линиях 5MT r , мы провели отдельные эксперименты с использованием культивированных в суспензии клеток из 5MT s N. tabacum и из 5MT s и 5MT r N. sylvestris . Временные активности GUS в суспензионных клетках N. tabacum 5MT r (AB15-12-1), контролируемых промоторами 35S, 606, 1356 и 2252, были приблизительно равны 1.В 8, 7,4, 3,7 и 3,6 раза больше, чем у клеток, культивируемых в суспензии табака 5MT s , соответственно. В случае N. sylvestris в суспензионно-культивируемых клетках 5MT r были обнаружены 3,4-, 10,6-, 6,4- и 9,1-кратные активности GUS по сравнению с суспензионно-культивированными клетками 5MT s . соответственно.
То, что промотор ASA2 демонстрирует тканеспецифическую экспрессию, также было показано в экспериментах по стабильной трансформации табака с использованием Agrobacterium tumefaciens (данные не показаны).
ОБСУЖДЕНИЕ
Мы клонировали кДНК ASA2 табака, кодирующую α-субъединицу нечувствительной к обратной связи AS и промоторную область. Саузерн-гибридизационный анализ показывает, что в геноме табака присутствует более одной близкородственной последовательности (рис. 2). Как и все гены AS растений, описанные на сегодняшний день, ASA2 табака имеет предполагаемую последовательность транзитного пептида; следовательно, фермент, по-видимому, локализован в пластидах.
Наши результаты подтверждают вывод о том, что ASA2 является геном дикого типа, кодирующим α-субъединицу нечувствительной к обратной связи AS табака.Эти результаты включают нечувствительную к обратной связи экспрессию гена ASA2 в мутанте trpE штамм E. coli (фиг.5), ферментно-кинетический анализ (фиг.6), нуклеотидную последовательность дикого типа ASA2 геномной фрагменты (фиг. 7) и высокий уровень экспрессии в суспензионных культивируемых клетках 5MT r (фиг. 4).
Было показано, что несколько участков аминокислотной последовательности AS влияют на ингибирование с помощью обратной связи (Bohlmann et al., 1996; Kreps et al., 1996; Li and Last, 1996).В табаке ASA2 как из линий клеток дикого типа, так и из линий клеток 5MT r две аминокислоты Phe-107 и Arg-108, обнаруженные в той же области, что и в AS α1 (Bohlmann et al., 1996), отличались от таковых, обнаруженных в чувствительных к обратной связи AS, Ser-107 и Gln-108. Когда мы заменили остатков ASA2 Phe-107 и Arg-108 на Ser-107 и Gln-108 с помощью сайт-направленного мутагенеза, продукт гена все еще был активен в E. coli , но он дополнял E. coli штамм не мог расти на 300 мкм 5MT, содержащем минимальную среду без Trp, тогда как исходный штамм с дополнением ASA2 мог.Кроме того, продукт гена ASA2 , экспрессируемый E. coli , был нечувствительным к обратной связи, как и AS, обнаруженный в экстрактах клеток табака 5MT r . Эти результаты показывают, что остатки Phe-107 и Arg-108 важны для контроля ингибирования обратной связи. Дальнейшие мутагенез и кинетические исследования и исследования связывания будут необходимы для определения эффекта других аминокислотных изменений в этой области.
Кажущийся Trp K i значений для чувствительных к обратной связи AS из табака, R.graveolens и Arabidopsis схожи, что и можно было ожидать, учитывая высокую степень идентичности в областях, которые, как известно, влияют на ингибирование бактериальной AS с помощью обратной связи, которые также считаются важными для AS растений (Caliguri and Bauerle, 1991; Bohlmann et al ., 1995). Кажущийся Trp K Значения i для табака и R. graveolens нечувствительных к обратной связи ферментов AS аналогичны друг другу и намного выше, чем K . i значение мутантного нечувствительного к обратной связи AS Arabidopsis.Этот образец констант ингибирования коррелирует с наблюдаемыми гомологиями последовательностей и подтверждает гипотезу о том, что замена Arg-163 в R. graveolens (соответствующая Arg-108 табака) и Phe-107-Arg-108 в AS табака связаны к более высокой нечувствительности к обратной связи, чем у мутантного AS Arabidopsis, когда Asp-341 заменяется на Asn.
То кажущееся K м. Значения для хоризмата для AS табака аналогичны значениям, полученным для Arabidopsis и R.graveolens AS также неудивительно, учитывая высокую степень сходства, особенно в области, которая, как известно, участвует в катализе бактериального AS (Caliguri and Bauerle, 1991; Bohlmann et al., 1995). У всех трех видов кажущаяся K м для хоризмата нечувствительного к обратной связи AS ниже, чем K м для чувствительных к обратной связи AS. Это предполагает аналогичный механизм торможения и нечувствительность к обратной связи. Этого следовало ожидать для табака и р.graveolens , поскольку, по-видимому, задействована аналогичная разница в последовательностях, но мутант Arabidopsis содержит другое структурное изменение, которое продуцирует фермент, который все еще заметно ингибируется Trp.
Представленные здесь результаты также показывают, что промотор ASA2 табака регулирует тканеспецифическую экспрессию гена. Этот вывод подтверждается измерениями экспрессии генов на уровне мРНК (рис.4) и исследованиями делеционного анализа промоторной области ASA2 (рис.9). Высокий уровень экспрессии гена ASA2 был обнаружен на уровне мРНК только в суспензионных культивируемых клетках 5MT r , что аналогично предыдущим результатам, полученным на уровне активности фермента (Brotherton et al., 1986). Кроме того, временная экспрессия GUS была выше в клетках 5MT r , чем в культивируемых в суспензии клетках 5MT s , и очень низкий уровень экспрессии был обнаружен в листьях. Эта тканеспецифическая экспрессия контролировалась областью промотора между -2252 и -607.Эти результаты предполагают, что могут быть изменения в количестве или типе регуляторных белков в суспензионных культивируемых клетках 5MT r , которые активируют экспрессию гена ASA2 . Изменения в цис--действующих элементах в промоторной области, вероятно, можно исключить, поскольку мы обнаружили только несколько нуклеотидных различий в промоторной области ASA2, выделенной из трех клеточных линий табака дикого типа, по сравнению с ASA2. Мы также обнаружили сходные паттерны временной экспрессии с этими промоторами по сравнению с паттернами, обнаруженными с промотором ASA2 из AB15-12-1 (данные не показаны).
У Arabidopsis и R. graveolens экспрессия гена ASA1 и AS α1 индуцируется, тогда как гены ASA2 и AS α2 экспрессируются конститутивно. Несмотря на то, что мы обозначили наш клон ASA2 , поскольку наивысшая аминокислотная идентичность была с R. graveolens AS α2 (72%), и поскольку мы выделили укороченную кДНК ASA1 , аналогичную кДНК Arabidopsis ASA1 (данные не показан), табак ASA2 также очень похож на R.graveolens AS α1 (идентичность 70%). До сих пор нам не удалось индуцировать синтез мРНК ASA2 листа ASA2 путем обработки ауксином или салициловой кислотой (в течение 8 часов) или путем ранения (в течение 48 часов). Чтобы помочь понять экспрессию гена ASA2 in vivo в растениях и влияние условий окружающей среды на экспрессию генов, мы недавно получили растения табака, трансформированные репортерным геном GUS, управляемым промотором ASA2 длиной 2252 п.н. Нечувствительный к обратной связи R. graveolens AS α1, по-видимому, участвует в пути синтеза алкалоидов, производных антранилата (Bohlmann et al., 1996). Однако нам не известны какие-либо идентифицированные антранилатные или индольные фитоалексины или алкалоиды табака; следовательно, функция гена ASA2 в табаке еще не определена.
В этой статье мы сообщили о клонировании гена ASA2 табака дикого типа, кодирующего α-субъединицу AS, нечувствительного к обратной связи. Нечувствительность к обратной связи в суспензионных культивируемых клетках 5MT r коррелировала с высоким уровнем экспрессии гена ASA2 и тканеспецифической экспрессией этого гена, контролируемой определенными промоторными областями.Роль нечувствительных к обратной связи AS в табаке до сих пор неизвестна, и необходимы дальнейшие исследования, чтобы понять регуляцию и функцию этого фермента у растений.
БЛАГОДАРНОСТИ
Мы благодарим Кришну Нийоги за клоны кДНК, Mani Subramanian за линию E. coli, экспрессирующую Arabidopsis ASA1, Чарльза Янофски и Центр генетических ресурсов Escherichia coli за мутантные штаммы E. coli , Роба Аллемана за техническую помощь с кинетическим анализом AS, и Люген Чжу за техническую помощь с анализами MUG.
Сноски
↵1 Это исследование было поддержано фондами Совета Иллинойса по продовольственным и сельскохозяйственным исследованиям и Иллинойсской сельскохозяйственной экспериментальной станции.
↵ * Автор, ответственный за переписку; электронная почта widholm {at} uiuc.edu; факс 1–217–333–4777.
Сокращения:
- AS
- антранилатсинтаза
- CaMV
- Вирус мозаики цветной капусты
- IPTG
- изопропилтиогалактозид
- MS
- Мурашиге -143-143-14914 914 914 914 914 914 914 914 914 519Murashige-14-Skoog 914 Methyl 914 914 914 Tr -Trp-устойчивый
- 5MT s
- 5-метил-Trp-чувствительный
- MUG
- 4-метилумбеллиферона глюкуронид
- RACE
- быстрая амплификация концов кДНК
14 UTRA
ЦИТИРОВАННАЯ ЛИТЕРАТУРА
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
Haslam E (1993) Шикимовая кислота: метаболизм и метаболиты. John Wiley & Sons, Чичестер, Великобритания, стр. 156–254
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
Ochman H, Ajioka JM, Garza D, Hartl L (1989) Обратная полимеразная цепная реакция. В HA Erlich, ed, PCR Technology. Stockton Press, New York, pp 105–111
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
Yanofsky C, Crawford IP (1987) Оперон триптофана. В FC Neidhardt, JL Ingraham, KB Low, B Magasanik, M Schaechter, HE Umbarger, eds, Escherichia coli и Salmonella typhimurium : Клеточная и молекулярная биология.Американское общество микробиологии, Вашингтон, округ Колумбия, стр. 1453–1472
- ↵
LG4525B
% PDF-1.4 % 1 0 объект > эндобдж 2 0 obj > эндобдж 3 0 obj > поток Acrobat Distiller 6.0 (Windows) LG4525BPScript5.dll Версия 5.2.22009-03-04T20: 22: 30 + 09: 002018-04-19T12: 25: 54 + 04: 00uuid: 1984c6b2-a295-42cc-9b72-f1c96760e7beuuid: 8d601f5a- a4ca-43b6-84ab-83b813ef7ca0application / pdf
ChIP-анализ раскрывает исключительно широкое распределение участков связывания ДНК для фактора транскрипции NtcA в цианобактериях, образующих гетероцисты.
Иммунопреципитация ДНК, связанной с NtcA
Дикий тип Anabaena sp.Клетки PCC 7120, растущие в барботажных культурах с аммонием в качестве источника азота, подвергали инкубации в объединенной обедненной азотом среде в течение 3 часов, после чего культуры обрабатывали формальдегидом для фиксации белков, связанных с ДНК. После лизиса клеток и фрагментации ДНК экстракты обрабатывали антителом против NtcA для специфического иммунопреципитации ДНК, связанной с NtcA (см. Методы). Затем иммунопреципитированный материал инкубировали при 65 ° C для обратного сшивания и выделяли ДНК.Образец тотальной ДНК также выделяли перед обработкой экстрактов анти-NtcA, чтобы служить в качестве входного контрольного образца.
Количественная ПЦР была проведена для проверки качества иммунопреципитированной ДНК и подтверждения того, что известные целевые области NtcA были обогащены. Использовали праймеры, которые амплифицировали промоторную область nrrA (all4312) в качестве положительного контроля и промоторную область ORF all0770 в качестве отрицательного контроля. Результат анализа Q-PCR (дополнительный файл 1: рисунок S1) подтвердил значительное обогащение NtcA-зависимого промотора.Иммунопреципитированные и входящие образцы ДНК были подвергнуты высокопроизводительному секвенированию, и результаты были проанализированы с использованием алгоритма Triform [16] и картированы на геном Anabaena sp. PCC 7120 [17] (Дополнительный файл 2: Таблица S1).
Распределение ДНК, связанной с NtcA, по геному
Anabaena sp. PCC 7120Анализ иммунопреципитированной ДНК показал 2424 связывающих участка, все из которых статистически значимы, расположены в геноме Anabaena и распределены по хромосоме и пяти из шести плазмид (Таблица 1; Дополнительный файл 3: Рисунок S2) .Мы проанализировали расположение этих связывающих областей на геномной последовательности Anabaena и отнесли их к одному гену, двум генам (когда было невозможно выбрать между фланкирующими генами) или мРНК (дополнительный файл 2: таблица S1). Программа Integrative Genome Viewer (Broad Institute) [18] использовалась для картирования последовательностей связывающих областей, полученных в эксперименте ChIP-Seq, на геном Anabaena (данные доступны для его визуализации по адресу GEO, доступ GSE51865).Информация о 2424 полученных областях связывания показана в Дополнительном файле 2: Таблица S1, включая расположение в хромосоме или плазмидах, ген (ы), которым приписана область связывания, и статистическая значимость пика, идентифицирующего область связывания (значение Q, значение NLQ) (см. Методы). Области связывания имеют длину около 200–250 п.н., что является их средней точкой, наиболее вероятной позицией для связывания NtcA. Поскольку 5′-UTR цианобактерий часто бывают очень длинными [13, 14], некоторые связывающие области были отнесены к относительно далеким генам, даже если связывающие области были расположены внутри кодирующей области соседнего гена.Для этих случаев мы приняли во внимание информацию о дифференциальной экспрессии генов после депривации комбинированного N [13, 14, 19] (дополнительный файл 2: Таблица S1).
Таблица 1 Результаты ChIP-Seq NtcA в Anabaena sp. РСС 7120 после N понижающейЧтобы оценить роль связывания NtcA в связывающих областях, программы CLC Sequence Viewer (CLC Bio) и Artemis Genome Browser (Wellcome Trust Sanger Institute) [20] были использованы для интеграции транскриптомных данных из Flaherty et al. . [13] и из Mischke et al. [14] с расположением последовательностей связывающей области NtcA, полученных в нашем эксперименте ChIP-Seq. С этой целью местоположение определенной области связывания (средняя точка диапазона обнаруженной целевой области) (дополнительный файл 2: таблица S1) было определено с использованием вышеупомянутых программ, и ее геномный контекст был проанализирован по отношению к транскриптам. / сайты начала транскрипции, которые были описаны. Кроме того, предполагаемые консенсусные сайты связывания NtcA были идентифицированы (если возможно) в последовательности области связывания (таблица 2; дополнительный файл 2: таблица S1).Также было проанализировано относительное расположение области связывания по отношению к гену, и области связывания были разделены на три категории: выше по течению от гена (различая между дальним и близким по течению), внутренним по отношению к гену (близко к 5 ‘ или 3’-конец гена) и ниже гена (таблица 1 и дополнительный файл 2: таблица S1). Интересно, что около 65% идентифицированных областей связывания были обнаружены в кодирующих областях. Среди тех, что расположены в межгенных областях, большинство из них (865) присутствовали перед назначенными генами и некоторые (79) ниже кодирующих областей.
Таблица 2 Целевые регионы с наибольшим количеством балловФункциональные категории генов с сайтами связывания NtcA
Гены, идентифицированные как мишени NtcA через 3 часа после снижения N, были разделены на восемь функциональных категорий (таблица 3). Около половины назначенных генов (1074) кодируют белки с неизвестной функцией. Среди генов, кодирующих белки, участвующие в клеточных процессах (234 гена), 90 были генами, связанными с трансляцией, включая несколько рибосомных белков, а 22 были связаны с транскрипцией.Среди генов, кодирующих белки, участвующие в метаболизме и фиксации азота, 33 гена участвовали в поглощении и метаболизме азота, включая хорошо известные NtcA-зависимые гены, такие как ntcB , nirA , amt4 , cphB и т. Д. . [4, 8, 21].
Таблица 3 Функциональные категории присвоенных геновВсего было обнаружено 179 генов, кодирующих регуляторные белки, некоторые из которых уже были известными NtcA-мишенями, включая гены, кодирующие железо-чувствительный репрессор транскрипции FurA [22], регуляторный фактор PipX [23], двухкомпонентный ответ регулятор NrrA [24] и гены pkn41 и pkn42 , кодирующие Ser / Thr киназы с двухкомпонентными сенсорными доменами [25].Некоторые из недавно идентифицированных регуляторных генов со связывающими областями NtcA были генами, кодирующими репрессор транскрипции SmtB (присутствующий в бета-плазмиде), двухкомпонентный регулятор ответа RpaA или фосфатазу Ser / Thr Alr3732. Интересно, что доля внутренних областей связывания NtcA в генах, кодирующих регуляторные белки, была выше, чем в других категориях генов. Что касается генов, связанных с метаболизмом, то было 55 генов центрального метаболизма, включая гены ассимиляции углерода и активированные гены (например,грамм. zwf ) или репрессированные (например, rbcL ) в гетероцистах [10].
Анализ областей связывания с наивысшим значением NLQ
Все области связывания, идентифицированные с помощью анализа ChIP-Seq, имели чрезвычайно низкие значения Q из-за исключительного качества глубокого секвенирования иммунопреципитированной ДНК. Мы использовали значение NLQ для анализа областей связывания, что позволило нам отсортировать те области со значением Q, близким к нулю. Список из 40 наиболее представленных областей связывания (соответствующих наивысшим значениям NLQ) показан в таблице 2.Внутри этих целевых областей мы локализовали предполагаемые NtcA-связывающие последовательности, которые в большинстве случаев были обнаружены около середины целевой области и, когда это возможно, они коррелировали с ранее описанными TSP (все из которых дифференциально экспрессировались в условиях N-истощения. ) по сравнению с данными Flaherty et al. [13] и Mitschke et al. [14], а также из отдельных анализов [21, 25–32].
Некоторые области-мишени были включены в последовательности ДНК, соответствующие NtcA-зависимым промоторам класса II.Среди них целевая область с наивысшим значением NLQ (# 1203), которая была расположена выше ORF alr2921 , с последовательностью TAC GTGN 8 , отделенной 35 нуклеотидов от основной массы концов 5′-транскрипта, активированных в сочетании Среда, свободная от N [13], которая может представлять предполагаемый TSP, которому предшествует блок -10 (рисунок 1). В других случаях целевая область была включена в последовательности ДНК, показывающие положение, подобное репрессору. Наконец, в других областях-мишенях может быть обнаружен только предполагаемый сайт связывания без корреляции с каким-либо известным TSP.В некоторых случаях один сайт связывания может влиять на экспрессию двух TSP (оба одного и того же гена или двух разных), а в других предполагаемый сайт связывания может быть приписан двум генам (без очевидной связи с каким-либо TSP). Целевая область со вторым по величине значением NLQ (область № 1856) была расположена выше гена ilvG , кодирующего фермент биосинтеза аминокислот с разветвленной цепью, который слегка репрессируется в комбинированной среде, не содержащей N [13, 19]. . Сайт связывания NtcA, идентифицированный в этой целевой области, GAAN 8 TAC, перекрывает TSP, который репрессируется в условиях обеднения N (таблица 2 и [14]).В дополнение к репрессорному эффекту, описанному выше, последовательность связывания NtcA перед ilvG отделена 22 нуклеотидами от -10 бокса N-зависимого активированного TSP, что представляет собой предполагаемый сайт активатора промотора класса II. Это похоже на то, что мы обнаружили для целевой области, расположенной выше glnA , которая уже была описана (Таблица 2 и [27]).
Рисунок 1Нуклеотидный анализ области-мишени с наивысшей оценкой в эксперименте ChIP-Seq, показывающий присутствие сайта связывания NtcA.A) Местоположение целевой области (черным цветом) относительно соседних ORF и в контексте транскриптомных данных Flaherty et al. [13]. B) Увеличенное изображение средней точки целевой области (обведено A), предполагаемого TSP (G) и предполагаемой NtcA-связывающей последовательности (GTGTGTCTTGATAC), расположенных 22 нуклеотида из предполагаемого -10-бокса (TATGCT) (последовательность CLC Зритель). Горизонтальные серые линии обозначают транскрипцию [13].
На рис. 2 показаны два примера областей-мишеней, последовательности которых содержат NtcA-зависимую структуру промотора класса II.Одним из них является хорошо известный NtcA-зависимый ген ntcB , промоторные детерминанты которого уже были описаны [30] (Рисунок 2A и Таблица 2). Другой пример – целевая область с высоким NLQ, картированная внутри ORF alr2482 (целевая область № 1165), последовательность которой также демонстрирует структуру промотора, активируемого NtcA класса II, которая может регулировать внутренний TSP, активированный при депривации N [14 ].
Рисунок 2Нуклеотидный анализ двух областей-мишеней с высокой оценкой.A) Промоторная последовательность ntcB ( all0602 ), включая сайт связывания NtcA, блок -10 и TSP (или TSS) (уже описано) [30], а также среднюю точку целевой области, обнаруженной в ChIP-Seq эксперимент (обведен G). B) Анализ последовательности целевой области, обнаруженной внутри ORF alr2482 (средняя точка целевой области, представленной кружком C), которая включает предполагаемый сайт связывания NtcA, отделенный 22 нуклеотида от предполагаемого блока -10 внутреннего регулируемый TSP [14].Транскриптомные данные были получены из [13] (12 ч после отмены азота).
Связывание NtcA in vitro
Чтобы подтвердить наши результаты, мы сначала проверили, идентифицировал ли наш анализ ChIP-Seq уже известные сайты связывания NtcA, расположенные в промоторных областях генов, регулируемых этим фактором транскрипции. Действительно, Дополнительный файл 4: Таблица S2 показывает восемнадцать областей-мишеней, идентифицированных в этой работе, которые включают сайты связывания NtcA, идентифицированные ранее различными способами, такими как те, которые соответствуют генам ассимиляции нитрата nirA и ntcB , гену ассимиляции аммония glnA и связанные с гетероцистами гены hetC , xisA и devB , среди прочих.
Кроме того, были проведены анализы сдвига электрофоретической подвижности (EMSA) для проверки способности NtcA связываться in vitro с фрагментами ДНК, включенными в различные области связывания (рис. 3), идентифицированные в этом исследовании. Целевые области № 605 и № 931 (которые включают сайты связывания NtcA, расположенные выше rbcL и glnA , соответственно [дополнительный файл 2: таблица S1]) использовали в качестве положительных контролей. В качестве отрицательного контроля мы использовали фрагмент ДНК перед all2096 , внутреннюю область nrrA и промоторную область гена Amaranthus hybridus psbA , которые не были описаны как мишени NtcA.Был протестирован выбор из двенадцати областей связывания с различными характеристиками. Мы включили связывающие области с низким NLQ (№ 204, 259, 602, 1128 и 1570) и с высоким NLQ (№ 364, 996, 1135, 1137, 1203 и 1756). Анализируемые связывающие области были расположены внутригенно (№204, 259, 364, 602, 1135 и 1570), выше (№892, 996, 1128, 1203 и 1756) или ниже (№1137) генов. Предполагаемый консенсусный сайт связывания NtcA был идентифицирован в областях связывания # 204, 364, 602, 892, 996, 1128, 1137, 1203 и 1570, в то время как области связывания # 602, 996, 1128 и 1203 были обнаружены рядом с TSP (класс II -зависимая активация совместимых сайтов) (Рисунок 3).В анализах EMSA использовалось несколько концентраций NtcA, и в целом наблюдалась корреляция между NLQ и сродством NtcA к области связывания, что указывало на качество результатов. NtcA показал более высокое сродство к областям связывания с высоким NLQ, чем к участкам с низким NLQ, независимо от положения областей связывания по отношению к генам.
Рисунок 3Анализ сдвига электрофоретической подвижности нескольких областей связывания. Результаты EMSA, проведенного с использованием фрагментов ДНК из указанных испытанных областей связывания с использованием указанных концентраций NtcA.Указываются значение NLQ, положение относительно гена, приписанный ген, идентифицированный сайт связывания NtcA и присутствие TSP.
Консенсусный сайт связывания NtcA
Первоначальный анализ CisFinder всех данных, полученных в результате глубокого секвенирования иммунопреципитированной ДНК, с использованием случайно расположенных подпоследовательностей в качестве контроля, показал преобладающий мотив, специфичный для целевой области: GTAN 8 TAC. Мы провели дополнительный анализ последовательностей связывания NtcA, идентифицированных с помощью приложения Weblogo [33] (рис. 4).Была проанализирована консенсусная последовательность трех различных групп сайтов связывания NtcA. Первый включал предполагаемые последовательности NtcA, идентифицированные в 40 областях-мишенях с наивысшим NLQ (рис. 4A). Была получена хорошо известная последовательность сайта связывания NtcA [4], имеющая несколько консервативных нуклеотидов Т и С в положениях 4 и 5 и А в положениях 10 и 11. Когда были идентифицированы 508 предполагаемых последовательностей NtcA (в основном расположенных в областях-мишенях с самыми высокими значениями NLQ; Дополнительный файл 2: Таблица S1), был получен консенсусный сайт связывания NtcA, показанный на Фигуре 4B.В этом случае GTN 10 AC был высококонсервативным, с некоторым преобладанием A в положении 3 и T в положении 12. Наконец, мы провели анализ расширенного сайта связывания NtcA (т. Е. Включая 6 нуклеотидов выше и ниже). 6 нуклеотидов ниже ядра GTAN 8 TAC). В этом случае использовалось всего 135 последовательностей, полученных либо из областей-мишеней с наивысшим NLQ, либо из тех сайтов связывания NtcA, которые были связаны с промотором (указаны в дополнительном файле 2: Таблица S1).Наблюдалась четкая консервация вдоль последовательности длиной 24 нт, особенно наличие пар A / T в положениях 3, 4, 6, 21, 23 и 24 (рис. 4C), что согласуется с предыдущими наблюдениями [4, 11].
Рисунок 4Консенсусный сайт связывания NtcA. A) Консенсусная последовательность NtcA, полученная из 40 областей-мишеней с наивысшим NLQ (таблица 2). B) Консенсусный сайт связывания, обнаруженный с использованием всех предполагаемых связывающих последовательностей NtcA в Дополнительном файле 2: Таблица S1 (508 последовательностей). {\ text {r}} $), клеточная линия табака (Nicotiana tabacum) (AB15-12-1).Строки {\ text {r}} $, скорее всего, связаны с изменениями в регуляторном гене, влияющем на экспрессию ASA2.
Journal InformationМеждународный журнал «Физиология растений», основанный в 1926 году, посвящен физиологии, биохимии, клеточной и молекулярной биологии, генетике, биофизике и экологической биологии растений. Физиология растений – один из старейших и наиболее уважаемых журналов по науке о растениях.
Информация об издателеOxford University Press – это отделение Оксфордского университета.Издание во всем мире способствует достижению цели университета в области исследований, стипендий и образования. OUP – крупнейшая в мире университетская пресса с самым широким присутствием в мире. В настоящее время он издает более 6000 новых публикаций в год, имеет офисы примерно в пятидесяти странах и насчитывает более 5500 сотрудников по всему миру. Он стал известен миллионам людей благодаря разнообразной издательской программе, которая включает научные работы по всем академическим дисциплинам, библии, музыку, школьные и университетские учебники, книги по бизнесу, словари и справочники, а также академические журналы.
