
Характеристики и параметры интегральных микросхем
1. Характеристики и параметры интегральных микросхем
2. Общие параметры ИМС
• Напряжение(я) питания Uпит (В)• Диапазон рабочих температур
• Температура конденсации влаги при
относительной влажности 98%
• Вибрационные нагрузки по частоте (Гц)
• Вибрационные нагрузки по ускорению
(м/с2)
• Гарантированная наработка на отказ (ч)
• Интенсивность отказов – плотность вероятности
возникновения отказа в заданном промежутке
времени:
N N1(t ) 1
1 / ч
N
T
N1 – число ИМС безотказно проработавших к
моменту времени t; N – число изначально исправных
ИМС; T – время проведения испытаний.
4. Параметры аналоговых ИМС
• Параметры аналоговых ИМСсоответствуют параметрам устройств,
которые реализованы в данных
микросхемах.
# для интегральных усилителей
основными параметрами будут
коэффициент усиления, диапазон
усиливаемых частот.
 В качестве
В качествехарактеристик АЧХ, ФЧХ и др.
5. Параметры цифровых интегральных микросхем
• Уровни напряжений логического «0» илогической «1»:
# для ТТЛ
U0 ≤0.4В; U1 ≥2.4 В.
# для КМОП логики
U0 ≤ 0,3Uпит; U1 ≥0.7 Uпит.
• Коэффициент разветвления по выходу
Кразв– число входов базовых элементов
данной серии, которое можно
подключить к выходу элемента без
дополнительных устройств
согласования.
# для ТТЛ Кразв = 10
для КМОП Кразв = 100
• Коэффициент объединения по входу Коб
– число входов данного элемента.
• Статическая помехоустойчивость –
определяется уровнем напряжения,
подаваемого на вход элемента
относительно 0 и 1 при котором
состояние микросхемы не меняется.
Uпор – пороговый уровень переключения микросхемы. При его
достижении микросхема переходит из одного логического состояния в
другое;
U 0ст.пу – уровень статической помехоустойчивости относительно «0»;
U 1ст.
 пу – уровень статической помехоустойчивости относительно «1».
пу – уровень статической помехоустойчивости относительно «1».U 0ст.пу = Uпор – U0;
U 1ст.пу = U 1 – Uпор.
# Для ТТЛ |U 0ст.пу| = |U 0ст.пу| ≥ 0,4
• Динамическая помехоустойчивость –
оценивается при подаче на вход
элемента импульсов определенной
формы, длительности и амплитуды.
• Параметры динамической
помехоустойчивости: амплитуда помехи
Uп и ее длительность τп
• Быстродействие – определяется по
среднему времени задержки
распространения сигнала:
• tзд.р.ср.=0,5*(tзд.р.1,0+ tзд.р.0,1)
* быстродействие сложной логической
схемы находится как сумма tзд.р.ср. для
всех последовательно включенных
микросхем.
# для ТТЛ tзд.р.ср. ~ 20 нс,
для КМОП tзд.р.ср. ~ 200 нс,
* также для оценки быстродействия
используют предельную рабочую
частоту fпр. Для ТТЛ fпр =10 МГц, для
КМОП fпр = 1 МГц.
• Средняя статическая мощность
потребления Pср.
• Pср= 0.5*(P0 + P1), где
выходе; P1 – мощность при логической
«1» на выходе.

* Также для оценки потребляемой энергии
используют параметр ток потребления
Iпотр (мА).
• Динамическая мощность Pдин –
мощность, измеренная на предельной
частоте. Pдин > Pср.
В общем случае Pдин= f*Cн*(U1-U0), где
f – частота входных импульсов,
Cн – емкость нагрузки.
• Предельные входные токи при сигналах
логического «0» и «1»:
I0вх; I1вх
| — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | ÷ | -40 ÷ 85 | — | |||
| — | 1,1÷3,3 | — | ±3 | — | — | — | — | — | — | — | — | 0,4 | — | — | — | — | — | — | — | — | 1. 5 5 | 1,7÷3,65 | 100÷400 | 3 ÷ 5.5 | -60 ÷ 85 | — | ||||
| — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | 23,5 | 6000 | — | — | — | — | 3.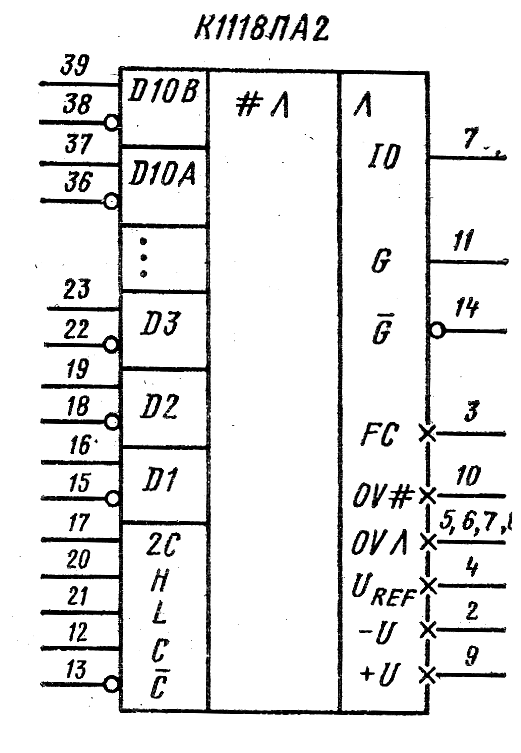 0 ÷ 3.6 0 ÷ 3.6 | -60 ÷ 85 | — | ||||
| — | — | — | — | — | — | — | — | — | — | — | — | 1 | 1 | 55 | — | — | — | — | — | — | — | — | — | — | ÷ | -45 ÷ 85 | — | |||
| — | — | — | — | — | — | — | — | — | — | — | — | 1 | 1 | 55 | — | 16М (2М*8) | — | — | — | — | — | — | — | — | 3 ÷ 3. 6 6 | -60 ÷ 125 | — | |||
| — | — | — | — | — | — | — | — | — | — | — | — | 2 | 2 | 20 | — | 1М (128К*8) | — | — | — | — | — | — | — | — | 3. 0 ÷ 5.5 0 ÷ 5.5 | -40 ÷ 85 | — | |||
| — | — | — | — | — | — | — | — | — | — | — | — | 2 | 2 | 65 | — | 1М (128К*8) | — | — | — | — | — | — | — | — | 3 ÷ 5.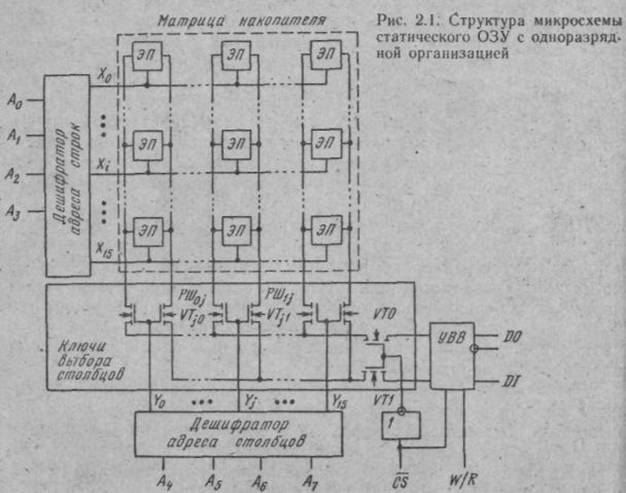 5 5 | -60 ÷ 125 | — | |||
| — | — | — | — | — | — | — | — | — | — | — | — | — | 30 | 10 | 16384 | 16М (1М*16) | — | — | — | — | — | — | — | — | 3 ÷ 3. 6 6 | -60 ÷ 100 | — | |||
| — | — | — | — | — | — | — | — | — | — | — | — | — | 30 | 10 | 16384 | 16М (1М*16) | — | — | — | — | — | — | — | — | 3 ÷ 3. 6 6 | -60 ÷ 100 | — | |||
| — | — | — | — | — | — | — | — | — | — | — | 1 | — | — | — | — | — | — | — | — | — | — | — | — | — | 3 ÷ 3. 6 6 | -40 ÷ 85 | — | |||
| — | — | — | — | — | — | — | — | — | — | 200 | — | — | — | — | — | — | — | — | — | — | — | — | — | — | 3,0 ÷ 3,6 | -60 ÷ 105 | — | |||
| — | — | — | — | — | — | — | — | — | — | 200 | — | — | — | — | — | — | — | — | — | — | — | — | — | — | 3,0 ÷ 3,6 | -40 ÷ 85 | — | |||
| — | — | — | — | — | — | — | — | — | 2 | 144 | — | — | — | — | — | — | 10/100 | 10/100 | — | — | — | — | — | — | 3 ÷ 3. 6 6 | -40 ÷ 85 | — | |||
| — | — | — | — | — | — | — | — | — | 2 | 80 | 1 | — | — | — | — | — | — | — | — | — | — | — | — | — | 2. 2 ÷ 3.6 2 ÷ 3.6 | -40 ÷ 85 | — | |||
| — | — | — | — | — | — | — | — | — | 3 | 160 | 1 | — | — | — | — | — | 1 | — | — | — | — | — | — | — | 3. 0 ÷ 3.6 0 ÷ 3.6 | -40 ÷ 85 | — | |||
| — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | ÷ | -50 ÷ 85 | — | |||
| — | — | — | — | — | — | — | — | — | — | 36 | — | — | — | — | — | — | — | — | — | — | — | — | — | — | 2. 2 ÷ 3.6 2 ÷ 3.6 | +85 ÷ -40 | — | |||
| — | — | — | — | — | — | — | — | — | — | 36 | — | — | — | — | — | — | — | — | — | — | — | — | — | — | 3,0 ÷ 3,6 | +85 ÷ -40 | — | |||
| 1 | — | 16 | — | 10М – 95М | 75,0 | 96 | ±3,0 | — | — | 10 – 480 | — | 303 | 0.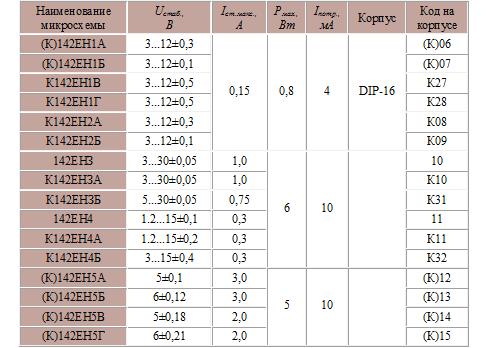 16 16 | — | — | — | — | — | — | — | — | — | — | — | 1.7 ÷ 1.9 | -40 ÷ 85 | — | |||
| — | — | — | — | — | — | — | — | — | — | — | — | 1. 8 8 | — | — | — | — | — | — | — | — | — | — | — | — | 4.5 ÷ 5.5 | -45 ÷ 85 | — | |||
| — | — | — | — | — | — | — | — | — | — | — | — | 1 | — | — | — | — | — | — | — | — | — | — | — | — | 4. 5 ÷ 5.5 5 ÷ 5.5 | -45 ÷ 85 | до 1 Мбит/с | |||
| — | — | — | — | — | — | — | — | — | — | — | — | 7 | — | — | — | — | — | — | — | — | — | — | — | — | 3 ÷ 5. 5 5 | -60 ÷ 125 | не более 30 Мбит/c | |||
| — | — | — | — | — | — | — | — | — | — | — | — | 2 | — | — | — | — | — | — | — | — | — | — | — | — | 3 ÷ 5.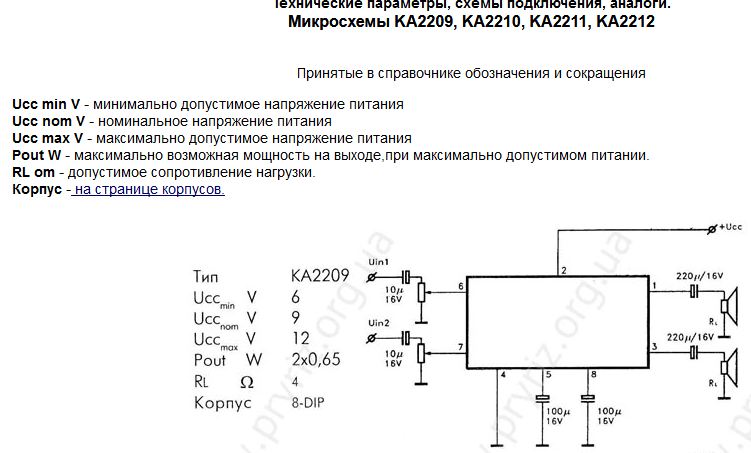 5 5 | -60 ÷ 85 | до 250Кбит/с; до 1Мбит/с | |||
| — | — | — | — | — | — | — | — | — | — | — | — | 0. 03 03 | — | — | — | — | — | — | — | — | — | — | — | — | 1.65 ÷ 5.5 | -60 ÷ 125 | — | |||
Как измерить характеристики компонентов электронных микросхем?
Эффективная система для измерения и анализа характеристик компонентов микросхем
Комплексная система, состоящая из зондовой станции Semiprobe PS4L M6 и параметрического анализатора Keithley 4200А-SCS позволяет производить измерения характеристик как активных, так и пассивных компонентов электронных микросхем, а также математически анализировать полученные результаты (рис. 1). В зависимости от модели, измерения могут проводиться в ручном, полуавтоматическом и автоматическом режимах.
1). В зависимости от модели, измерения могут проводиться в ручном, полуавтоматическом и автоматическом режимах.
Рис.1. а) Внешний вид Semiprobe PS4L M6, б) Внешний вид Keithley 4200А – SCS
Функционал зондовой станции Semiprobe PS4L M6 в составе измерительной системы
Для проведения исследования электронная микросхема крепится на специальном столике в архитектуре зондовой станции. Все последующие перемещения микросхемы по осям X, Y, Z, а также повороты осуществляются с помощью него.
Микроскоп, которым оснащена зондовая станция, позволяет получить увеличение до 4000х. Но, как правило, при работе достаточно 200х-400х. Револьверный тип подключения объективов позволяет легко менять увеличение в процессе работы (рис.2).
Рис.2.
а) Столик, на котором размещается образец,
б) управление движением столика по осям, в) объективы
Важный момент заключается в том, что элементы, которые необходимы для анализа должны быть оторваны от цепи. А если они располагаются на внутренних слоях микросхемы, то для них нужно вывести контактные площадки.
Рис.3.
а) Вид микросхемы с подведенными зондами на экране монитора (изображение получено с помощью камеры, установленной на микроскопе), б) Общий вид микросхемы, где выделены компоненты, характеристики которых будут представлены ниже. VT1, VT2 – биполярные транзисторы n-типа, VD1 – стабилитрон, R1, R2 – резисторы
Функционал параметрического анализатора Keithley 4200А-SCS в составе измерительной системы
Когда найдена область представляющая интерес, оператор фокусируется и подводит зонды. Далее все операции осуществляются на параметрическом анализаторе Keithley 4200А-SCS.
Для каждого типа измерения создан специальный шаблон, что значительно упрощает работу. Задаются необходимые параметры для токов и напряжений, шаг и количество точек. Рабочий диапазон напряжений до 200В, токов – до 1А.
Рис.4. а) Пример шаблона с различными типами измерений характеристик для транзистора, б) примеры полученных характеристик.
Далее представлены примеры полученных характеристик для биполярных транзисторов n-типа, стабилитрона, резисторов (рис.5-8).
Рис.5. Выходные ВАХ- характеристики биполярных транзисторов n-типа а) для VT1, б) для VT2
Рис.6. Входные ВАХ- характеристики биполярных транзисторов n-типа а) для VT1, б) для VT2
Рис.7. ВАХ- характеристика стабилитрона VD1
Рис.8. Характеристики резисторов а) для R1, б) для R2
Все измерения сопровождаются таблицами, которые содержат полную информацию по каждой точке. При необходимости можно составлять собственные таблицы для совмещения графиков. Также есть возможность задавать формулы относительно измерений и сразу получать значения (рис. 9).
Рис.9. На примере резистора, мы видим графу, где представлено среднее значение сопротивления
В полностью автоматическом режиме, есть возможность запуска измерений всего дерева проекта (рис.10). Для этого необходимо сконфигурировать параметры измерений и тесты для каждого устройства в дереве и запустить проект на выполнение.
Рис.10.Пример внешнего вида дерева проекта
Стоит выделить, что количество SMU 9 шт, при этом нет ограничений на измерения по току до 1А. Установка предусилителя возможна на любой SMU. CVU до 10 МГц. Смещение +/- 30 В. Импульсный генератор работает до 800мА, минимальная ширина импульса 10 нсек. CVIV – коммутатор, который позволяет без перекоммутации (без дополнительного переподключения проводов) на зондовой станции проводить измерения вольт-амперных и вольт-фарадных характеристик. Keithley 4200А-SCS работает на Windows 10. Имеет габариты меньше, чем у аналогов.
Вывод
Keithley 4200А-SCS – отличное решение для создания, разработки, отладки и исследования новых материалов и устройств. Параметрический анализатор прекрасно справляется с такими задачами, как:
-
Проверка характеристик полупроводниковых и гибридных устройств;
-
Управление внешними устройствами, такими, как климатическая камера (для характеризации устройств при изменяющихся условиях, например, температурных) или автоматическая зондовая станция Semiprobe PS4L M6 через единый пользовательский интерфейс системы;
-
Возможно получение вольт-амперных (имеется импульсный режим) и вольт-фарадных характеристик при работе на зондовой станции Semiprobe PS4L M6.
Подходы к измерению параметров интегральных микросхем ИМС со всех сторон
Прежде всего, кому предназначены эти подходы? Вряд ли они требуются многочисленной аудитории любителей электроники, пользователей телевизоров и компьютеров. Маловероятно, что большой интерес проявят и производители этих самых телевизоров и компьютеров. Хотя им следовало бы знать о том, что бывает и, несмотря на их неведение, их напрямую касается.
В ком я уверен и для кого, в первую очередь, пишу – так это производители электронной техники специального назначения. Под техникой специального назначения понимается не только военная техника. Хотя и она тоже, но о ней говорить не будем.
Техники специального назначения хватает и в мирных областях. Мои друзья рассказывали мне такую байку. При работах на Чернобыльской АЭС после известных событий, в горячей зоне надо было что?то сделать. А зона настолько горячая, что человека туда нельзя было послать даже в тех условиях. В одной конторе на скорую руку соорудили робот?автомат, по внешнему виду больше похожий на трехколесный велосипед. Испытали, привезли. Через шахту спустили в преддверие этой самой горячей зоны, куда он должен был приехать сам и что-то там сделать. К сожалению, спуск прошел неудачно, – этот робот?автомат лег на бок. Ставили его на колеса около получаса. За это время он набрал такую дозу, что куда он поехал и что сделал, так и осталось невыясненным.
Конечно, это такой экстрим, что Бог даст, не повторится. Но знать, что творится в горячей зоне работающих в штатном режиме АЭС совсем не лишнее.
Или еще. Мало кто знает, хотя это и не секрет, что пилотируемые космические корабли летают на возможно низких орбитах высотой километров двести?триста. Чтобы минимизировать получаемую космонавтами дозу космического облучения. А на высоте 36 000 км, где годами летают спутники связи, можно нахватать такого…
О температурном режиме тоже можно было бы поведать, но тогда трудно было бы умолчать и о других воздействиях, а это не тема моей статьи.
Так вот, те, кто делают аппаратуру для ответственных и особенных целей, должны (вынуждены, обязаны) применять электронные компоненты, которые выдерживают все, что от них потребуется, и вообще имеют повышенную надежность. Соответствие критериям надежности проверяется в процессе испытаний электронных компонентов, а выявляется это посредством измерений (до и после каждого воздействия).
Теперь подходим к практическому вопросу: какие, как и в каком объёме ИМС подлежат измерению и какова должна быть в каждом случае совокупность измеряемых (тестируемых) параметров?
Для отечественных компонентов здесь вопроса нет – все написано в ТУ и ГОСТах, надо их просто честно исполнить и точка. Для иностранных микросхем никаких нормативных документов, как правило, нет (они, разумеется, есть, но недоступны). Data-sheet является некоторым аналогом РТМ и не содержит сведений, необходимых для измерений.. Содержащиеся в нем ограниченные справочные данные (типа нашего РТМ), предназначенны для тех, кто применяет ИМС, чтобы иметь представление, в каких режимах ИМС работает, и на что можно рассчитывать.
Хотя, казалось бы, чего измерять импортные ИМС? Западные производители должны делать на совесть. Они дело знают, не первый год по ISO работают. Конечно, в большей части случаев это так, но… Западные производители очень хорошо разбираются во всяких тонкостях. Если на импортную микросхему указано, что она работает в диапазоне температур 0…70 °C, то можно не сомневаться, что в этом диапазоне температур она работает. А если нужно +75? Увидите, не всякая и до +71 °C дотянет. А если, например, при -20 °C эта микросхема отказала, правильные люди скажут – “Покупайте те, для которых указано -20 °C”. А не продают! В этом случае измерения опять помогают – и найти, то, что нужно, и определить запасы, и прогнозировать надежность.
Какие ИМС измерять, таким образом, разобрались (все). Остались еще два вопроса. Как и в каком объёме. Вот эти-то вопросы главные и подлежат тщательному и внимательному рассмотрению.
Как уже сказано, для отечественных ИМС есть ТУ, в которых написано, что, когда, в каком объёме измерять или испытывать и критерии годности. Казалось бы все ясно. Но и здесь могут быть разнообразные нюансы.
Например, требуется промерить 16?ти разрядный ЦАП. По ОСТ 11.0078. Какие вопросы? Вот только в ОСТе написано, что для измерения 16?ти разрядного ЦАП требуется вольтметр с точностью не менее, чем в 4 раза лучшей измеряемой величины. Т.е. с точностью 18?ти двоичных разрядов или с погрешностью не хуже 0.00038%.
Для таких точностей применяют не проценты, а другие единицы – ppm (одна миллионная доля). 1%=10 000 ppm. А то в нулях процентов запутаться легко. В такой записи потребная точность вольтметра равна 3,8 ppm. Мало того, что эта цифра находится на пределе (даже за пределами для отечественных приборов) возможности, так еще и измерения с такой точностью идут довольно долго. Например, одну секунду. А по ОСТу надо пройтись по всем кодовым точкам ЦАПа. Итого 65535 с = 18,2 час, или почти сутки (или три рабочих смены) на одну микросхему.
Для импортных ИМС не у кого спросить – что, как и в каком объёме измерять. Это коммерческая тайна производителя. DataSheet (из которого большинство читает только то, что написано в левом верхнем углу) – это исходные данные для написания ТУ. А писать ТУ – необходимо на всякую ИМС, подлежащую аттестации. Ибо только так можно понять и согласовать что, как и в каком объёме измерять.
Вроде наступает ясность и с вопросами как и в каком объёме измерять ИМС. Надо взять ТУ – а там все написано. А если ТУ нет, то их надо написать. И возникает новый вопрос – а как написать ТУ? Подход к этому тоже понятен – в ГОСТах и ОСТах нормируются и порядок, и методы измерений, на которых зиждется разработка любого ТУ. Всего-то и надо – профинансировать ОКР, разработать ТЗ, провести испытания опытной партии и, исходя из требований ТЗ, а также по результатам испытаний, написать ТУ.
Спускаемся на ступень ниже. Допустим, деньги есть, микросхемы есть и интеллект тоже есть. Надо найти подход к написанию ТЗ (для проведения ОКР). Степень неопределенности возрастает. Конечно, в ГОСТах и ОСТах написано, какие разделы должны быть отражены в ТЗ и изложены основные требования к содержанию этих разделов. Но вот конкретные технические данные надо написать самостоятельно. И это не просто. Например, бездумно их писать на основе DataSheet – категорически неправильно и опасно. Можно загнать себя в такой тупик, выход из которого будет очень дорого стоить. Но других исходных данных на импортные ИМС нет. Поэтому рассмотрим подход к препарированию DataSheet в ТЗ.
Для примера возьмем два изделия фирма “Analog Device” AD9042 и AD9224 , – сходные по своим параметрам АЦП – и попробуем для начала составить перечень параметров, подлежащих включению в ТЗ для ИМС AD9042. Имеется 2 разновидности AD9042 – AST и AD. Вроде, они отличаются только одной строчкой DataSheet, и то не очень сильно. В одном случае (AST) – указаны предельные значения DNL при комнатной температуре и типовое значение во всем диапазоне температур, а во втором (AD) указаны предельные значения DNL и при комнатной и при предельной температуре. Интересно, что цена этих изделий составляет соответственно $40.25 и $225 (см. Digi-Key). О INL в полном температурном диапазоне (всего-то 85 °C) речь не идет не в первом, ни во втором случае.
Смотрим в DataSheet. Первый параметр – “Resolution” (разрядность). Вроде бы не худо включить в ТЗ. Но к чему это приводит? Если мы его впишем в ТЗ, то это надо будет проверять. А как? Либо найти в ГОСТах, ОСТах метод измерения этого параметра, либо расписать собственный метод. При этом то, что AD9042 – это 12-ти разрядный АЦП можно бы поверить фирме Analog Device. Я бы параметр “Resolution” в ТЗ не включал ввиду его неинформативности, и наличии вреда в виде дополнительных измерений (этот параметр скорее конструктивный).
Далее группа параметров “DC accuracy” (точность на постоянном токе). Важнейшая группа параметров, безусловно, подлежащая включению в ТЗ. Однако тут тоже скрываются некоторые “но”. Рассмотрим эту группу параметров внимательно на примере двух параметров из неё – “Offset Error” и “Offset Tempco”. Для обоих параметров в графе “Temp” (температурный диапазон) указано “Full” – полный. И, если не подходить к составлению списка параметров ТЗ очень тщательно и внимательно, можно в своём ТЗ с ходу тоже записать полный температурный диапазон. И получить необходимость всегда мерить эти параметры еще и при, по крайней мере, трех температурах. А у фирмы AD в этом DataSheet есть еще графа “Test Level” (глубина проверки). И только на 4-ой странице в уголке есть табличка:
где объясняется, что на самом деле параметр “Offset Error” измеряется на всех микросхемах только в нормальных условиях и только на небольшой выборке этот параметр поверяется при крайних значениях температур. А для параметра “Offset Tempco” вообще приводится только типовое значение. Возникает вопрос, а стоит ли нам быть святее Папы Римского и мерить все подряд, даже то, что сама фирма-изготовитель не меряет? Однозначного ответа на этот вопрос нет. И в каждом конкретном случае решать этот вопрос следует крепко подумавши, исходя из степени ответственности предполагаемого применения микросхем, имеющихся ресурсов, сложности измерений и цены, какую придется заплатить за измерения и за проблемы, возникающие как в случае проведения измерений, так и в случае отказа от них. Кроме того, при решении этого вопроса следует учесть следующие моменты. Во-первых, в соответствии с ГОСТами и ОСТами, типовое значение параметра определяется в ходе ОКР путем проверки этого параметра на установочной партии. Строится гистограмма распределения этого параметра по количеству ИМС и максимум этой гистограммы – и есть типовое значение параметра. Соответственно, включить такой параметр, для которого приведено только типовое значение в ТЗ на ОКР можно. Только не в раздел основных (сдаточных) параметров, а в список параметров, подлежащих включению в раздел “Справочные данные” ТУ. И можно даже оговорить в ТЗ, проводить перепроверку фирменных данных, или просто поверить фирме-изготовителю. Вопросы ответственности в данном случае целиком лежат на том, кто эту ИМС применил, поскольку никаких гарантий на реальное значение этого параметра никто не даст. Во-вторых. Если следовать логике фирмы-изготовителя и для какого-то параметра назначать выборочный контроль, то в этом случае необходимо прописать конкретные объёмы выборок.
Короче, для ИМС AD9042 я бы в отсутствие специальных указаний на повышенную ответственность применения выбрал бы следующий список параметров для измерений:
- “DC Accuracy”:
“Offset Error”
“Gain Error” - “Encode Input”:
“Logic “1” Current”
“Logic “0” Current” - “Digital Outputs”:
“Logic “1” Voltage”
“Logic “0” Voltage” - “Power Supply”:
“Icc (Total) Supply Current” - “Differential Nonlinearity (ENCODE=41MSPS)”
- “Integral Nonlinearity (ENCODE=41MSPS)”
с измерением всех параметров только в нормальных условиях и нормами разбраковки, взятыми из DataSheet. Исключение на нормы разбраковки составил бы последний параметр. О нем и о предыдущем параметре следует сказать особо.
В фирменном DataSheet оговорены следующие параметры, характеризующие точность этого АЦП, которые контролируются на каждом образце):
- SNR (signal-to-noise ratio – отношение сигнал/шум)
- SINAD (signal-to-noise and distortion ratio – отношение сигнал/(шум+искажения))
- Worst Spur (отношение полезной гармоники к наихудшей паразитной)
- Differential Nonlinearity (ENCODE=20MSPS) (дифференциальная нелинейность при выборке с частотой 20 мегавыборок в секунду).
Почему же предложена совсем другая система параметров точности? Потому что в существующем у нас ОСТе описаны методы измерения именно предложенных параметров. А, поскольку, параметры точности, измеряемые на максимальной частоте, отвергнуты, то предложенные параметры требуется измерять на максимальной частоте. Такой ответ, хотя и вызывает некоторое понимание, оставляет немало вопросов. Правомерна ли такая замена? Если да, то почему у них так, а у нас эдак? Какая система параметров предпочтительна?
Итак, правомерно ли? Правомерно, коли речь идет о характеристики линейности преобразования. И ненулевая интегральная нелинейность, и искажения, и паразитные гармоники порождаются нелинейностью передаточной характеристики АЦП (и ЦАПа тоже). И в этом смысле эти параметры взаимосвязаны и могут применяться в любом наборе. Что касается параметра SNR, то интегральная нелинейность его не характеризует. И, если предполагаемое применение данного образца АЦП, предъявляет жесткие требования по шуму, а само применение крайне ответственно, то промерить какой-либо параметр, эквивалентный параметру SNR необходимо. В остальных случаях вполне можно поверить фирме-производителю.
Ну а почему у нас так, а у них эдак? Ну, потому, что когда почти четверть века назад у нас писали методы измерений параметров АЦП и ЦАП параметры SNR, SINAD и Worst Spur были еще не актуальны. А потом никто ничего не хотел исправлять – больно волокитно, да и перестройка и то, что было за ней, помогли. А вообще-то все эти параметры имеют право на жизнь. Вот только какой в каком случае применять – это следует обдумать. А еще лучше не только обдумать, но и утвердить в качестве стандартизованных параметров и методов измерений к ним.
Пример с микросхемой AD9042 можно считать рассмотренным. И достаточно полно и наглядно. Зачем к рассмотрению предложена еще одна, практически такая же микросхема? А затем, чтобы показать, что информация о степени тестирования, изложенная в DataSheet это не правило, а, скорее, исключение. И определять, какие параметры фирма-изготовитель измеряет на всех образцах, а какие – только на выборке, какие во всем температурном диапазоне, а какие только при нормальной температуре, надо по наитию, имея некоторые паранормальные способности в электронике.
А для ИС AD9224 я бы предложил следующую систему параметров при аттестации:
- “Accuracy” (все параметры за исключением “No Missing Codes Guaranteed”)
- “Power Supply Rejection”
- “Internal Voltage Reference”:
“Output Voltage (1V mode)” 1000±17 mV
“Output Voltage (2V mode)” 2000±35 mV
“Load Regulation” “Power Supply”:
“IAVDD”
“IDRVDD”
Все параметры в температурном диапазоне в соответствии с DataSheet.
А теперь наверно надо совершить непосредственный подход к разработке измерения какой-нибудь микросхемы. Есть предложение начать с ADG608. Почему? Ведь все предыдущие рассуждения относились к АЦП. Начинать разборки с адаптерами с такого сложного в измерениях изделия, как АЦП не хочется. Вроде бы логично начать с ЛА3, но тут уж все слишком просто, поэтому и предложено начать с аналогового ключа – и не очень сложно, но и есть о чём поговорить.
Для начала составим список параметров, подлежащих измерениям:
- Ron – сопротивление в открытом состоянии
- Is – токи утечки закрытого и открытого канала
- Iin – входные токи цифровых входов
- Idd, Iss – токи потребления по каждому питанию.
Измерения динамических параметров не включены в этот список вслед за фирмой-изготовителем, которая гарантирует выполнение этих параметров качеством разработки.
Измерения предлагается проводить при нормальной температуре при двух напряжения питания: ±5В и +3В. Сопротивление ключа в открытом состоянии измерять при трех значениях напряжения на ключе относительно общего провода. Нормы разбраковки взять из DataSheet для каждого условия измерения.
В DataSheet на эту микросхему приведены и схемы методик измерения соответствующих параметров, (слава этому DataSheet`у!) так что задача несколько облегчается. Остается только разработать оснастку, подобрать измерительные приборы и можно мерить. В качестве комплекта измерительных приборов предлагается взять автоматизированную установку “Тестер БИС Formula 2К”. Данная установка позволяет провести измерение всех перечисленных выше параметров. Для проведения измерения большинства параметров из вышеприведенного списка достаточно подключить все цифровые входы и аналоговые входы-выходы измеряемой микросхемы на измерительные каналы тестера “Formula 2К”, и подать на измеряемую микросхему питание с измерительных источников тестера. Исключение составит измерение тока утечки канала в открытом состоянии, для измерения которого потребуются дополнительные детали.
Принципиальная схема оснастки для измерения параметров ИС ADG608 приведена на рисунке:
Рис. 1. Принципиальная схема оснастки для измерения параметров ИС ADG608
Если при изготовлении оснастки учесть требования, предъявляемые возможностью помещения измеряемой микросхемы в термокамеру, то можно обеспечить проведение измерений параметров в температурном диапазоне.
Далее, на мой взгляд, следует провести подход к методам измерения перечисленных параметров и их реализации на предложенном оборудовании.
Начать наверно можно с простого (на первый взгляд) параметра – с тока потребления.
В ГОСТ 19799?74 изложен один из методов его измерения – Метод 2570.
Посмотрим, как этот метод можно реализовать на Тестере “Formula 2К”. Самый простой вариант – подключить внутренний АЦП блока статических измерений к измерительному источнику и измерить протекающий ток. Такой подход работает, если ток потребления ИС заметно превосходит аддитивную составляющую погрешности измерительной цепи источника. В данном случае это не так. Чтобы промерить столь малый ток потребления необходимо воспользоваться измерителем статических параметров – DCM. Сделать это можно двумя способами.
Можно подключить DCM вместо источника питания, по которому необходимо произвести измерение потребления, произвести установку всех необходимых режимов, измерить ток потребления, выбрав такой диапазон тока ограничения, который обеспечит наилучшую погрешность измерения, снять входные воздействия, а затем снять питание и отключив ИС от источников и DCM, продолжить измерение остальных параметров.
А можно, в процессе проведения измерений установить требуемый для измерения тока потребления режим на DCM, подключить его параллельно измерительному источнику, по которому требуется измерить потребление, затем отключить измерительный источник, произвести измерение тока, протекающего через DCM, подключить на свое место измерительный источник, и, после отключения DCM, продолжить измерения.
Разница между предложенными способами невелика, но в некоторых случаях очень важна. Во втором варианте в процессе коммутаций DCM на измерительный источник и обратно возможно возникновение кратковременных колебательных переходных процессов, уравнивающих потенциалы на двух источниках напряжения. Однако сам процесс измерения тока потребления занимает заметно меньше времени. Если такие переходные процессы некритичны, то этот вариант предпочтителен. Если же допускается только плавный переход от одного напряжения к другому, то необходимо использовать первый вариант. К “неженкам” в первую очередь относятся ИС, изготовленные по технологии “Кремний?на?Сапфире” (КНС). Классический КМОП, а тем более, биполярные технологии (ТТЛ, ТТЛШ, большинство ОУ и компараторов) в чувствительности к таким переходным процессам замечены не были.
После такого описания простого на первый взгляд параметра, становится ясно, что простых параметров не бывает и можно перейти к более сложному – входной ток цифровых входов.
И опять есть несколько вариантов проведения измерений. Для начала самый правильный (и, понятное дело, самый сложный).
В соответствии с требованиями ОСТ при измерении входного тока низкого уровня надо на все входы, кроме измеряемого, подать высокий уровень, а на измеряемый – низкий и проверить ток, протекающий через задатчик входного сигнала. Входные сигналы можно подавать с драйверов – с устройств, обеспечивающих подачу входных сигналов при проведении функционального контроля (ФК). Для реализации такого способа подачи необходимо подготовить участок тестовой последовательности (ТП) ФК, реализующий бегущий 0 (или 1 для входного тока высокого уровня). Продвигая “0” по всем входам с помощью потактового выполнения ТП и поочередно подключая ко входу вместо драйвера DCM можно промерить все входные токи.
Вроде все ясно. Но если вдуматься, то вопросов возникает довольно много. Во-первых. Как подключать ко входу вместо драйвера DCM? Вариант первый. Отключить драйвер, затем подключить DCM. Вариант второй. Подключить к выводу ИС DCM, не отключая драйвер, а затем отключить драйвер. Обратную коммутацию проводить в обратном порядке в обоих вариантах соответственно. Недостаток первого варианта очевиден, на время пока драйвер отключен, а DCM еще не подключен, вывод болтается в воздухе. В этом нет ничего страшного для ТТЛ ИС, но совершенно не допустимо для КМОП ИС. У второго способа тоже есть недостаток. При коммутации одного источника напряжения (драйвер) параллельно с другим (DCM), поскольку точно выровнять напряжения на них нельзя, возникают переходные процессы, связанные с ограничением тока в одном из них. Этот недостаток, однако, не такой страшный, поскольку выходное сопротивление драйверов (50 Ом) достаточно большое, чтобы при правильно подобранных параметрах DCM свести переходные процессы при коммутации к минимуму. Однако сразу вылезает другая сторона этого недостатка: правильно подобрать параметры DCM для коммутации совсем не просто, а в некоторых случаях и вообще невозможно.
Но здесь есть и еще один вопрос, связанный с методом измерений. Все хорошо, пока измеряются безусловные входы. Однако, если выводы ИС двунаправленные, то все может усложниться, ибо подача на вход бегущего “0” (“1”), означает подачу на входы ИС различных кодовых комбинаций, и как отреагирует на такое издевательство сама ИС, требуется дополнительно изучать. А если входы перестанут быть входами и станут выходами? В этом случае ТП должна быть посложней, а именно, между каждым тактом бегущего “0” должна быть ТП, устанавливающая на этих выводах состояние входов. Во всяком случае, упускать это обстоятельство из вида нельзя.
Выше было отмечено, что организация бегущего “0” не единственно возможный способ. Можно использовать ТП только для установки всех “1” на входах, а бегущий “0” организовать с помощью подключаемого DCM. Или даже вовсе не использовать ТП, а все “1” подать с помощью команды установки слова на драйверах. Разница между этими двумя последними вариантами невелика и объясняется необходимостью предварительно выполненной ТП для схем с памятью состояний или отсутствием такой необходимости для простых ИС с безусловными входами. Достоинство этого метода в простоте реализации, недостаток – в наличии переходных процессов и ненормированном фронте входного сигнала, формируемого с помощью DCM. Подключение DCM к выводу ИС опять же может быть с обрывом сигнала, когда вход ИС какое-то время висит в воздухе и безобрывным. Однако этот метод проще и поэтому именно такой метод в безобрывном варианте реализован в методах табличного редактора, причем возможно как использование подготовительной ТП, так и отсутствие таковой. Опробование этого метода показало, что для большинства ИС простой логики, таких как стандартный КМОП и ТТЛ, проблем не возникает. Этот опыт, однако, не гарантирует успехов в особых случаях например ИС КМОП КНС или сложных микропроцессоров.
Ну а в данном конкретном случае (ИС ADG608) я бы воспользовался самым простым безобрывным методом без ТП.
Далее для рассматриваемой ИС идут параметры утечки закрытого и открытого канала. Утечки закрытого канала по сути ничем не отличаются от входных токов цифровых входов, разве что ОСТовские методы применять к этим токам надо поняв, как организовывать бегущий “0” (“1”). Измерение тока утечки открытого канала несколько отличается от простого измерения входного тока и будет подробно рассмотрено позднее.
Следует помянуть, что для цифровых ИС необходимо проверить еще несколько параметров: выходное напряжение (низкого и высокого уровня), максимальный выходной ток и пороги входных напряжений. Параметры расположены в порядке частоты встречи. Выходные уровни – обязательно, входные пороги – почти никогда. Все изложенное по поводу измерения входных токов почти в полной мере относится и к перечисленным параметрам. Разве что обрывный метод коммутации при измерении выходных уровней не только допустим, но и предпочтителен.
Не описанными остались временные параметры и функциональный контроль, ну да об этом в другой раз.
А сейчас о токе утечки открытого канала ИС ADG608. Как уже отмечалось выше в DataSheet на эту ИС приведены методы измерения параметров, что выгодно отличает данный DataSheet от многих других. Вот какие данные там приведены по поводу измерения тока утечки открытого канала:
На первый взгляд все ясно. Но только на первый. Если внимательно и вдумчиво рассмотреть эту схему, то станет ясно, что Vs должен быть равен VD с очень высокой точностью (с точностью до нескольких микровольт), т.е. это должен быть один и тот же источник. Но, к сожалению, в тестере “Formula 2К” не предусмотрена возможность подать напряжение от какого-нибудь источника (в том числе и DCM) к двум нагрузкам, а мерить протекающий ток только по одной нагрузке. Эта ситуация прекрасный пример того, как можно с помощью измерительного адаптера обеспечить измерение параметров методом, не заложенным в возможности тестера. Для измерения тока утечки открытого канала необходимо провести следующие действия (нумерация элементов дана в соответствии с Рис. 1):
- Подать на каналы 1…4 нулевой уровень от драйверов.
- Подать питание на измеряемую ИС от VCC и VEE.
- Подать питание на измеряемую ИС от VCC и VEE.
- Подать на канал 1 от драйвера уровень +2.4 В, воспользовавшись дополнительной парой уровней драйвера.
- С помощью команд “Connect pin-to-pin” и “Connect pin to drivers” с одного из драйверов подать требуемый уровень на каналы 5…12 и 14 тестера.
- Подключить DCM к VCD.
- Произвести замер напряжения на DCM.
- Установить нулевое напряжение на всех каналах тестера.
- Отключить источники питания и DCM.
- Отключить все каналы тестера.
- Вычислить искомое значение тока утечки открытого канала:
ID(ON)[мкА]=V(DCM)[В]/KU,
Где ID(ON)[мкА] – измеренное значение тока утечки,
V(DCM)[В] – напряжение, измеренное DCM,
KU – коэффициент усиления дифференциального усилителя U2.
Из описанной процедуры ясно, почему выход дифференциального усилителя подключен к источнику VCD – вовсе не для того, чтобы подать питание на выход, а для того, чтобы исключить конфликт, возникающий при подаче двух разных сигналов по одному проводу. Необходимо объединить все каналы с помощью общей шины платы PIN на один драйвер для подачи входного воздействия. И, если воспользоваться каналом, по той же шине платы PIN подать сигнал с выхода дифференциального усилителя на измерительную шину тестера. Можно, правда, воспользоваться каналом дополнительной платы PIN, в этом случае конфликта тоже не будет, но в этом случае требуется привлечение дополнительного оборудования, а это не есть красиво.
Не рассмотренным остался один параметр – Ron.
Как и со всеми остальными параметрами, с этим параметром тоже есть не сразу заметные вопросы. Казалось бы чего такого – померить величину резистора. Берем источник тока, запускаем этот ток в измеряемый резистор, меряем падение напряжения, делим одно на другое – и готово. Но беда в том, что сопротивление открытого ключа КМОП схемы – штука весьма нелинейная, да еще по многим параметрам. Поэтому измерять ее надо в нескольких режимах, да аккуратно. И вот задача: на одном конце резистора вольт эдак 5, на другом – милливольт на 30 меньше, а подвешенного вольтметра нет, т.е. можно измерять только напряжение относительно общего провода. По общепринятым метрологическим нормам погрешность разности двух измерений равна сумме погрешностей каждого измерения. Если погрешность нашего вольтметра составляет около 1% (а это весьма неплохо), то погрешность разности в 30 мВ между двумя замерами напряжений около 5 В составит порядка 100 мВ, т.е. такие измерения несостоятельны. Можно, конечно, поступить как и с предыдущим параметром, а именно добавить еще один дифференциальный усилитель и мерить падение напряжения на открытом ключе с его помощью. Можно и ничего страшного. Но при использовании Тестера “Formula 2К” можно поступить и по-другому. Для некоторых (наиболее важных) диапазонов DCM введена еще одна составляющая погрешности (помимо аддитивной и мультипликативной) – погрешность интегральной нелинейности. Общепринятый параметр для АЦП и ЦАП, но в метрологии почему-то практически не применяется. Данная составляющая погрешности указывает, насколько результаты всех измерений данным прибором отличаются от идеальной прямой. Если рассмотреть идеализированный вольтметр с нулевой интегральной нелинейностью, то все измерения такого вольтметра будут описаны идеальной прямой типа:
U=AV+B, где
V – измеряемое напряжение,
U – показание вольтметра,
а А и В – некоторые коэффициенты.
За счет того, что вот эти самые А и В известны не совсем точно, каждое отдельное измерение имеет некоторую погрешность, но, поскольку известно что интегральная нелинейность равна нулю, то для всех замеров эти А и В постоянны, и, следовательно, погрешность разности будет содержать только мультипликативную составляющую, определяемую погрешностью величины А. Если же погрешность интегральной нелинейности отлична от нуля, то погрешность разности будет содержать не только мультипликативную составляющую, но и некоторую аддитивную составляющую, которая с очевидностью равна погрешности интегральной нелинейности.
В данном случае-примере (ADG608) погрешность измерения падения напряжения на открытом ключе составит менее 3 мВ, что вполне приемлемо. А это значит, что никаких дополнительных телодвижений кроме замеров с помощью DCM поданного от драйвера напряжения на один вывод открытого ключа и образовавшегося потенциала на другом выводе того же ключа (на DCM должен быть установлен один и тот же ток ограничения) делать не надо.
Полный текст измерительной программы для измерения параметров этой микросхемы в данной статье не приводится по двум причинам. Во-первых, это коммерческий продукт. А во-вторых, здесь приведено столько поясняющей информации, что любой инженер-электронщик, как следует изучивший документацию на Тестер “Formula 2К”, сможет такую программу написать в своей излюбленной манере.
Чтобы окончательно разделаться с аналоговыми ключами рассмотрим как преобразовалась под присмотром представителей Заказчика система параметров ИС ADG408 (более высоковольтный, но более медленный старший собрат ИС ADG608) в систему параметров её аналога – ИС 590КН6. Ну всякие там токи утечки – это понятно. Только один нюанс. На закрытом канале ток утечки меряется при полном напряжении, да еще с 10%-ной надбавкой – это 33 В. При обеспечении этого измерения на Тестере “Formula 2К” тоже необходимо извращаться на адаптере. Ну да ладно. А вот ток утечки открытого канала – отсутствует. Наверно при написании ТУ на ИС 590КН6 его откинули из-за трудности измерения.
Но это все цветочки. Ягодки же такие. Для ИС 590КН6 по ТУ не меряется сопротивление открытого ключа! Зато: меряется время включения (только!) и проводится нечто, именуемое функциональный контроль (ФК). Т.е. на определенной нагрузке проверяется появление/пропадание заданного входного сигнала. Все это следствие нежелания разрабатывать новое оборудование для непростых измерений, негибкого применения ГОСТов и ОСТов и нежелание представителя Заказчика вникнуть в DataSheet поглубже, а также нежелание ГК (главного конструктора разработки) зарабатывать себе лишнюю головную боль, убеждая представителя Заказчика, что ГОСТы и ОСТы и DataSheet, а также полнота производственного контроля и возможности оборудования – вещи взаимосвязанные и требующие творческого подхода и не всегда стандартных решений.
Например, что такое ФК все знают и всем все понятно, пока не копнешь поглубже. А вообще-то эта тема также заслуживает обсуждения и осмысления. Но как-нибудь при случае в другой раз.
Автор:form.ru
Интернет справочник основных параметров микросхем. Цифровые интегральные микросхемы
Справочник «микросхемы современных телевизоров». В этом справочном пособии собраны данные о наиболее распространенных интегральных микросхемах, которые применяются в современной телевизионной технике. В книге представлена справочная информация о более чем 100 микросхемах таких известных фирм-производителей, как SAMSUNG, SANYO, SONY, SIEMENS, MATSUSHITA, PHILIPS, SGS-THOMSON и других.
Формат книги DjView. Размер архива – 3,29Mb. СКАЧАТЬ
Справочник «микросхемы для современных мониторов». Данная книга является справочным пособием по микросхемам для современных LCD и CRT мониторов. В ней приведена исчерпывающая информация о 150 микросхемах ведущих производителей полупроводниковых компонентов для мониторов.
Формат книги DjView. Размер архива – 5,77Mb. СКАЧАТЬ
Справочник «отечественные транзисторы для бытовой, промышленной и специальной аппаратуры». В этом справочнике представлена полная информация о номенклатуре, изготовителях, параметрах, корпусах и аналогах 5000 наименований транзисторов!
Формат книги DjView. Размер архива – 16,4Mb СКАЧАТЬ
Сборник их 3х справочников по импортным микросхемам, транзисторам, диодам, тиристорам и SMD компонентам. Книга 1 из 3х . В этом справочнике представлена информация по радиоэлектронным компонентам зарубежных производителей с буквенным индексом от A до R . Приводятся характеристики, цоколевка, аналоги и производители компонентов.
Размер файла – 198Mb. Формат книги DjView. Скачать с Deposit Files
Справочник по импортным микросхемам, тиристорам, диодам, транзисторам и SMD компонентам. Книга 2 из 3х . В этом справочнике представлена информация по радиоэлектронным компонентам зарубежных производителей с буквенным индексом от R до Z .
Размер файла – 319Mb. Формат книги DjView. Скачать с Deposit Files
Справочник по импортным микросхемам, тиристорам, диодам, транзисторам и SMD компонентам. Книга 3 из 3х . В этом справочнике представлена информация по радиоэлектронным компонентам зарубежных производителей с цифровым индексом от 0 до 9 .
Размер файла – 180Mb. Формат книги DjView. СКАЧАТЬ
Справочник по активным SMD компонентам. Приводятся SMD коды для 33 тысяч транзисторов, тиристоров, микросхем и диодов, типовые схемы включения SMD микросхем, маркировка, характеристики, замена.
Размер архива – 16Mb. Формат книги DjView. СКАЧАТЬ
Справочник «транзисторы и их зарубежные аналоги» том 1. В первом томе справочника приводятся электрические и эксплуатационные характеристики полупроводниковых приборов – полевых и биполярных транзисторов малой мощности. Даются классификация и система обозначений, основные стандарты для описанных в справочнике приборов. Для конкретных типов приборов приводятся сведения об основном назначении, габаритных и присоединительных размерах, маркировке, предельных эксплуатационных режимах и условиях работы. В приложении даются зарубежные аналоги транзисторов, помещенных в справочнике.
Формат книги DjView. Размер архива – 6,19Mb СКАЧАТЬ
Справочник «транзисторы и их зарубежные аналоги» том 2. Во втором томе справочника приводится информация по низкочастотным биполярным транзисторам средней и большой мощности с указанием их зарубежных аналогов.
Формат книги DjView. Размер архива – 5,62Mb. СКАЧАТЬ
Справочник «транзисторы и их зарубежные аналоги» том 3. В третьем томе приводится справочная информация по полевым и высокочастотным биполярным транзисторам средней и большой мощности с указанием их зарубежных аналогов.
Формат книги DjView. Размер архива – 6,28Mb . СКАЧАТЬ
Справочник «маркировка радиодеталей» том 1. В книге приведены данные по буквенной, цветовой и кодовой маркировке компонентов, по кодовой маркировке зарубежных полупроводниковых приборов для поверхностного монтажа (SMD). Приведены рекомендации по использованию и проверке исправности электронных компонентов.
Формат книги DjView. Размер архива – 8Mb СКАЧАТЬ
Справочник «маркировка радиодеталей» том 2. В этой книге читатель найдет много полезной информации по маркировке микросхем, некоторых типов полупроводниковых приборов, установочных и коммутационных изделий и много другой полезной информации.
Формат книги DjView. Размер архива – 3,95Mb СКАЧАТЬ
Справочник «маркировка радиодеталей». В книге описана система маркировки отечественных и зарубежных: резисторов, конденсаторов, индуктивностей, кварцевых резонаторов, пьезоэлектрических и ПАВ-фильтров, полупроводниковых приборов, SMD-компонентов, микросхем. Описаны особенности тестирования электронных компонентов.
Формат книги DjView. Размер архива – 3,60Mb СКАЧАТЬ
Справочник по микросхемам для импортных телевизоров. В книге на Русском языке приводятся структурные схемы и назначение выводов более трехсот микросхем, применяемых в европейских и восточно-азиатских цветных телевизорах. Описание каждого прибора сопровождается функциональными диаграммами и характеристиками.
Формат книги DjWiev. Размер архива – 16Mb СКАЧАТЬ
Справочник по микросхемам для аудио и радиоаппаратуры: генераторы, ключи и переключатели, УНЧ, малошумящие и предварительные усилители, операционные усилители, регуляторы громкости и тембра, схемы управления индикаторами. В книге представлены основные особенности, цоколевки, структурные схемы и типовые схемы применения свыше 300 типов микросхем для аудиотехники.
Формат книги DjWiev. Размер архива – 10,7Mb СКАЧАТЬ
Справочник по интегральным микросхемам для промышленной электронной аппаратуры. В книге приведены условные обозначения, электрические параметры, структурные схемы, функциональное назначение (цоколевка) и конструкции корпусов широко распространенных зарубежных аналоговых и цифровых микросхем.
Формат книги DjWiev. Размер архива – 2,68Mb СКАЧАТЬ
Лучший в Европе справочник по УНЧ . В нем обобщены и систематизированы сведения о большинстве ИМС УНЧ в интегральном исполнении, выпускаемых мировыми производителями. Приведены наиболее важные характеристики микросхем, типы корпусов, цоколевка, внешний вид, аналоги, производители, функциональное назначение .
Формат книги DjWiev. Размер архива – 19,9Mb СКАЧАТЬ
Справочник по интегральным микросхемам для телевидения. В книге дан обзор интегральных микросхем, применяемых в современных телевизионных приемниках, видео- и аудиотехнике. Приведены основные параметры и характеристики микросхем, блок-схемы внутренней структуры и типовые схемы их включения.
Формат книги DjWiev. Размер архива – 2,30Mb СКАЧАТЬ
В настоящем справочнике приведены перечни различных классов отечественных транзисторов и их зарубежных аналогов с указанием фирм-изготовителей, а так же зарубежных транзисторов и их отечественных аналогов. Для удобства работы книга разделена на две части. В первой части приведены зарубежные аналоги отечественных транзисторов, которые выстроены в алфавитно-цифровой последовательности. Во второй части приводятся отечественные аналоги зарубежных транзисторов, которые так же выстроены в алфавитно-цифровой последовательности.
Рассмотрены свойства и особенности биполярных и полевых транзисторов (с p-n-переходом, МОП, биполярных транзисторов со статической индукцией и с изолированным затвором), предназначенных для применения в бытовой, промышленной и специальной аппаратуре, приведены их электрические параметры, области применения, стандартизованные корпуса и зарубежные аналоги, а также указаны предприятия-изготовители. В справочном пособии представлена информация об особенностях применения, параметрах и характеристиках биполярных и полевых транзисторов, изготовленных в странах СНГ и Балтии.
В книге приведено самое полное описание устройств на полевых транзисторах. Особое внимание уделено новым классам этих приборов – мощным полевым транзисторам различного типа, IGBT и интегральным микросхемам на их основе. Даны основы теории, расчета и описание работы самых различных схем на полевых транзисторах: ключей, электронных регуляторов, импульсных и резонансных источников электропитания, высокоскоростных импульсных устройств, формирователей мощных импульсов, усилителей и генераторов различных частот.
Книга является вторым, исправленным и дополненным изданием выпуска, посвященного микросхемам для линейных источников питания. По сравнению с первым изданием введено большое дополнение, посвященное современным микросхемам для линейных источников питания ведущих зарубежных фирм, доступным на Российском рынке, а также исправлены все замеченные опечатки, внесены сведения о новых приборах. Для специалистов в области проектирования, эксплуатации и ремонта практически любых изделий радиоэлектроники, а также широкого круга радиолюбителей и студен юв технических ВУЗов.
Эта книга, оставаясь в рамках серии “Интегральные микросхемы”, является попыткой “быстрого реагирования” на постоянно возрастающие потребности в информации рынка электронных компонентов. В ней приводятся подробные технические данные некоторых новых для России микроэлектронных изделий, а также путеводитель по продукции фирмы “Burr-Brown” и, соответственно, торговые марки и адреса производителей описанных приборов. Для специалистов в области радиоэлектроники, студентов технических вузов и широкого круга читателей интересующихся достижениями электроники.
Наиболее полный и подробный справочник по всем внутренним и внешним интерфейсам персонального компьютера последовательного и параллельного портов, инфракрасного порта, IDE, SCSI, шинам ISA, EISA, MCA, VESA, PCI, AGP, PC-Card (PCIMCIA) и др В книге приводятся полные описания и спецификации интерфейсов от адресов ввода/вывода и прерываний до уровней сигналов и контактов разъемов Эта книга посвящена интерфейсам, позволяющим подключать к персональным (и не только) компьютерам разнообразные периферийные устройства (ПУ) и их контроллеры.
В книге описаны терминология, концепции, технологии и устройства, которые ис пользуются во всем мире для передачи данных и голосовых сигналов. Она написана та ким образом, чтобы материал был понятен даже тем читателям, которые мало знают о телекоммуникациях, но при этом книга может стать хорошим техническим руководством и для опытных инженеров. Эта книга призвана стать справочным руководством к широкому диапазону технологий, применяемых в современных телекоммуникационных сетях. Материал, представленный в книге, не ограничивается лишь североамериканскими технологиями.
Справочник является логическим продолжением аналогичного издания 1997 г. Он устанавливает связь между типом отечественной микросхемы, ее зарубежным аналогом, функциональным назначением и производителем. В нем перечислены все когда-либо разработанные и произведенные на территории СНГ микросхемы – всего около 8000 типов. По сравнению с первым изданием, значительно расширен перечень приборов, уточнены аналоги многих микросхем и добаалена информация о состоянии производства приборов. Кроме этого, добавлены таблица функционального назначения микросхем и таблица всех типономиналов, рекомендуемых к применению приборов.
Книга начинает серию энциклопедических справочников по современной элементной базе электронной техники. Отличительной особенностью справочника является широкое использование ссылок на электронные базы компонентов фирм-производителей. В книге приводятся все необходимые сведения, позволяющие понять принципы работы, систему классификации, терминологию, типовые параметры и схемы включения, а вся фактическая информация о конкретных микросхемах содержится в виде ссылок на сайты производите лей. Такой подход позволил в книге небольшого формата разместить данные о 10 видах современных БИС: ЦАП, АЦП, синтезаторах частот, ИМС памяти, микропроцессорах, мик роконтроллерах, цифровых сигнальных процессорах, программируемых логических матри цах, схемах с квадратурной обработкой и кодеках.
В настоящем справочнике приведены схемы подключения и параметры более чем 3500 микросхем усилителей мощности низкой частоты, выпускаемых ведущими фирмами-производителями – ” ECG-Philips, Matsushita-Panasonic, National Semiconductors, NTE, Philips, RCA, Sanyo, Siemens, SGS-Thomson, Telefunken-Temic, Toshiba и др. Здесь представлены как стандартные, так и нестандартные (мостовые) схемы включения. Микросхемы, имеющие идентичную электрическую схему включения, собраны в одном разделе, так как практически они являются аналогами.
Предисловие
РАЗДЕЛ I ЦИФРОВЫЕ МИКРОСХЕМЫ
Глава 1. Общие сведения о цифровых микросхемах средней степени интеграции
1.1. Классификация
1.2. Основные характеристики и параметры ЛЭ
1.3. Сравнение обобщенных параметров цифровых микросхем
1.4. Типовые корпуса микросхем
Глава 2. Микросхемы на основе ТТЛШ
2.1. Маломощные ТТЛШ ИС
2.2. Базовый логический элемент ИС533 (К533, КМ533, К555, КМ555)
2.3. Базовый логический элемент ИС1533 (КР1533)
2.4. Быстродействующие ТТЛШ ИС
2.5. Базовый логический элемент ИС530 (К530, КМ530, КМ531, КР531)
2.6. Базовый логический элемент ИС1531 (КР1531)
2.7. Логические элементы
2.8. Магистральные элементы
2.9. Мультиплексоры
2.10. Шифраторы и дешифраторы
2.11. Триггеры
2.12. Регистры
2.13. Счетчики
2.14. Арифметическо-логические устройства
2.15. Приемопередатчики
2.16. Формирователи
2.17. Мультивибраторы
2.18. Зависимость параметров микросхем от режимов работы и условий эксплуатации
Глава 3. Микросхемы на основе КМОП-технологии
3.1. Основные характеристики ИС К564
3.2. Сопряжение ИС К564 с другими сериями
3.3. Базовые логические элементы
3.4. Комбинационные устройства
3.5. Последовательностные устройства
Глава 4. Микросхемы на основе ЭСЛ
4.1. Быстродействующие микросхемы
4.2. Базовый логический элемент ИС К500 (100, К100,500)
4.3. Базовый логический элемент ИС К1500 (1500)
4.4. Особенности применения ЭСЛ ИС
Глава 5. Микросхемы на основе арсенида галлия
5.1. Базовый логический элемент сверхбыстродействующих ИС К6500
5.2. Логические элементы, триггер, счетчики, регистры
РАЗДЕЛ II. ЦИФРОВЫЕ БАЗОВЫЕ МАТРИЧНЫЕ КРИСТАЛЛЫ
Глава 6. Проектирование полузаказных БИС на основе базовых матричных кристаллов
6.1. Классификация методов проектирования специализированных БИС
6.2. Особенности конструкции БК
6.3. Терминология, основные параметры и характеристики БК
6.4. Основные направления развития БМК
6.5. Организация разработки БИСМ
6.6. Применение матричных БИС в унифицированных узлах РЭА
Глава 7. Цифровые базовые матричные кристаллы на основе эмиттерно-связанной логики
7.1. Базовые матричные кристаллы типа К1520ХМ1, К1521ХМ1
7.2. Базовый матричный кристалл типа К1520ХМ2
7.3. Базовый матричный кристалл типа К1572ХМ1
Глава 8. Цифровые базовые матричные кристаллы на основе транзисторно-транзисторной логики с диодами Шатки
8.1. Базовый матричный кристалл типа К1527ХМ1
8.2. Базовый матричный кристалл типа К1548ХМ1
Глава 9. Цифровые базовые матричные кристаллы на основе я-МОП-структур типа К1801ВП1
Глава 10. Цифровые базовые матричные кристаллы на основе КМОП-структур
10.1. Базовые матричные кристаллы типа К1806ВП1, КР1806ВП1
10.2. Базовый матричный кристалл типа К1515ХМ1
Глава 11. Цифровые базовые матричные кристаллы на новых материалах типа К6501ХМ1
РАЗДЕЛ III. ПРОГРАММИРУЕМЫЕ ПОТРЕБИТЕЛЕМ ЛОГИЧЕСКИЕ МИКРОСХЕМЫ
Глава 12. Ресурс программируемых логических микросхем
Глава 13. Программируемые логические микросхемы
13.1. Микросхемы К556РТ1, К556РТ2
13.2. Микросхема КМ1556ХП4
13.3. Микросхема КМ1556ХП6
13.4. Микросхема КМ1556ХП8
13.5. Микросхема КМ1556ХЛ8
13.6. Программируемая логическая интегральная микросхема с ультрафиолетовым стиранием информации
13.7. Расширение логических возможностей ПЛМ
Приложение 1. Принципиальные электрические схемы и условные графические обозначения библиотечных ФЯ БМК типа К1521ХМ1
Приложение 2. Принципиальные электрические схемы и условные графические обозначения библиотечных ФЯ БМК типа К1520ХМ1
Приложение 3. Принципиальные электрические схемы и условные графические обозначения библиотечных ФЯ БМК типа К1520ХМ2
Приложение 4. Принципиальные электрические схемы и условные графические обозначения библиотечных ФЯ БМК типа К1572ХМ1
Приложение 5. Принципиальные электрические схемы и условные графические обозначения библиотечных ФЯ БМК типа К1527ХМ1
Приложение 6. Принципиальные электрические схемы и условные графические обозначения библиотечных ФЯ БМК типа К1548ХМ1
Приложение 7. Условные графические обозначения библиотечных ФЯ БМК типа К1801ВП1
Приложение 8. Принципиальные электрические схемы и условные графические обозначения библиотечных ФЯ БМК типа К1515ХМ1
Приложение 9. Условные графические обозначения библиотечных ФЯ БМК типа К6501ХМ1
Список литературы
Название: Справочник – Популярные цифровые микросхемы.
Приведены сведения о трех самых распространенных в радиолюбительской практике видах цифровых микросхем: ТТЛ, КМОП и ЭСЛ. Кратко рассмотрены основы их схемотехники, показаны структуры, цоколевки и дано описание работы более 300 типов массовых цифровых микросхем: логических элементов, триггеров, регистров, счетчиков, мультиплексоров, арифметических и др. Даны рекомендации по их применению. Для подготовленных радиолюбителей и специалистов народного хозяйства, разрабатывающих и применяющих импульсно-цифровую аппаратуру.
Предлагаемая книга посвящена схемотехнике самых массовых серий микросхем – цифровых малой и средней степени интеграции. Известно, что в 70-е – 80-е годы в аппаратуре доминируют три вида таких микросхем; ТТЛ, КМОП и ЭСЛ. Их выпускают сотнями миллионов штук в год. Возможно, многие из них будут изготавливать до конца столетия.
В каждом из трех видов микросхем существуют преемственно развивающиеся серии. Имея описание микросхемы, можно реализовать ее свойства полностью. Каждая группа микросхем (к примеру, счетчиков, регистров) имеет сейчас много схемотехнических применений. Варианты схемотехники отображают* как ход развития микросхем, так и расширение запросов потребителей. Вдумчивый читатель может проследить путь развития схемотехники от простейших микросхем до современных и перспективных. Кроме того, полезно сравнить, как исходные устройства оптимизируются и трансформируются под схемотехнику ТТЛ, КМОП и ЭСЛ.
1. ЦИФРОВЫЕ МИКРОСХЕМЫ ТТЛ
1.1. Общие сведения об элементах ТТЛ
1.2. Схемотехника элементов ТТЛ
1.3. Традиционные серии ТТЛ
1.4. Перспективные серии ТТЛ
1.5. Буферные и разрешающие элементы ТТЛ
1.6. Схемотехника элементов И, ИЛИ, И/ИЛИ
1.7. Микросхемы ТТЛ: И,И, ИЛИ, И/ИЛИ, расширители
1.8. Автогенераторы на элементах ТТЛ
1.9. Логические элементы – триггеры Шмитта
1.10. Исключающее ИЛИ
1.11. Триггерные схемы
1.12. RS- и D-триггеры
1.13. JK-триггеры
1.14. Счетчики ТТЛ
1.15. Регистры ТТЛ
1.16. Дешифраторы и шифраторы ТТЛ
1.17. Мультиплексоры ТТЛ
1.18. Сумматоры ТТЛ
1.19. Оперативные и постоянные запоминающие устройства ТТЛ
1.20. Узлы вычислительных устройств
1.21. Ждущие мультивибраторы и автогенераторы
2. ЦИФРОВЫЕ МИКРОСХЕМЫ КМОП
2.1. Устройство и свойства логического элемента КМОП
2.2. Основные логические элементы И, ИЛИ, Z
2.3. Микросхемы с инверторами и их применение
2.4. Схемы генераторов и преобразователей
2.5. Преобразователи уровней логических сигналов
2.6. Коммутаторы цифровых и аналоговых сигналов
2.7. Триггерные микросхемы КМОП
2.8. Счетчики-делители КМОП
2.9. Регистры КМОП
2.10. Дешифраторы КМОП
2.11. Арифметические схемы КМОП
2.12. Микросхемы ФАП и мультивибраторы
3. ЦИФРОВЫЕ МИКРОСХЕМЫ ЭСЛ
3.1. Схемотехника логических элементов
3.2. Комбинаторные микросхемы серии К500
3.3. Триггеры, счетчики и регистры серии К500
3.4. Элементы вычислительных устройств из серии К500
3.5. Комбинаторные микросхемы серии К1500
3.6. Триггеры и регистры серии К1500
3.7. Узлы вычислительных устройств серии К1500
Приложение
Список литературы
Бесплатно скачать электронную книгу в удобном формате, смотреть и читать:
Скачать книгу Справочник – Популярные цифровые микросхемы – Шило В.Л. – fileskachat.com, быстрое и бесплатное скачивание.
Скачать djvu
Ниже можно купить эту книгу по лучшей цене со скидкой с доставкой по всей России.
| Наименование и разновидности | Тип корпуса | Аналог | Функциональное назначение | Описание | Применение |
| 1. Микросхемы для работы с сетевым напряжением | |||||
| К1182ГГ2Р | PowerDIP (12+4) | без аналога | Полумостовой высоковольтный автогенератор. ЭПРА компактных люминесцентных ламп мощностью до 15 Вт. | ||
| К1182ГГ3Р | DIP-8 | без аналога | Полумостовой высоковольтный автогенератор,преобразующий выпрямленное сетевое напряжение в высокочастотное напряжение 30-50 кГц для гальванически развязанных вторичных источников питания мощностью до 12 Вт и галогенных ламп. | ||
| К1182КП1П, К1182КП1(А-Г)П |
TO-92 | К1100Е70 | Диодный симистор с фиксированным напряжением открывания для стартовых цепей автогенераторных схем, мощных тиристоров (симисторов), для коммутации тока через нити подогрева маломощных люминесцентных ламп до момента зажигания лампы. Uon=8.5В-105B, Imax=1A | ||
| К1182КП2Р | TO-126 | без аналога | Фазовый регулятор,функционально аналогичный газоразрядному стартеру люминесцентных ламп. Uon>500 B, Imax=1A. | ||
| К1182ПМ1Р | PowerDIP (12+4) | без аналога | Фазовый регулятор для плавного вкл./выкл. ламп накаливания, регулировки яркости, управления мощными симисторами U=240VAC, Uост.=2.0VAC, Pнагр.=150W. | ||
| К1182ПМ1Р1 | DIP-8 | ||||
| К1182ПМ1Т | SO-8 | ||||
| К1182СА1Р | DIP-16 | без аналога | Двухфазный контроллер сетевого питания. Автоматическое прерывание сетевого питания для защиты человека от поражения электрическим током, для защиты нагрузки от перенапряжения. Ui=400 B, tсраб.=10 мс, dIсраб.=5 мА, Iупр.=300 мА | ||
| 2. Микросхемы для импульсных и линейных источников питания | |||||
| К1033ЕУ(15,16)(А,Б,В,Г)Р | DIP-8 | UC3842, UC3843, UC3844, UC3845 | ИС управления однотактными ИСН, Usmax=30В, Is=1A, fsmax =500кГц. | ||
| К1033ЕУ(15,16)(А,Б,В,Г)Т | SO-8 | ||||
| К1033ЕУ(15,16)(В,Г)Р1 | Д8К-2Н | ИС управления однотактными ИСН, Usmax=30В, Is=1A, fsmax =500кГц. | |||
| К1033ЕУ(15,16)(В,Г)Т1 | Н02.8-2В | ||||
| К1156ЕК1(А,Б)П, К1156ЕК3.3(А,Б)П, К1156ЕК5(А,Б)П, К1156ЕК12(А,Б)П |
TO-220-5 | LM2596 | Импульсный стабилизатор напряжения на нагрузку до 3А. Модификация с фиксированным выходным напряжением 3.3В,5В,12В и с программируемым выходным напряжением от 1,2В до 37В. | ||
| K1156ЕН1 | TO-220-5 | LM2925 | “Low drop” стабилизатор положительного напряжения на 5В со схемой “Сброс” Uвх.max=26В, Uвых.=5В, Iн=0.5А, Uпдмин.=0.6В. | ||
| K1156ЕН5ВП, K1156ЕН5ДП |
TO-220-5 | без аналога | Регулируемый ЛСН с низким Uпдмин. Uвх.max=26В , Uвых.=1.5-15В, Iн=0.5А, Uпдмин.=0.6В. | ||
| K1156ЕН5ДУ | SLCC5/5 | ||||
| K1156ЕР2У | SLCC5/5 | PQ30RV1/2 | Регулируемый ЛСН с малым падением напряжения и низкой потребляемой мощностью. Uвх.=4.5-35В, Uвых.=1.5-30В, Iвых.=2А. | ||
| К1156ЕР5П | TO-92 | TL431A, К142ЕН19П |
Регулируемый стабилизатор напряжения параллельного типа. Опорное напряжение – 2,47-2,52В, напряжение на катоде не более 36В. | ||
| К1156ЕР5Т | SOT-89 | ||||
| К1156ЕУ1Т | ТАКТ-256 | mA78S40 | ИС управления и регулирования импульсными источниками питания. Us=3-40В B, Is=1А, fsmax=0.1-100кГц. |
||
| К1156ЕУ(2,3)(А,Б,В,Г)Р | DIP-16 | UC3825, UC3823, UC2825, UC2823 |
ИС управления двухтактными ИСН. Us=30В , Is=1.5А, fs=1MГц,tзад=50нс. | ||
| К1156ЕУ(2,3)(А,Б,В,Г)Т | ТАКТ-256 | ||||
| К1156ЕУ5(В)Р | DIP-8 | MC34063 | DC-DC конвертер. Us=3-40В, Is=1.5А, fs=0.1-100кГц. | ||
| К1156ЕУ5(В)Т | SO-8 | ||||
| К1156ЕУ5Р1 | Д8К-2Н | ||||
| К1156ЕУ5Т1 | Н02.8-2В | ||||
| KР1158ЕН(3-15)01(А,Б) | TO-251 | L4945, LM2930, LM2931 |
“Low drop” стабилизатор положительного фиксированного напряжения. Uвых.=3-15В с дискретностью 0.1В | ||
| КФ1158ЕН(3-15)01(А,Б) | TO-252 | ||||
| KР1158ЕН(3-15)(В,Г) | TO-220 | ||||
| KФ1158ЕН(3-15)(В,Г) | TO-263 | ||||
| K1158ЕН(3-15)(Ж,И)П | TO-220 | ||||
| K1158ЕН(3-15)(Ж,И)Т3 | 4116.4-6 | ||||
| K1158ЕН(3-15)(Ж,И)У | SMD-05 | ||||
| К1278ЕНхх(Б,В,Г,Д,Ж,И,К,Л)П | TO-220 | IRU1117-хх, APL1117-хх, CS5203-х, CS5205-x |
“Low drop” cтабилизаторы положительного фиксированного напряжения. (Б, В, Г, Д – аналоги IRU1117-хх, APL1117-хх, CS5203-х, CS5205-x) |
||
| К1278ЕНхх(Ж,И,К,Л)Т3 | 4116.4-6 | Входное напряжение: 7В (Ж)/12В (И,К,Л), ток нагрузки 0.8/1.0/3.0/5.0А | |||
| К1278ЕНхх(Ж,И,К,Л)У | SMD-05 | ||||
| К1278ЕР1(Б,В,Г,Д,Ж,И,К,Л)П | TO-220 | IRU1117, APL1117, CS5203-1, CS5205-1 |
“Low drop” cтабилизатор положительного регулируемого напряжения. | ||
| К1278ЕР1(Ж,И,К,Л)Т3 | 4116.4-6 | Входное напряжение: 7В (Ж)/12В (И,К,Л), ток нагрузки 0.8/1.0/3.0/5.0А | |||
| К1278ЕР1(Ж,И,К,Л)У | SMD-05 | ||||
| 3. Микросхемы для управления двигателями, реле. | |||||
| K1128КТ3Р | PowerDIP (12+4) | L293D | Четырёхканальный полумостовой коммутатор тока. Us=4.5-36В, Is=1A. | ||
| K1128КТ4Р | PowerDIP (12+4) | L293D | Четырёхканальный полумостовой коммутатор тока с внутренними ограничительными диодами на выходах. Us=4.5-36В, Is=0.6A. | ||
| 4. Микросхемы операционных усилителей, компараторов, детекторов, кодовых ключей | |||||
| К1230ДП(46,87)П, К1230ДП(46,87)1П |
TO-92 | MC34064 | 46 – детектор понижения напряжения в системах с 5-ти вольтовым питанием; формирует сигнал Reset при понижении напряжения питания до 4.5-4.7В. | ||
| К1230ДП(46,87)Т, К1230ДП(46,87)1Т |
SOT-89 | 87 – детектор понижения напряжения в системах с 9-ти вольтовым питанием; формирует сигнал Reset при понижении напряжения питания до 8.5-8.9В. | |||
| К1230ДП(46,87)Т1, К1230ДП(46,87)1Т1 |
SO-8 | ||||
| К1233КТ2П | TO-92 | без аналога | Микросхема электронного кодового ключа для систем контроля и управления доступом контактного типа (268 435 456 комбинаций кода). | ||
| К1233КТ2Н | бескорпусная | ||||
| К1407УД2Р | DIP-8 | LM4250 | Программируемый малошумящий операционный усилитель. Ucc плюс-минус 6…12 В, Ку>50000, Uсм3МГц, Кос,сф>70дБ. | ||
| К1407УД2Т | SO-8 | ||||
| К1460УД2Р | DIP-8 | TCA0372 | Сдвоенный мощный операционный усилитель для управления индуктивными нагрузками. Io=1А, dUo/dt=1.3В/мкс, f=1.1МГц. | ||
| К1460УД2Р1 | PowerDIP (12+4) | ||||
| К1464CA1(Р,БР) | DIP-8 | LM393 | Сдвоенный микромощный компаратор напряжения. Ucc=2В…36В, +1В…+18В. | ||
| К1464CA1(Т,БР) | SO-8 | ||||
| К1464CA1Р1 | Д8К-2Н | ||||
| К1464CA1Т1 | Н02.8-2В | ||||
| К1464СА2БР | DIP-14 | LM339 | Счетверенный микромощный компаратор напряжения. Ucc=2В…36В, +1.0В…+18В. |
||
| К1464СА2Т | 401.14-5М | ||||
| К1464УД1(Б)Р | DIP-8 | LM358 | Сдвоенный микромощный операционный усилитель. Ucc=3В…32В, +1.5В…+16В. | ||
| К1464УД1(Б)Т | SO-8 | ||||
| К1464УД1Р1 | Д8К-2Н | ||||
| К1464УД1Т1 | Н02.8-2В | ||||
| К1464УД2(Б)Р | DIP-14 | LM324M | Счетверенный микромощный операционный усилитель. Ucc=3В…32В, +1.5В…+16В. | ||
| К1464УД2Т | 401.14-5М | ||||
| 5. Микросхемы для ключей, коммутаторов тока и напряжения | |||||
| K1109КН12(А,Б,В,Г) | DIP-18 | DI510-80 | 8-ми разрядный коммутатор анодного напряжения для управления газоразрядными индикаторами. Uсс=90B, Us=90B, Is=30mA. | ||
| K1109КН15Р | DIP-16 | без аналога | 6-ти разрядный драйвер сеток вакуумных индикаторов. Последовательный регистр, регистр-защелка. Uсс=5B, Us=минус 100B, Is=0.5mA. | ||
| K1109КН16Р | DIP-18 | DI512BR | 8-ми разрядный коммутатор анодного напряжения для управления газоразрядными индикаторами. Uсс=110B, Us=110B, Is=30mA. | ||
| К1109КТ8 | 4118.24-1Н | без аналога | Четырехканальный биполярный коммутатор тока с логикой на входе. | ||
| К1109КТ11 | 4118.24-1Н | без аналога | Четырехканальный биполярный коммутатор тока с логикой на входе. | ||
| К1109КТ14У | Н06.24-1В | без аналога | Двухканальный биполярный коммутатор тока. | ||
| К1109КТ21РБС | DIP-16 | ULN2002 | Набор мощных составных ключей индуктивных нагрузок. U=50 B, I=500 мА | ||
| К1109КТ21ТБС | 402.16-34 | ||||
| К1109КТ22РБС | DIP-16 | ULN2003 | Набор мощных составных ключей индуктивных нагрузок. U=50 B, I=500 мА | ||
| К1109КТ22ТБС | 402.16-34 | ||||
| К1109КТ23РБС | DIP-16 | ULN2004 | Набор мощных составных ключей индуктивных нагрузок. U=50 B, I=500 мА | ||
| К1109КТ23ТБС | 402.16-34 | ||||
| К1109КТ25РБС | DIP-16 | ULN2001 | Набор мощных составных ключей индуктивных нагрузок. U=50 B, I=500 мА | ||
| К1109КТ25ТБС | 402.16-34 | ||||
| К1224ПН1Р | DIP-8 | SP4424 | DC-AC преобразователь низкого входного напряжения питания (2-5В) в высокое (60-200В)для подсветки ЖКИ. | ||
| К1224ПН1Т | SO-8 | ||||
| К1224ПН3Р | DIP-8 | SP4412A | DC-AC преобразователь низкого входного напряжения питания (2-5В) в высокое (60-200В)для подсветки ЖКИ. | ||
| К1224ПН3Т | SO-8 | ||||
| К1224ПН4Р | DIP-8 | SP4422A | DC-AC преобразователь низкого входного напряжения питания (2-5В) в высокое (до 200В)для подсветки ЖКИ. | ||
| К1224ПН4Т | SO-8 | ||||
| К1224ПН5Р | DIP-8 | SP4425Q | DC-AC преобразователь низкого входного напряжения питания (2-5В) в высокое (до 200В) для подсветки ЖКИ. | ||
| К1224ПН5Т | SO-8 | ||||
| 6. Микросхемы для автоэлектроники. | |||||
| K1055ХП2Р | DIP-16 | L497 | Контроллер электронного коммутатора для бесконтактных систем зажигания с датчиком Холла. | ||
| К1055ХП2Т | SO-16 | ||||
| K1156ЕН1П | TO-220-5 | LM2925 | “Low drop” стабилизатор положительного напряжения на 5В со схемой “Сброс”. Uвх.max=26В , Uвых.=5В, Iн=0.5А, Uпдмин.=0.6В. | ||
| K1156ЕН5ВП | TO-220-5 | без аналога | Регулируемый ЛСН с низким Uпдмин. Uвх.max=26В , Uвых.=1.5-15В, Iн=0.5А, Uпдмин.=0.6В. | ||
| K1156ЕН5ДП | TO-220-5 | ||||
| K1156ЕН5ДУ | SLCC5/5 | ||||
| KР1158ЕН(3-15)01(А,Б) | TO-251 | L4945, LM2930, LM2931 | “Low drop” стабилизатор положительного фиксированного напряжения. Uвых.=3-15В с дискретностью 0.1В | ||
| КФ1158ЕН(3-15)01(А,Б) | TO-252 | ||||
| KР1158ЕН(3-15)(В,Г) | TO-220 | ||||
| KФ1158ЕН(3-15)(В,Г) | TO-263 | ||||
| K1158ЕН(3-15)(Ж,И)П | TO-220 | ||||
| K1158ЕН(3-15)(Ж,И)Т3 | 4116.4-6 | ||||
| K1158ЕН(3-15)(Ж,И)У | SMD-05 | ||||
| К1230ДП46П, К1230ДП461П |
TO-92 | MC34064 | Детектор понижения напряжения в системах с 5-ти вольтовым питанием; формирует сигнал Reset при понижении напряжения питания до 4.5-4.7В. | ||
| К1230ДП46Т, К1230ДП461Т |
SOT-89 | ||||
| К1230ДП46Т1, К1230ДП461Т1 |
SO-8 | ||||
| КТ863/БС(1) | TO-220 | без аналога | Кремниевый n-p-n транзистор. Imax.=12A, Imax.имп.=15А, UкеR>160В(R=1кОм), h31>200 с малым изменением в диапазоне рабочих температур -60њС…+130 њС |
||
| TO-263 | |||||
| 7. Микросхемы для телефонии. | |||||
| K1059КН2Р | DIP-18 | без аналога | ИС электронного коммутатора телефонной линии для систем охранной сигнализации (электронный аналог реле РПС-34). | ||
| K1059КП1Р | DIP-16 | без аналога | Автоматический телефонный комутатор. | ||
| 8. Транзисторные и диодные сборки, силовые модули | |||||
| КД2117(А,Б,В)-5 | бескорпусная | без аналога | Кремниевый диод с малым временем обратного восстановления. Постоянный прямой ток через диод до 100А, обратное напряжение до 1200В. | ||
| КД636(А,Б,В,Г,Д,Е)С | TO-220 | MBR20200CT | Пара диодов Шоттки с общим катодом: Imax.=2*15A, Imax.имп.=2*30А, Uпр.=1.2В при I=15A, tвост. | ||
| TO-263 | |||||
| КД637(А,Б,В,Г,Д,Е)С | TO-218 | без аналога | Пара диодов Шоттки с общим катодом: Imax.=2*25A, Imax.имп.=2*50А, Uпр.=1.4В при I=25A, tвост. | ||
| КД638(АО,А,Б,В,Г,Д,Е)С | TO-220 | без аналога | Пара диодов Шоттки с общим катодом: Imax.=2*5A, Imax.имп.=2*8А, Uпр.=1.0В при I=5A, tвост. | ||
| TO-263 | |||||
| КМ435(А,Б,В)С | 64х106х35 мм | без аналога | Силовые модули на базе IGDT и FRD. Uкэmax=1200В, Iкmax=200А(А), 300А(Б) и 400А(В). | ||
| КТ222(А,Б,В)С | DIP-8 | DI4044 | Согласованная пара транзисторов. Ubr=60В, I=10мкА…1мА, Kсогл_h31e=0.8…1.0, Rсогл_Ube=3мВ. | ||
| SO-8 | |||||
| КТ863/БС(1) | TO-220 | без аналога | Кремниевый n-p-n транзистор. Imax.=12A, Imax.имп.=15А, UкеR>160В(R=1кОм), h31>200 с малым изменением в диапазоне рабочих температур -60њС…+130њС | ||
| TO-263 | |||||
| М10 | SOT-227 (38х25х14 мм) | без аналога | Модули тиристорные, диодные, диодно-тиристорные и мосты. Uобр=400-1200В, It(f)sm=300-1600А. |
||
| МД(1,2,6,Ч2) | SOT-227 (25х38х20.7 мм) | ||||
| МТ(1,6,Д3) | |||||
| МПТ16 | 88х45х28 мм | ||||
Samsung подтвердила планы строительства завода по выпуску чипов в Техасе за $17 млрд — «Interfax» — последние новости Азербайджана, Грузии и Армении
Южнокорейская Samsung Electronics Co. подтвердила появившуюся ранее в СМИ информацию о том, что компания планирует инвестировать порядка $17 млрд в строительство предприятия по выпуску микросхем в городе Тейлор (штат Техас).
Это будут крупнейшие инвестиции Samsung в США, заявил производитель.
Завод будет выпускать высокотехнологичные чипы для смартфонов и других устройств, а также приложения для сферы искусственного интеллекта.
Строительство объекта стартует в следующем году. Samsung рассчитывает начать выпуск микросхем на этом заводе во втором полугодии 2024 года.
“Увеличив производственные мощности, мы сможем лучше служить потребностям наших клиентов и вносить свой вклад в стабильность глобальной цепочки поставок полупроводниковых компонентов”, – заявил главный исполнительный директор Samsung Electronics Device Solutions Ким Ки-нам.
Быстрое внедрение цифровых решений компаниями по всему миру в период пандемии COVID-19 создало дефицит полупроводниковых компонентов, который ударил по многим отраслям – от производства смартфонов и бытовой техники до автомобильной промышленности. Ранее Samsung сообщила, что планирует инвестиции более чем на $205 млрд в течение трех лет, отдавая приоритет развитию сегмента производства микросхем.
Акции Samsung Electronics подешевели на 0,7% по итогам торгов в Сеуле в среду. С начала этого года их стоимость снизилась на 7,7%.
Вся информация, размещенная на данном веб-сайте, предназначена только для персонального использования и подлежит дальнейшему распространению в какой-либо форме только с разрешения агентства «Интерфакс-Азербайджан».
Баку, Ичеришехер, улица В.Мустафазаде 6/10, 3-й этаж Бизнес-центра “Сахибтадж”
Телефоны: (99412) 497-35-07, 497-42-93 ● Факс: (99412) 497-42-94
E-mail: [email protected]
Различия в подкорково-корковых взаимодействиях, выявленных на моделях коннектома и микросхемы при аутизме
Калужна-Чаплинска, Й., Журавич, Э. и Ювик-Пруска, J. Сосредоточьтесь на социальном аспекте аутизма. J. Autism Dev. Disord. 48 , 1861–1867 (2018).
PubMed Статья PubMed Central Google ученый
Ганц, М. Л. Распределение дополнительных социальных издержек аутизма на протяжении жизни. Arch. Педиатр. Adolesc. Med. 161 , 343–349 (2007).
PubMed Статья PubMed Central Google ученый
Christensen, D. L. et al. Распространенность и характеристики расстройства аутистического спектра среди детей в возрасте 8 лет – Сеть мониторинга аутизма и нарушений развития, 11 сайтов, США, 2012 г. MMWR Surveill. Сумм. 65 , 1–23 (2018).
PubMed PubMed Central Статья Google ученый
Ли, Э., Ли, Дж. И Ким, Э. Дисбаланс возбуждения / торможения в моделях расстройств аутистического спектра на животных. Biol. Психиатрия 81 , 838–847 (2017).
PubMed Статья PubMed Central Google ученый
Нельсон, С. Б. и Валах, В. Возбуждающий / тормозной баланс и гомеостаз контура при расстройствах аутистического спектра. Нейрон 87 , 684–698 (2015).
CAS PubMed PubMed Central Статья Google ученый
Hong, S.-J. и другие. Атипичная функциональная иерархия коннектомов при аутизме. Nat. Commun. 10 , 1022 (2019).
ADS PubMed PubMed Central Статья CAS Google ученый
Наир, А., Трейбер, Дж. М., Шукла, Д. К., Ши, П. и Мюллер, Р. А. Нарушение таламокортикальной связи при расстройстве аутистического спектра: исследование функциональной и анатомической связи. Мозг 136 , 1942–1955 (2013).
PubMed PubMed Central Статья Google ученый
Нуньес, А.С., Питфилд, Н., Вакорин, В. и Дусбург, С.М. Идиосинкратическая организация корковых сетей при расстройствах аутистического спектра. Neuroimage 190 , 182–190 (2019).
PubMed Статья Google ученый
Сохал, В. С., Рубенштейн, Дж. Л. Р.Баланс возбуждения-торможения как основа для исследования механизмов психоневрологических расстройств. Мол. Психиатрия 24 , 1248–1257 (2019).
PubMed PubMed Central Статья Google ученый
Tesink, C. M. J. Y. et al. Нейронные корреляты прагматического понимания речи при расстройствах аутистического спектра. Мозг 132 , 1941–1952 (2009).
CAS PubMed Статья PubMed Central Google ученый
Вудворд, Н. Д., Хиральдо-Чика, М., Роджерс, Б. и Касио, К. Дж. Таламокортикальная дисфункция при расстройстве аутистического спектра: анализ обмена данными визуализации мозга при аутизме. Biol. Психиатрия Cogn. Neurosci. Нейровизуализация 2 , 76–84 (2017).
PubMed PubMed Central Google ученый
Pereira, A. M. et al. Различия в структуре коры и функциональной связи МРТ при высокофункциональном аутизме. Фронт. Neurol . 9 , 539 (2018).
PubMed PubMed Central Статья Google ученый
Jou, R.J. et al. Визуализация тензора диффузии при расстройствах аутистического спектра: предварительное свидетельство аномальной нейронной связи. Aust. Н. З. Дж. Психиатрия 45 , 153–162 (2011).
PubMed Статья PubMed Central Google ученый
Ли Ю., Пак Б., Джеймс О., Ким С.-Г. И Парк, Х. Расстройство аутистического спектра, связанное с функциональными изменениями связности в языковой сети у детей, подростков и взрослых. Фронт. Гм. Neurosci. 11 , 418 (2017).
PubMed PubMed Central Статья Google ученый
Ракич П. Спецификация областей коры головного мозга. Science 241 , 170–176 (1988).
ADS CAS PubMed Статья PubMed Central Google ученый
Робертсон К. Э. и Барон-Коэн С. Сенсорное восприятие при аутизме. Nat. Rev. Neurosci. 18 , 671–684 (2017).
CAS PubMed Статья PubMed Central Google ученый
Belmonte, M. K. et al. Аутизм как нарушение обработки нейронной информации: направления исследований и цели терапии. Мол. Психиатрия 9 , 646–663 (2004).
CAS PubMed Статья PubMed Central Google ученый
Wang, P. et al. Инверсия крупномасштабной модели схемы показывает корковую иерархию в динамическом состоянии покоя человеческого мозга. Sci. Adv. 5 , eaat7854 (2019).
ADS PubMed PubMed Central Статья Google ученый
Янг, Д. Ю. Дж., Бим, Д., Пелфри, К. А., Абдуллахи, С. и Джоу, Р. Дж. Морфологические маркеры коры у детей с аутизмом: исследование с помощью структурной магнитно-резонансной томографии толщины, площади, объема и гирификации. Мол. Аутизм 7 , 1–14 (2016).
Артикул Google ученый
Лау, В. К. В., Люнг, М. К. и Лау, Б. В. М. Аномалии состояния покоя при расстройствах аутистического спектра: метаанализ. Sci. Отчетность 9 , 3892 (2019).
ADS PubMed PubMed Central Статья CAS Google ученый
Williams, D. L. et al. Различия функций мозга при обработке речи у детей и взрослых с аутизмом. Autism Res. 6 , 288–302 (2013).
PubMed PubMed Central Статья Google ученый
Pagani, M. et al. Удаление гена риска аутизма shank3 нарушает префронтальную связь. J. Neurosci. 39 , 5299–5310 (2019).
CAS PubMed PubMed Central Статья Google ученый
Liska, A. et al. Гомозиготная потеря гена риска аутизма cntnap2 приводит к снижению локальной и дальнодействующей префронтальной функциональной связи. Cereb. Cortex 28 , 1141–1153 (2018).
PubMed Статья PubMed Central Google ученый
Bertero, A. et al. Связанная с аутизмом микроделеция 16p11.2 нарушает префронтальную функциональную связь у мыши и человека. Мозг 141 , 2055–2065 (2018).
PubMed Статья PubMed Central Google ученый
Антуан, М. В., Лангберг, Т., Шнепель, П.И Фельдман Д. Э. Повышенное отношение возбуждения к подавлению стабилизирует возбудимость синапсов и контуров в четырех моделях мышей с аутизмом. Neuron 101 , 648–661.e4 (2019).
CAS PubMed PubMed Central Статья Google ученый
Caeyenberghs, K. et al. Нейронная подпись нарушения координации развития в структурном коннектоме, независимом от коморбидного аутизма. Dev. Sci. 19 , 599–612 (2016).
PubMed Статья PubMed Central Google ученый
Catani, M. et al. Фронтальные сети у взрослых с расстройством аутистического спектра. Мозг 139 , 616–630 (2016).
PubMed PubMed Central Статья Google ученый
D’Albis, M. A. et al. Локальная структурная взаимосвязь связана с социальным познанием при расстройстве аутистического спектра. Мозг 141 , 3472–3481 (2018).
PubMed Статья PubMed Central Google ученый
Rudie, J. D. et al. Измененная функциональная и структурная организация сети мозга при аутизме. NeuroImage Clin. 2 , 79–94 (2013).
Артикул Google ученый
Tournier, J. D. et al. MRtrix3: быстрая, гибкая и открытая программная среда для обработки и визуализации медицинских изображений. Neuroimage 202 , 116137 (2019).
PubMed PubMed Central Статья Google ученый
Турнье, Дж. Д., Каламанте, Ф. и Коннелли, А. MRtrix: диффузионная трактография в пересекающихся областях волокон. Внутр. J. Imaging Syst. Technol. 22 , 53–66 (2012).
Артикул Google ученый
Смит Р.Э., Турнье, Дж. Д., Каламанте, Ф. и Коннелли, А. SIFT2: обеспечение плотной количественной оценки связности белого вещества мозга с использованием упрощенной трактографии. Neuroimage 119 , 338–351 (2015).
PubMed Статья Google ученый
Смит, Р. Э., Турнье, Дж. Д., Каламанте, Ф. и Коннелли, А. Анатомически ограниченная трактография: улучшенная диффузионная МРТ упрощает трактографию за счет эффективного использования анатомической информации. Neuroimage 62 , 1924–1938 (2012).
PubMed Статья Google ученый
Jeurissen, B., Tournier, J. D., Dhollander, T., Connelly, A. & Sijbers, J. Сферическая деконволюция с ограничениями нескольких тканей для улучшенного анализа данных МРТ диффузии с несколькими оболочками. Neuroimage 103 , 411–426 (2014).
PubMed Статья Google ученый
Christiaens, D. et al. Глобальная трактография данных визуализации, взвешенных по диффузии, с использованием модели нескольких тканей. Neuroimage 123 , 89–101 (2015).
PubMed Статья Google ученый
Хаак, К. В., Маркванд, А. Ф. и Бекманн, К. Ф. Коннекопическое картирование с помощью фМРТ в состоянии покоя. Neuroimage 170 , 83–94 (2018).
PubMed Статья Google ученый
Хаак, К. В. и Бекманн, К. Ф. Понимание организации мозга перед лицом функциональной неоднородности и функциональной множественности. Neuroimage 220 , 117061 (2020).
PubMed Статья Google ученый
Larivière, S. et al. Мультимасштабные структурно-функциональные градиенты в коннектоме новорожденных. Cereb. Cortex 30 , 47–58 (2020).
PubMed Статья Google ученый
Paquola, C. et al. Сдвиги миелоархитектуры характеризуют развитие корковых градиентов у подростков. elife 8 , 1-23 (2019).
Артикул Google ученый
Paquola, C. et al. Микроструктурные и функциональные градиенты все больше диссоциируют в трансмодальной коре. PLoS Biol. 17 , e3000284 (2019).
CAS PubMed PubMed Central Статья Google ученый
Vos de Wael, R. et al. BrainSpace: набор инструментов для анализа макромасштабных градиентов в наборах данных нейровизуализации и коннектомики. Commun. Биол. 3 , 103 (2020).
PubMed PubMed Central Статья Google ученый
Deco, G. et al. Функциональная связность в состоянии покоя возникает из медленных линейных флуктуаций структурной и динамической формы. J. Neurosci. 33 , 11239–11252 (2013).
CAS PubMed PubMed Central Статья Google ученый
Ширнер, М., Макинтош, А. Р., Джирса, В., Деко, Г. и Риттер, П. Вывод многомасштабных нейронных механизмов с моделированием сети мозга. elife 7 , 1–30 (2018).
Артикул Google ученый
Хансен, Э. К. А., Батталья, Д., Шпиглер, А., Деко, Г.И Джирса, В. К. Динамика функциональной связности: моделирование поведения переключения в состоянии покоя. Neuroimage 105 , 525–535 (2015).
PubMed Статья Google ученый
Брейкспир М. Динамические модели крупномасштабной мозговой деятельности. Nat. Neurosci. 20 , 340–352 (2017).
CAS PubMed Статья Google ученый
Васкес-Родригес, Б. и др. Градиенты структурно-функционального связывания в неокортексе. Proc. Natl Acad. Sci. США 116 , 21219–21227 (2019).
PubMed Статья CAS Google ученый
Суарес, Л. Э., Маркелло, Р. Д., Бетцель, Р. Ф. и Мисич, Б. Связывающая структура и функции в макромасштабных сетях мозга. Trends Cogn. Sci. 24 , 302–315 (2020).
PubMed Статья Google ученый
Miŝic, B. et al. Отношения структуры и функции сетевого уровня в неокортексе человека. Cereb. Cortex 26 , 3285–3296 (2016).
PubMed PubMed Central Статья Google ученый
Baum, G. L. et al. Развитие структурно-функциональной связи в сетях человеческого мозга в молодости. Proc. Natl Acad. Sci. США 117 , 771–778 (2020).
CAS PubMed Статья Google ученый
Маргулис, Д. С. и др. Размещение сети режима по умолчанию вдоль основного градиента макроуровневой кортикальной организации. Proc. Natl Acad. Sci. США 113 , 12574–12579 (2016).
CAS PubMed Статья PubMed Central Google ученый
Кулешов М.В. и др. Enrichr: обновление веб-сервера для комплексного анализа обогащения набора генов 2016 г. Nucleic Acids Res. 44 , W90 – W97 (2016).
CAS PubMed PubMed Central Статья Google ученый
Chen, E. Y. et al. Enrichr: интерактивный и совместный инструмент анализа пополнения списка генов HTML5. BMC Bioinformatics 14 , 128 (2013).
PubMed PubMed Central Статья Google ученый
Догерти, Дж. Д., Шмидт, Э. Ф., Накадзима, М.& Хайнц, Н. Аналитические подходы к данным профилирования РНК для идентификации генов, обогащенных конкретными клетками. Nucleic Acids Res. 38 , 4218–4230 (2010).
CAS PubMed PubMed Central Статья Google ученый
Цамардинос, И., Рахшани, А. и Лагани, В. Свойства оценки производительности протоколов на основе перекрестной проверки с одновременной оптимизацией гиперпараметров. Внутр. J. Artif. Intell. Инструменты 24 , 1540023 (2015).
Артикул Google ученый
Cawley, G.C. & Talbot, N.L.C. О чрезмерной подгонке при выборе модели и последующем предвзятом выборе при оценке эффективности. J. Mach. Учиться. Res. 11 , 2079–2107 (2010).
MathSciNet МАТЕМАТИКА Google ученый
Варма, С. и Саймон, Р. Смещение оценки ошибок при использовании перекрестной проверки для выбора модели. BMC Bioinformatics 7 , 91 (2006).
PubMed PubMed Central Статья CAS Google ученый
Parvandeh, S., Yeh, H. W., Paulus, M. P. & McKinney, B. A. Консенсус включает вложенную перекрестную проверку. Биоинформатика 36 , 3093–3098 (2020).
CAS PubMed PubMed Central Статья Google ученый
Di Martino, A. et al. Обмен данными визуализации мозга при аутизме: к крупномасштабной оценке внутренней архитектуры мозга при аутизме. Мол. Психиатрия 19 , 659–667 (2014).
PubMed Статья Google ученый
Di Martino, A. et al. Расширение исследований коннектома при аутизме с использованием обмена данными изображений мозга аутизма II. Sci. Данные 4 , 170010 (2017).
PubMed PubMed Central Статья Google ученый
Бецель, Р. Ф., Гриффа, А., Хагманн, П. и Мишиц, Б. Зависимые от расстояния пороги консенсуса для создания структурных сетей мозга, репрезентативных для группы. Netw. Neurosci. 3 , 475–496 (2019).
PubMed PubMed Central Статья Google ученый
Лангс, Г., Голланд, П. и Гош, С. С. Прогнозирование активации для отдельных лиц с помощью функции функционального подключения в состоянии покоя на основе слияния меток с несколькими атласами. в Международной конференции по медицинской обработке изображений и компьютерным вмешательствам 313–320 (2015).
Месулам, М. М. От ощущения к познанию. Мозг 121 , 1013–1052 (1998).
PubMed Статья PubMed Central Google ученый
Yeo, B. T. T. et al. Организация коры головного мозга человека оценивается по внутренней функциональной связности. J. Neurophysiol. 106 , 1125–1165 (2011).
PubMed Статья PubMed Central Google ученый
Bethlehem, R.A. I. et al. Дисперсия функциональных градиентов на протяжении взрослой жизни. Neuroimage 222 , 117299 (2020).
PubMed PubMed Central Статья Google ученый
Park, B. et al. Расширяющееся многообразие характеризует подростковые реконфигурации структурной организации коннектома. bioRxiv https://doi.org/10.1101/2020.06.22.165621 (2020).
Артикул PubMed PubMed Central Google ученый
Alexander-Bloch, A. F. et al. О тестировании пространственного соответствия между картами структуры и функций человеческого мозга. Neuroimage 178 , 540–551 (2018).
PubMed PubMed Central Статья Google ученый
Hawrylycz, M. J. et al. Анатомически исчерпывающий атлас транскриптома мозга взрослого человека. Природа 489 , 391–399 (2012).
ADS CAS PubMed PubMed Central Статья Google ученый
Gorgolewski, K. J. et al. NeuroVault.Org: веб-репозиторий для сбора и обмена статистическими картами человеческого мозга без обновления. Фронт. Нейроинформ. 9 , статья 8 (2015).
Артикул Google ученый
Арнаткевичуте, А., Фулчер, Б. Д. и Форнито, А. Практическое руководство по объединению экспрессии генов в мозге и данных нейровизуализации. Neuroimage 189 , 353–367 (2019).
PubMed Статья PubMed Central Google ученый
Gandal, M. J. et al. Нарушение регуляции на уровне изоформ в масштабе всего транскриптома при РАС, шизофрении и биполярном расстройстве. Наука 362 , 1265 (2018).
Артикул CAS Google ученый
Lake, B. B. et al. Подтипы и разнообразие нейронов, выявленные с помощью одноядерного секвенирования РНК головного мозга человека. Наука 352 , 1586–1590 (2016).
ADS CAS PubMed PubMed Central Статья Google ученый
Lake, B. B. et al. Интегративный одноклеточный анализ транскрипционных и эпигенетических состояний в мозге взрослого человека. Nat. Biotechnol. 36 , 70–80 (2018).
CAS PubMed PubMed Central Статья Google ученый
Lord, C. et al. Расписание диагностических наблюдений за аутизмом – универсальное: стандартная мера социальных и коммуникативных дефицитов, связанных со спектром аутизма. J. Autism Dev. Disord. 30 , 205–223 (2000).
CAS PubMed Статья PubMed Central Google ученый
Zou, H. & Hastie, T. Регуляризация и выбор переменных через Elastic Net. J. R. Stat. Soc. Сер. В 67 , 301–320 (2005).
MathSciNet МАТЕМАТИКА Статья Google ученый
Mueller, S. et al. Конвергентные результаты измененных функциональных и структурных соединений мозга у людей с высокофункциональным аутизмом: мультимодальное МРТ-исследование. PLoS ONE 8 , e67329 (2013).
ADS CAS PubMed PubMed Central Статья Google ученый
Berman, J. I. et al. Мультимодальная диффузионно-МРТ и МЭГ-оценка развития слуховой и языковой систем при расстройствах аутистического спектра. Фронт. Нейроанат. 10 , 30 (2016).
PubMed PubMed Central Статья Google ученый
Travers, B. et al. Визуализация тензора диффузии при расстройствах аутистического спектра: обзор. Autism Res . 5 , 289–313 (2012).
PubMed PubMed Central Статья Google ученый
Эрнандес, Л.М., Руди, Дж. Д., Грин, С. А., Букхаймер, С. и Дапретто, М. Нейронные сигнатуры расстройств аутистического спектра: понимание динамики сети мозга. Нейропсихофармакология 40 , 171–189 (2015).
PubMed Статья PubMed Central Google ученый
Li, H., Xue, Z., Ellmore, TM, Frye, RE & Wong, STC Сетевой анализ показывает более сильную локальную связь на основе диффузии и различные корреляции с навыками устной речи в мозгу детей с высокофункциональные расстройства аутистического спектра. Hum. Brain Mapp. 35 , 396–413 (2014).
PubMed Статья PubMed Central Google ученый
Li, S.J. et al. Изменения связности белого вещества у детей дошкольного возраста с расстройством аутистического спектра. Радиология 288 , 209–217 (2018).
PubMed Статья PubMed Central Google ученый
Roine, U. et al. Аномальное соединение коннектома у взрослых с высокофункциональным расстройством аутистического спектра. Мол. Аутизм 6 , 65 (2015).
PubMed PubMed Central Статья CAS Google ученый
Fang, H. et al. Структурные сети у детей с расстройством аутистического спектра с регрессией: исследование теории графов. Behav. Brain Res. 378 , 112262 (2020).
PubMed Статья PubMed Central Google ученый
Belmonte, M. K. et al. Аутизм и аномальное развитие мозговой связи. J. Neurosci. 24 , 9228–9231 (2004).
CAS PubMed PubMed Central Статья Google ученый
Assaf, M. et al. Аномальное функциональное соединение подсетей режима по умолчанию у пациентов с расстройством аутистического спектра. Neuroimage 53 , 247–256 (2010).
PubMed PubMed Central Статья Google ученый
Мизуно, А., Вильялобос, М. Э., Дэвис, М. М., Даль, Б. К. и Мюллер, Р. А. Частичное усиление таламокортикальной функциональной связи при аутизме. Мозг Res . 1104 , 160–174 (2006).
CAS PubMed Статья Google ученый
Di Martino, A. et al. Аберрантная функциональная связность полосатого тела у детей с аутизмом. Biol. Психиатрия 69 , 847–856 (2011).
PubMed Статья Google ученый
Nielsen, J. A. et al. Аномальная латерализация функциональной связи между языком и регионами режима по умолчанию при аутизме. Мол. Аутизм 5 , 8 (2014).
PubMed PubMed Central Статья Google ученый
Jouravlev, O. et al. Сниженная латерализация языка при аутизме и более широкий фенотип аутизма по оценке с помощью надежного индивидуального анализа. Autism Res. 13 , 1746–1761 (2020).
PubMed Статья Google ученый
Кляйнханс, Н. М., Мюллер, Р. А., Коэн, Д. Н., Курчесн, Э. Атипичная функциональная латерализация речи при расстройствах аутистического спектра. Мозг Res . 1221 , 115–125 (2008).
CAS PubMed PubMed Central Статья Google ученый
Postema, M. C. et al. Измененная структурная асимметрия мозга при расстройстве аутистического спектра в исследовании 54 наборов данных. Nat. Commun. 10 , 4958 (2019).
ADS PubMed PubMed Central Статья CAS Google ученый
Кардинале, Р. К., Ши, П., Фишман, И., Форд, Л. М. и Мюллер, Р. А. Распространенные сдвиги асимметрии функциональных сетей вправо при расстройстве аутистического спектра. JAMA Psychiatry 70 , 975–982 (2013).
PubMed PubMed Central Статья Google ученый
Эйлер, Л. Т., Пирс, К. и Курчесн, Э. Неспособность левой височной коры специализироваться на речи является ранним и фундаментальным свойством аутизма. Мозг 135 , 949–960 (2012).
PubMed PubMed Central Статья Google ученый
Floris, D. L. et al. Атипичная латерализация функциональной связности моторных цепей у детей с аутизмом связана с моторным дефицитом. Мол. Аутизм 7 , 35 (2016).
PubMed PubMed Central Статья Google ученый
Облак, А. Л., Розен, Д. Л., Кемпер, Т. Л., Бауман, М. Л. и Блатт, Г. Дж. Измененная циктоархитектура коры задней части поясной извилины, но нормальная плотность нейронов и интернейронов в задней части поясной коры и веретенообразной извилине при аутизме. Autism Res. 4 , 200–211 (2011).
PubMed PubMed Central Статья Google ученый
Симмс, М. Л., Кемпер, Т. Л., Тимби, К. М., Бауман, М.Л. и Блатт, Г. Дж. Передняя поясная кора при аутизме: неоднородность качественных и количественных цитоархитектонических характеристик предполагает возможные подгруппы. Acta Neuropathol. 118 , 673–684 (2009).
PubMed Статья PubMed Central Google ученый
Кемпер Т. Л. и Бауман М. Вклад невропатологических исследований в понимание аутизма. Neurol. Clin. 11 , 175–187 (1993).
CAS PubMed Статья PubMed Central Google ученый
Кемпер Т. Л. и Бауман М. Невропатология детского аутизма. J. Neuropathol. Exp. Neurol. 57 , 645–652 (1998).
CAS PubMed Статья PubMed Central Google ученый
Казанова, М.Ф., Buxhoeveden, D. P., Switala, A. E. & Roy, E. Плотность и архитектура нейронов (индекс серого уровня) в мозге аутичных пациентов. J. Child Neurol. 17 , 515–521 (2002).
PubMed Статья PubMed Central Google ученый
Казанова, М. Ф., Буксхеведен, Д. П., Свитала, А. Э. и Рой, Е. Миниколоночная патология при аутизме. Неврология 58 , 428–432 (2002).
PubMed Статья PubMed Central Google ученый
Авино, Т. А. и Хатслер, Дж. Дж. Аномальное формирование клеточного паттерна на границе серого и белого вещества коры при расстройствах аутистического спектра. Brain Res. 1360 , 138–146 (2010).
CAS PubMed Статья PubMed Central Google ученый
Зикопулос, Б.И Барбас, Х. Измененные нейронные связи в возбуждающих и тормозных корковых цепях при аутизме. Фронт. Гм. Neurosci. 7 , 609 (2013).
PubMed PubMed Central Статья Google ученый
Хатслер, Дж. Дж. И Чжан, Х. Повышенная плотность дендритных позвонков на корковых проекционных нейронах при расстройствах аутистического спектра. Мозг Res . 1309 , 83–94 (2010).
CAS PubMed Статья PubMed Central Google ученый
Gonçalves, J. et al. Проверка гипотезы дисбаланса возбуждения / торможения на мышиной модели расстройства аутистического спектра: нейроспектроскопия in vivo и молекулярные доказательства региональных фенотипов. Мол. Аутизм 8 , 47 (2017).
PubMed PubMed Central Статья CAS Google ученый
Gogolla, N. et al. Дефект общей цепи возбудительно-тормозного баланса в мышиных моделях аутизма. J. Neurodev. Disord. 1 , 172–181 (2009).
PubMed PubMed Central Статья Google ученый
Guilmatre, A., Huguet, G., Delorme, R. & Bourgeron, T. Возникающая роль генов SHANK в нервно-психических расстройствах. Dev. Neurobiol. 74 , 113–122 (2014).
CAS PubMed Статья PubMed Central Google ученый
Bourgeron, T. От генетической архитектуры до синаптической пластичности при расстройствах аутистического спектра. Nat. Rev. Neurosci. 16 , 551–563 (2015).
CAS PubMed Статья PubMed Central Google ученый
Shao, Y. et al. Точное отображение аутичного расстройства на хромосоме 15q11-q13 с использованием фенотипических подтипов. г. J. Hum. Genet. 72 , 539–548 (2003).
CAS PubMed PubMed Central Статья Google ученый
млн лет назад, D. Q. et al. Выявление значительной ассоциации и взаимодействия генов-генов субъединиц ГАМК-рецепторов при аутизме. г. J. Hum. Genet. 77 , 377–388 (2005).
CAS PubMed PubMed Central Статья Google ученый
Zheng, Z., Zhu, T., Qu, Y. & Mu, D. Уровни глутамата в крови при расстройствах аутистического спектра: систематический обзор и метаанализ. PLoS ONE 11 , e0158688 (2016).
PubMed PubMed Central Статья CAS Google ученый
Fatemi, SH, Folsom, TD, Kneeland, RE & Liesch, SB Повышение регуляции метаботропного рецептора глутамата 5 у детей с аутизмом связано с недостаточной экспрессией как ломкого протеина умственной отсталости X, так и рецептора бета 3 ГАМК у взрослых с аутизм. Анат. Рек. 294 , 1635–1645 (2011).
CAS Статья Google ученый
Макфадьен, Дж., Долан, Р. Дж. И Гарридо, М. I. Влияние подкорковых сокращений на неупорядоченную сенсорную и когнитивную обработку. Nat. Ред. Neurosci . https://doi.org/10.1038/s41583-020-0287-1 (2020).
Ван, X. Дж. Макроскопические градиенты синаптического возбуждения и торможения в неокортексе. Nat. Rev. Neurosci. 21 , 169–178 (2020).
CAS PubMed PubMed Central Статья Google ученый
Van Essen, D.C. et al. Проект WU-Minn Human Connectome: обзор. Neuroimage 80 , 62–79 (2013).
PubMed PubMed Central Статья Google ученый
Ханнант П., Кэссиди С., Тавассоли Т. и Манн Ф. Сенсомоторные трудности связаны с тяжестью состояний аутистического спектра. Фронт. Интегр. Neurosci. 10 , 28 (2016).
PubMed PubMed Central Статья Google ученый
Bertolero, M.A. et al. Сетевая архитектура человеческого мозга генетически закодирована модульной плейотропией. arXiv https://arxiv.org/abs/1905.07606 (2019).
Hawrylycz, M. et al. Канонические генетические признаки мозга взрослого человека. Nat. Neurosci. 18 , 1832–1844 (2015).
CAS PubMed PubMed Central Статья Google ученый
Альтманн, А.и другие. Анализ на системном уровне подчеркивает активацию микроглии как модифицирующий фактор при распространенных формах эпилепсии человека. bioRxiv https://doi.org/10.1101/470518 (2018).
van den Heuvel, M. P. et al. Эволюционные модификации связности человеческого мозга, связанные с шизофренией. Мозг 142 , 3991–4002 (2019).
PubMed PubMed Central Статья Google ученый
Форнито А., Арнаткявичюте А. и Фулчер Б. Д. Преодоление разрыва между коннектомом и транскриптомом. Trends Cogn. Sci. 23 , 34–50 (2019).
PubMed Статья PubMed Central Google ученый
Romero-Garcia, R., Warrier, V., Bullmore, E. T., Baron-Cohen, S. & Bethlehem, R.A. I. Синаптические и транскрипционно подавляемые гены связаны с различиями в толщине коры при аутизме. Мол. Психиатрия 24 , 1053–1064 (2019).
CAS PubMed Статья PubMed Central Google ученый
Burt, J. B. et al. Иерархия транскриптомной специализации коры головного мозга человека, зафиксированная топографией структурной нейровизуализации. Nat. Neurosci. 21 , 1251–1259 (2018).
CAS PubMed PubMed Central Статья Google ученый
Wang, Z. et al. Информированная сообществом коннектомика таламокортикальной системы при генерализованной эпилепсии. Неврология 93 , e1112 – e1122 (2019).
CAS PubMed PubMed Central Статья Google ученый
Хван, К., Бертолеро, М. А., Лю, В. Б. и Д’Эспозито, М. Таламус человека является интегративным центром функциональных сетей мозга. J. Neurosci. 37 , 5594–5607 (2017).
CAS PubMed PubMed Central Статья Google ученый
Colich, N. L. et al. Атипичная нейронная обработка иронических и искренних замечаний у детей и подростков с расстройствами аутистического спектра. Метафора Сим. 27 , 70–92 (2012).
PubMed PubMed Central Статья Google ученый
Varoquaux, G.Ошибка перекрестной проверки: малые размеры выборки приводят к большим ошибкам. Neuroimage 180 , 68–77 (2018).
PubMed Статья Google ученый
Poldrack, R.A. et al. Сканирование горизонта: к прозрачным и воспроизводимым исследованиям нейровизуализации. Nat. Rev. Neurosci. 18 , 115–126 (2017).
CAS PubMed PubMed Central Статья Google ученый
Ву, К. В., Чанг, Л. Дж., Линдквист, М. А. и Вейджер, Т. Д. Создание лучших биомаркеров: модели мозга в трансляционной нейровизуализации. Nat. Neurosci. 20 , 365–377 (2017).
CAS PubMed PubMed Central Статья Google ученый
Хонг, С. Дж., Валк, С. Л., Ди Мартино, А., Милхэм, М. П. и Бернхардт, Б. С. Многомерное нейроанатомическое подтипирование расстройства аутистического спектра. Cereb. Cortex 28 , 3578–3588 (2018).
PubMed Статья PubMed Central Google ученый
Lombardo, M. V. et al. Режим по умолчанию – гипоконнективность визуальной сети в подтипе аутизма с выраженными трудностями социального визуального взаимодействия. elife 8 , e47427 (2019).
CAS PubMed PubMed Central Статья Google ученый
Lombardo, M. V. et al. Крупномасштабные ассоциации между лейкоцитарным транскриптомом и ЖЕЛТЫМИ ответами на речь различаются в подтипах ранних языковых исходов аутизма. Nat. Neurosci. 21 , 1680–1688 (2018).
CAS PubMed PubMed Central Статья Google ученый
Ломбардо, М. В., Лай, М. К. и Барон-Коэн, С. Подходы с использованием больших данных для разложения неоднородности в спектре аутизма. Мол. Психиатрия 24 , 1435–1450 (2019).
PubMed PubMed Central Статья Google ученый
Lord, C., Rutter, M. & Le Couteur, A. Пересмотренное диагностическое интервью для аутизма: исправленная версия диагностического интервью для лиц, осуществляющих уход за людьми с возможными распространенными нарушениями развития. J. Autism Dev. Disord. 24 , 659–685 (1994).
CAS PubMed Статья PubMed Central Google ученый
Дейл, А. М., Фишл, Б. и Серено, М. I. Анализ кортикальной поверхности: I. Сегментация и реконструкция поверхности. Neuroimage 9 , 179–194 (1999).
CAS PubMed Статья PubMed Central Google ученый
Фишл Б., Лю А. и Дейл А. М. Автоматизированная хирургия многообразия: построение геометрически точных и топологически правильных моделей коры головного мозга человека. IEEE Trans. Med. Imaging 20 , 70–80 (2001).
CAS PubMed Статья PubMed Central Google ученый
Фишл Б., Серено М. И. и Дейл А. М. Анализ кортикальной поверхности: II. Инфляция, выравнивание и система координат на основе поверхности. Neuroimage 9 , 195–207 (1999).
CAS PubMed Статья PubMed Central Google ученый
Фишл, Б., Серено, М. И., Тутелл, Р. Б. Х. и Дейл, А. М. Межсубъектное усреднение с высоким разрешением и поверхностная система координат. Hum. Brain Mapp. 8 , 272–284 (1999).
CAS PubMed PubMed Central Статья Google ученый
Фишл, Б. FreeSurfer Neuroimage 62 , 774–781 (2012).
PubMed PubMed Central Статья Google ученый
Ségonne, F., Pacheco, J. & Fischl, B. Геометрически точная коррекция топологии кортикальных поверхностей с использованием неразделимых петель. IEEE Trans. Med. Imaging 26 , 518–529 (2007).
PubMed Статья Google ученый
Craddock, C. et al. На пути к автоматическому анализу коннектомов: настраиваемый конвейер для анализа коннектомов (C-PAC). Фронт. Нейроинформ. https: // doi.org / 10.3389 / conf.fninf.2013.09.00042 (2013).
Артикул Google ученый
Behzadi, Y., Restom, K., Liau, J. & Liu, T. T. Метод коррекции шума на основе компонентов (CompCor) для BOLD и перфузионной фМРТ. Neuroimage 37 , 90–101 (2007).
PubMed PubMed Central Статья Google ученый
Пауэр, Дж.D. et al. Методы обнаружения, характеристики и удаления артефактов движения в состоянии покоя фМРТ. Neuroimage 84 , 320–341 (2014).
PubMed Статья Google ученый
Пауэр, Дж. Д., Барнс, К. А., Снайдер, А. З., Шлаггар, Б. Л. и Петерсен, С. Е. Ложные, но систематические корреляции в функциональной связности МРТ-сетей возникают из-за движения субъекта. Neuroimage 59 , 2142–2154 (2012).
PubMed Статья PubMed Central Google ученый
Турнье, Ж.-Д., Каламанте, Ф. и Коннелли, А. Улучшенная трактография вероятностных линий тока за счет интеграции 2-го порядка по распределению ориентации волокон. в Proc. Международное общество магнитного резонанса в медицине (2010).
Schaefer, A. et al. Локально-глобальное расчленение коры головного мозга человека с помощью МРТ внутренней функциональной связности. Cereb. Cortex 28 , 3095–3114 (2018).
PubMed Статья PubMed Central Google ученый
Форнито А., Залески А. и Баллмор Э. Основы анализа сети мозга . (Академик Пресс, 2016).
Тененбаум, Дж. Б., де Сильва, В. и Лэнгфорд, Дж. С. Глобальная геометрическая структура для нелинейного уменьшения размерности. Наука 290 , 2319–2323 (2000).
ADS CAS PubMed Статья PubMed Central Google ученый
Фон Люксбург, У. Учебное пособие по спектральной кластеризации. Stat. Comput. 17 , 395–416 (2007).
MathSciNet Статья Google ученый
Park, B. et al. Распространение сигнала по градиентам коннектома и маршрутизация между концентраторами по-разному влияют на динамическую функцию человеческого мозга. Neuroimage 224 , 117429 (2021).
PubMed Статья PubMed Central Google ученый
Бенджамини Ю. и Хохберг Ю. Контроль уровня ложных открытий: практичный и эффективный подход к множественному тестированию. J. R. Stat. Soc. 57 , 289–300 (1995).
MathSciNet МАТЕМАТИКА Google ученый
Stoodley, C.J. et al. Изменение связности мозжечка при аутизме и опосредованное мозжечком спасение связанного с аутизмом поведения у мышей. Nat. Neurosci. 20 , 1744–1751 (2017).
CAS PubMed PubMed Central Статья Google ученый
Francis, S. M. et al. Гипосоединение изолированных сетей состояния покоя у подростков с расстройством аутистического спектра. Psychiatry Res. Нейровизуализация 283 , 104–112 (2019).
PubMed Статья PubMed Central Google ученый
Рамос, Т. К., Балардин, Дж. Б., Сато, Дж. Р. и Фуджита, А. Аномальная кортико-мозжечковая функциональная связь при расстройстве аутистического спектра. Фронт. Syst. Neurosci. 12 , 74 (2019).
PubMed PubMed Central Статья Google ученый
Cerliani, L.и другие. Повышенная функциональная связь между подкорковыми и корковыми сетями состояния покоя при расстройстве аутистического спектра. JAMA Psychiatry 72 , 767–777 (2015).
PubMed PubMed Central Статья Google ученый
Максимо, Дж. О. и Кана, Р. К. Аберрантная «глубокая взаимосвязь» при аутизме: исследование кортико-подкорковой функциональной взаимосвязи с помощью магнитно-резонансной томографии. Autism Res. 12 , 384–400 (2019).
PubMed Статья Google ученый
Патенауд Б., Смит С. М., Кеннеди Д. Н. и Дженкинсон М. Байесовская модель формы и внешнего вида для подкорковой сегментации мозга. Neuroimage 56 , 907–922 (2011).
PubMed PubMed Central Статья Google ученый
Дидрихсен, Дж., Балстерс, Дж. Х., Флавелл, Дж., Кассанс, Э. и Рамнани, Н. Вероятностный МР-атлас мозжечка человека. Neuroimage 46 , 39–46 (2009).
PubMed Статья Google ученый
Фристон, К. Дж., Харрисон, Л. и Пенни, В. Динамическое причинное моделирование. Neuroimage 19 , 1273–1302 (2003).
CAS PubMed Статья Google ученый
Хайнцле, Дж., Купманс, П. Дж., Ден Ауден, Х. Э. М., Раман, С. и Стефан, К. Э. Гемодинамическая модель для слоистых BOLD-сигналов. Neuroimage 125 , 556–570 (2016).
PubMed Статья Google ученый
Стефан К. Э., Вейскопф Н., Дрисдейл П. М., Робинсон П. А. и Фристон К. Дж. Сравнение гемодинамических моделей с DCM. Neuroimage 38 , 387–401 (2007).
PubMed PubMed Central Статья Google ученый
Wang, P. & Knösche, T.R. Реалистичная модель нейронной массы коры с ламинарно-специфическими связями и синаптической пластичностью – оценка со слуховым привыканием. PLoS ONE 8 , e77876 (2013).
ADS CAS PubMed PubMed Central Статья Google ученый
Гандал, М. Дж., Леппа, В., Вон, Х., Парикшак, Н. Н. и Гешвинд, Д. Х. Путь к точной психиатрии: перевод генетики в механизмы болезни. Nat. Neurosci. 19 , 1397–1407 (2016).
CAS PubMed Статья Google ученый
Steijger, T. et al. Оценка методов реконструкции транскриптов для RNA-seq. Nat. Методы 10 , 1177–1184 (2013).
CAS PubMed PubMed Central Статья Google ученый
Любовь, м.I., Hogenesch, J. B. & Irizarry, R. A. Моделирование смещения последовательности фрагментов РНК-seq снижает систематические ошибки в оценке количества транскриптов. Nat. Biotechnol. 34 , 1287–1291 (2016).
CAS PubMed PubMed Central Статья Google ученый
Статистический анализ свойств обработки информации пластинчатых моделей кортикальных микросхем | Кора головного мозга
Аннотация
Основной задачей вычислительной нейробиологии является понимание вычислительной функции пластин-специфических паттернов синаптических соединений в стереотипных кортикальных микросхемах.Предыдущая работа над этой проблемой была сосредоточена на гипотезах о конкретных вычислительных ролях отдельных слоев и связях между уровнями, и эти гипотезы были проверены путем моделирования абстрактных моделей нейронных сетей. Вместо этого мы подходим к этой проблеме, изучая динамическую систему, определяемую более реалистичными моделями кортикальных микросхем в целом, и исследуя влияние, которое ее ламинарная структура оказывает на передачу и слияние информации в этой динамической системе. Модели цепей, которые мы изучаем, состоят из нейронов Ходжкина-Хаксли с динамическими синапсами, что основано на подробных данных Томсона и др. (2002), Маркрама и др. (1998) и Гупты и др. (2000).Мы исследуем, в какой степени этот шаблон кортикальной микросхемы поддерживает накопление и слияние информации, содержащейся в общих импульсных входах, в слой 4 и слои 2/3 и насколько хорошо он делает эту информацию доступной для проекционных нейронов в слоях 2/3 и 5. продемонстрировать конкретные вычислительные преимущества такой модели кортикальной микросхемы, зависящей от пластинки, на основе данных путем сравнения ее производительности с различными типами управляющих моделей, которые имеют те же компоненты и одинаковую глобальную статистику нейронов и синаптических связей, но не имеют характерной для пластинки структуры реальных корковые микросхемы.Мы пришли к выводу, что компьютерное моделирование детальных пластинчатых моделей кортикальных микросхем обеспечивает новое понимание вычислительных последствий анатомических и физиологических данных.
Введение
Неокортекс состоит из нейронов в различных пластинках, которые образуют четко структурированные микросхемы. Несмотря на многочисленные различия в зависимости от возраста, области коры и вида, многие свойства этих микросхем стереотипны, что позволяет предположить, что неокортикальные микросхемы являются вариациями общего шаблона микросхем (White 1989; Douglas and others 1995; Mountcastle 1998; Nelson 2002; Silberberg и другие 2002; Дуглас и Мартин 2004; Калисман и другие 2005).Можно предположить, что такой шаблон микросхемы отличается определенными функциональными свойствами, которые позволяют ему обслуживать огромные вычислительные и когнитивные возможности мозга более эффективно, чем, например, случайным образом подключенная схема с тем же количеством нейронов и синапсов. . Потенциальная вычислительная функция ламинарной структуры схемы уже рассматривалась в многочисленных статьях (см., Например, Raizada and Grossberg 2003; Treves 2003; Douglas and Martin 2004; и ссылки в этих недавних публикациях).Treves (2003) и Raizada и Grossberg (2003) исследовали конкретные гипотезы относительно вычислительной роли структуры, специфичной для ламины, и подтвердили эти гипотезы с помощью компьютерного моделирования довольно абстрактных моделей нейронных цепей.
Результаты Treves (2003) показывают, что в этой более абстрактной настройке структура ламинарной схемы дает небольшое преимущество в отношении разделения двух типов информации: в каком горизонтальном положении вводится схема (информация «где») и о конкретном шаблоне, который был введен туда («какая» информация).Мы дополняем этот анализ более внимательным изучением временной динамики информации в одном горизонтальном местоположении, точнее в одном столбце диаметром около 100 мкм. Мы обнаружили, что с этой точки зрения вычислительное преимущество ламинарных схем значительно больше: около 30% (в зависимости от конкретного типа задачи обработки информации), а не только 10%, как это наблюдалось в Treves (2003).
Стереотипная кортикальная микросхема – это очень повторяющаяся цепь, которая включает в себя множество наложенных друг на друга петель положительной и отрицательной обратной связи (Дуглас и Мартин 2004).Большинство методов, которые были разработаны в технических науках для проектирования и анализа таких рекуррентных схем, сосредоточены на системном поведении рекуррентной схемы в целом, поскольку оказалось невозможным понять возникающую динамику нелинейных рекуррентных схем только на основе на основе технических характеристик их компонентов. Эта системная точка зрения на стереотипные корковые микросхемы впервые была подчеркнута Дугласом и другими (1995) и привела к их определению абстрактной модели «канонической микросхемы».Дуглас и другие (1995) продемонстрировали, что подход этой системы обеспечивает новый способ понимания роли торможения в корковых микросхемах, в частности того, каким образом относительно небольшие изменения тормозной обратной связи могут вызывать большие изменения в усилении система. Однако в этих ранних моделях не учитывалась временная динамика нейрональной и синаптической активности. Также в то время было доступно гораздо меньше данных о стереотипных паттернах связи.Кроме того, ранее не предпринималось попыток проанализировать с помощью строгих статистических методов возникающие возможности обработки информации полученных в результате подробных моделей микросхем. Цель данной статьи – восполнить этот пробел.
Мы исследуем возможности обработки информации подробными моделями микросхем на основе данных Томсона и др. (2002) о вероятностях связей, специфичных для пластин, и силе связи между возбуждающими и тормозящими нейронами в слоях 2/3, 4, 5 и на данных. от Markram и др. (1998) и Gupta и др. (2000) относительно стереотипных динамических свойств (таких как парное подавление пульса и парное облегчение пульса) синаптических связей между возбуждающими и тормозящими кортикальными нейронами.Наш анализ основан на предположении, что стереотипные корковые микросхемы обладают некоторыми «универсальными» вычислительными возможностями и могут выполнять совершенно разные вычисления в разных областях коры. Следовательно, он концентрируется на общей способности обработки информации удерживать и объединять информацию, содержащуюся во входных цепочках импульсов Пуассона из 2 разных источников (моделирование таламического или кортикального прямого ввода в слой 4 и латерального или нисходящего ввода в слои 2/3). . Кроме того, мы исследовали способность таких схемных моделей выполнять линейные и нелинейные вычисления для изменяющихся во времени скоростей срабатывания этих двух афферентных входных потоков.Чтобы избежать – обязательно весьма предвзятых – предположений о нейронном кодировании результатов таких вычислений, мы проанализировали информацию, которая доступна о результатах таких вычислений для обычных «нейронных пользователей», то есть пирамидных нейронов в слои 2/3 (которые обычно проецируются в верхние области коры) и пирамидные нейроны в слое 5 (которые обычно не только проецируются в нижние области коры или подкорковые структуры, но также проецируются, например, из V1 обратно в неспецифический таламус, т.е.е., к интраламинарным и срединным ядрам, которые не получают прямого первичного сенсорного ввода, и через это реле к более высоким кортикальным областям, см. Callaway 2004).
В отличие от модели Маасса и других (2002) (обсуждение см. В Destexhe and Marder 2004), мы не использовали простую линейную регрессию для оценки информации, доступной для таких считывающих нейронов, выход которых моделируется взвешенным сумма постсинаптических потенциалов (PSP) (с постоянной времени экспоненциального затухания 15 мс) в ответ на спайки от пресинаптических нейронов.Скорее, мы добавили сюда ограничение, согласно которому вклад возбуждающего (тормозящего) пресинаптического нейрона должен иметь положительный (отрицательный) вес в такой взвешенной сумме. Кроме того, мы приняли во внимание, что считывающий нейрон в слоях 2/3 или 5 получает синаптические входы только от довольно небольшого подмножества нейронов в микросхеме согласно данным Thomson и др. (2002) (что подразумевает, что в цепь из 560 нейронов, нейрон в слоях 2/3 имеет в среднем 84 пресинаптических нейрона, а нейрон в слое 5 имеет в среднем 109 пресинаптических нейронов, см. рис.1). Но, как и в более ранней модели, мы не изменили параметры синапсов в схеме для конкретных вычислительных задач, а только веса синаптических связей с такими символьными считывающими нейронами на уровнях 2/3 и 5 (которые не были смоделированы как часть схему в том смысле, что они не проецировались обратно в схему). (Это упрощение было сделано в этой статье из прагматических соображений, поскольку первые результаты по случаю с обратной связью [Maass and others 2005] предполагают, что это требует отдельного анализа).
Рисунок 1
Шаблон кортикальной микросхемы. Цифры у стрелок обозначают силу соединения (средняя амплитуда PSP, измеренная в соме в мВ) и вероятности соединения (в скобках) согласно Thomson и др. (2002) для соединений между корковыми нейронами в 3 разных слоях, каждый из которых состоит из возбуждающего (E ) и ингибирующая (I) популяция с предполагаемым максимальным горизонтальным расстоянием до 100 мкм. Большинство данных взяты из коры головного мозга крысы, за исключением соединений в слое 4 (курсив), которые взяты из cat.(Соединения от L2 / 3-I до L5-E описаны в Thomson and others [2002], но обсуждаются только качественно. Следовательно, запись для соединений от L2 / 3-I до L5-E [отмечена знаком вопроса ] является лишь экстраполяцией. То же самое относится и к соединениям от L4-I к L2 / 3-I. Никаких данных об амплитудах тормозных PSP от L5-I к L5-I в Thomson и др. [2002] не приводится, следовательно, соответствующая запись – это всего лишь предположение.) Проценты во входных потоках обозначают вероятности соединения для входных нейронов, используемых в наших симуляциях.Кроме того, каждый нейрон получает фоновый шум, отражающий синаптические входы от большого количества более удаленных нейронов (см. Методы).
Рисунок 1
Шаблон кортикальной микросхемы. Цифры у стрелок обозначают силу соединения (средняя амплитуда PSP, измеренная в соме в мВ) и вероятности соединения (в скобках) согласно Thomson и др. (2002) для соединений между корковыми нейронами в 3 разных слоях, каждый из которых состоит из возбуждающего (E ) и ингибирующая (I) популяция с предполагаемым максимальным горизонтальным расстоянием до 100 мкм.Большинство данных взяты из коры головного мозга крысы, за исключением соединений в слое 4 (курсив), которые взяты из cat. (Соединения от L2 / 3-I до L5-E описаны в Thomson and others [2002], но обсуждаются только качественно. Следовательно, запись для соединений от L2 / 3-I до L5-E [отмечена знаком вопроса ] является лишь экстраполяцией. То же самое относится и к соединениям от L4-I к L2 / 3-I. Никаких данных об амплитудах тормозных PSP от L5-I к L5-I у Томсона и др. [2002] не приводится, следовательно, соответствующая запись – это всего лишь предположение.) Проценты на входных потоках обозначают вероятности подключения для входных нейронов, используемых в нашем моделировании. Кроме того, каждый нейрон получает фоновый шум, отражающий синаптические входы от большого количества более удаленных нейронов (см. Методы).
Методы
Наиболее полный в настоящее время набор данных о вероятностях соединения и эффективности синаптических связей между 6 конкретными популяциями нейронов в корковых микросхемах (возбуждающие и тормозящие нейроны в слоях 2/3, 4 и 5) был собран в Thomson et al. (2002). ).Для сбора этих данных были сделаны внутриклеточные записи с помощью острых электродов от 998 пар идентифицированных нейронов. В общей сложности 679 парных записей были сделаны из соматосенсорных, моторных и зрительных зон взрослых крыс и 319 из зрительных зон взрослых кошек. Отбор проб производился случайным образом с латеральным разбросом 50–100 мкм (А. М. Томсон, личное сообщение). Для тех пар, где данные как от крысы, так и от кошки приведены в Thomson and others (2002), мы взяли данные от крысы (см. Рис.1). Только для пар нейронов в слое 4 данные по крысе у Thomson и др. (2002) не приводятся; следовательно, соответствующие данные на Рисунке 1 взяты из cat. (Некоторые пары наблюдались редко, а соответствующие записи страдают от небольшого размера выборки [подробности см. В Thomson and others 2002]. Также очень маленькие нейроны у крысы могли быть пропущены [AM Thomson, личное сообщение]. Кроме того, возможно, что в некоторых кортикальных микросхемах существуют связи между парами нейронов, о которых не сообщалось в Thomson и др. [2002] [в случае соединений с тормозными нейронами в слоях 2/3, см., например, e.g., Dantzker and Callaway 2000]).
Известно, что краткосрочная динамика корковых синапсов (то есть их специфическая смесь парной депрессии пульса и парного облегчения пульса) зависит от типа пресинаптического и постсинаптического нейрона (см., Например, Markram and others 1998; Gupta et al. 2000; Томсон 2003). Мы смоделировали эту краткосрочную синаптическую динамику в соответствии с моделью, предложенной Маркрамом и другими (1998), с синаптическими параметрами U, D и F . Модель предсказывает амплитуду A k PSP для k -го спайка в цепочке спайков с интервалами между спайками Δ 1 , Δ 2 ,…, Δ k −1 через рекурсивные уравнения (1) со скрытыми динамическими переменными u ∈ [0,1] и R ∈ [0,1], чьи начальные значения для 1-го пика равны u 1 = U и R 1 = 1 (см. Maass and Markram [2002] для обоснования этой версии уравнений, которая исправляет небольшую ошибку в Markram и др. [1998]).Детерминированная модель синапсов разработана для моделирования средней суммы постсинаптических ответов, возникающих в результате согласованного действия нескольких стохастических сайтов синаптического высвобождения. Результаты показывают, что включение краткосрочной синаптической пластичности оказывает значительное влияние на способность моделей цепей к обработке информации. (Долговременная синаптическая пластичность в моделируемой цепи не была включена в это исследование по прагматическим причинам из-за дополнительных сложных проблем, но будет рассмотрена в последующих исследованиях).Параметры U, D и F были выбраны в нашей компьютерной модели из распределений Гаусса, которые отражают данные, представленные в Markram и др. (1998) и Gupta и др. (2000) для каждого типа соединения (обратите внимание, что параметр U согласно Markram и др. [1998] в значительной степени определяется вероятностью начального высвобождения задействованных сайтов синаптического высвобождения). В зависимости от того, был ли вход возбуждающим (E) или тормозным (I), были выбраны средние значения этих трех параметров U, D, F (с D, F в секундах), чтобы иметь средние значения, которые были сообщается в этих статьях (см. Таблицу 1).Стандартное отклонение (SD) каждого параметра было выбрано равным 50% от его среднего (с заменой отрицательных значений значениями, выбранными из равномерного распределения между нулем и двукратным средним).
Таблица 1Синаптические параметры, которые масштабируют краткосрочную динамику
| От / до | E | I |
| E | 0,5, 1,1, 0,05 | 0,05, 0,12516, 1,21691 |
| Я | 0.25, 0,7, 0,02 | 0,32, 0,144, 0,06 |
| От / до | E | I |
| E | 0,5, 1,116 0,05 0,05 | |
| I | 0,25, 0,7, 0,02 | 0,32, 0,144, 0,06 |
Синаптические параметры, которые масштабируют краткосрочную динамику
| От / до | E | |
| E | 0.5, 1,1, 0,05 | 0,05, 0,125, 1,2 |
| I | 0,25, 0,7, 0,02 | 0,32, 0,144, 0,06 |
| От / до | E | |
| E | 0,5, 1,1, 0,05 | 0,05, 0,125, 1,2 |
| I | 0,25, 0,7, 0,02 | 0,32, 0,144, 0,06 |
В состав микросхем входили модели из 3 слоев, с 30%, 20% и 50% нейронов, назначенных на уровни 2/3, слой 4 и слой 5, соответственно.Каждый слой состоял из популяции возбуждающих нейронов и популяции тормозных нейронов в соотношении 4: 1. Синаптические связи между нейронами в любой паре из 6 полученных популяций были случайно сгенерированы в соответствии с эмпирическими данными из таблицы 1 и рисунка 1. Большинство схем, которые были смоделированы, состояли из 560 нейронов. Среднее количество пресинаптических нейронов нейрона в такой цепи было тогда 76, что в целом давало в среднем 42 594 синапса в цепи.
В качестве моделей возбуждающих и тормозных нейронов мы выбрали основанные на проводимости одноэлементные модели нейронов Ходжкина-Хаксли с пассивными и активными свойствами, смоделированные согласно Destexhe и др. (2001) и Destexhe and Pare (1999).В соответствии с экспериментальными данными о пирамидных нейронах неокортекса и гиппокампа (Стюарт и Сакманн, 1994; Маги, Джонстон, 1995; Хоффман и др., 1997; Маги и др., 1998), активные токи содержат зависящий от напряжения ток Na + (Трауб и Майлз 1991) и выпрямитель тока с задержкой K + (Трауб и Майлз, 1991). Для возбуждающих нейронов в модель был включен неинактивирующий ток K + (Mainen и др., 1995), ответственный за адаптацию частоты спайков.Пиковая плотность проводимости для тока Na + и тока выпрямителя с задержкой K + была выбрана равной 500 и 100 пСм / мкм 2 , соответственно, а пиковая плотность проводимости для неинактивирующего тока K + было выбрано 5 пСм / мкм 2 . Площадь мембраны нейрона была установлена равной 34 636 мкм 2 , как у Destexhe и др. (2001). Для каждого моделирования начальные условия каждого нейрона, то есть напряжение на мембране в момент времени t = 0, выбирались случайным образом (равномерное распределение) из интервала [-70 мВ, -60 мВ].
Кортикальный нейрон получает синаптические входы не только от непосредственно соседних нейронов (которые были явно смоделированы в нашей компьютерной модели), но также и меньшие фоновые входные токи от большого количества более отдаленных нейронов. Фактически, внутриклеточные записи у бодрствующих животных предполагают, что нейроны неокортекса подвергаются интенсивной бомбардировке фоновыми синаптическими входами, вызывая деполяризацию мембранного потенциала и более низкое входное сопротивление, обычно называемое «состоянием высокой проводимости» (для обзора, см. Destexhe and others 2003).Это было отражено в нашей компьютерной модели фоновыми входными токами, которые вводились в каждый нейрон (в дополнение к явно смоделированным синаптическим входам от афферентных соединений и нейронов в цепи). Проводимость этих фоновых токов моделировалась согласно Destexhe и др. (2001) как случайный процесс с одной переменной, аналогичный процессу Орнштейна-Уленбека со средним значением g e = 0,012 мкс и g i = 0,057 мкСм, дисперсия σ e = 0.003 мкСм и σ i = 0,0066 мкСм, а постоянные времени τ e = 2,7 мСм и τ i = 10,5 мСм, где индексы e / i относятся к входной проводимости возбуждающего и тормозящего фона соответственно. Согласно Destexhe и др. (2001), эта модель захватывает спектральные и амплитудные характеристики входных проводимостей детальной биофизической модели неокортикальной пирамидной клетки, которая была сопоставлена с внутриклеточными записями в теменной коре кошек in vivo. Кроме того, соотношение средних вкладов возбуждающей и тормозной фоновых проводимостей было выбрано равным 5 в соответствии с экспериментальными исследованиями во время сенсорных реакций (Borg-Graham and others 1998; Hirsch and others 1998; Anderson and others 2000).
Максимальные проводимости синапсов были выбраны из гауссова распределения со стандартным отклонением 70% от его среднего (с заменой отрицательных значений значениями, выбранными из равномерного распределения между нулем и двукратным средним). Средние максимальные проводимости синапсов были выбраны для воспроизведения средней амплитуды PSP, представленной на рисунке 1, при мембранном потенциале покоя (при наличии синаптической фоновой активности).
Два афферентных входных потока, каждый из которых состоит из 4 или 40 цепочек пиков (т.е., 4 или 40 входных каналов), были введены в схему. Каждый из каналов 1-го входного потока (представляющий таламический или прямой кортикальный вход) вводился не только в слой 4, то есть в 50% его тормозных нейронов и 80% его возбуждающих нейронов, но также и в 20% возбуждающих нейронов. нейроны в слоях 2/3 и 10% возбуждающих нейронов в слое 5 (все выбираются случайным образом). (Это входное распределение качественно отражает данные, приведенные в главе III White [1989], что «таламокортикальные афференты к синапсу 4 уровня не только с непирамидными нейронами 4 уровня, но и с широким спектром как пирамидных, так и непирамидных типов нейронов, чьи клеточные тела встречаются повсюду. слои 2–6.Таким образом, среднее количество входов, сходящихся к возбуждающему нейрону в слое 4, составляет 3,2 или 32. (Компьютерное моделирование предполагает, что меньшие вероятности подключения от внешних входных нейронов могут быть выбраны, если амплитуды результирующих PSP увеличиваются соответствующим образом. Для 40 входных каналов мы выполнили моделирование с более низким входным соединением для входного потока 1, сохранив при этом постоянное произведение амплитуды PSP и вероятности соединения. инвариантен к этим изменениям, даже если вероятности подключения для внешних входных нейронов уменьшены до 1/5 от ранее заданных значений.Это примерно находится в диапазоне, предполагаемом экспериментальными измерениями вариабельности возбуждающих постсинаптических потенциалов (ВПСП) в простых клетках зрительной коры головного мозга кошек с различными уровнями стимуляции латерального коленчатого ядра (LGN) (Ferster 1987) и экспериментами по взаимной корреляции между моносинаптическими связанные клетки LGN и зрительной коры головного мозга кошки (Tanaka 1983), что предполагает, что по крайней мере 10 клеток LGN обеспечивают вход в каждую простую клетку. Средняя проводимость входных синапсов была выбрана для генерации PSP со средней амплитудой 1.9 мВ при мембранном потенциале покоя (при наличии синаптической фоновой активности). Это значение соответствует нижней границе оценки воздействия коленчатого тела на один нейрон в слое 4 взрослых кошек, данной Chung and Ferster (1998). Это было умножено в наших симуляциях на параметр масштабирования S I1 , который отражает биологически нереалистичное количество входных нейронов в этих симуляциях (см. Обсуждение ниже). Каждый из каналов 2-го афферентного входного потока вводился в 20% возбуждающих нейронов в слоях 2/3 (также со средней амплитудой 1.9 мВ, умноженное на другой параметр масштабирования S I2 ).
Таблица 2Снижение производительности цепей управления по сравнению со схемами, основанными на данных
| Задачи / схемы | Аморфные | В малых масштабах | С контролем по степени | С контролем по степени без специфичности входа или выхода | Случайно краткосрочная синаптическая динамика | Статические синапсы | ||||
| Память | 32.6 | 41,6 | 12,0 | 35,8 | 48,3 | 65,7 | ||||
| Нелинейный | 36,9 | 11,3 | −2,3 | 4,6 | 40,18 5,3 | -0,6 | 5,6 | 14,1 | 6,9 | |
| Все | 25,0 | 15,4 | 1,6 | 12,0 | 30.4 | 30,6 |
| Задачи / схемы | Аморфный | Маленький мир | Градусный | Градусный без специфичности входа или выхода | Случайная краткосрочная динамика | Случайная краткосрочная динамика Статические синапсы | |||||
| Память | 32,6 | 41,6 | 12,0 | 35,8 | 48,3 | 65,7 | |||||
| Нелинейный | 36.9 | 11,3 | -2,3 | 4,6 | 40,1 | 38,7 | |||||
| Прочие | 12,2 | 5,3 | -0,6 | 5,6 | 1691 | ,0 | 15,4 | 1,6 | 12,0 | 30,4 | 30,6 |
Снижение производительности схем управления по сравнению со схемами, основанными на данных
| Задачи / схемы | Аморфные Управляемая по степени | Регулируемая по степени без специфичности ввода или вывода | Случайная краткосрочная синаптическая динамика | Статические синапсы | ||||||
| Память | 32.6 | 41,6 | 12,0 | 35,8 | 48,3 | 65,7 | ||||
| Нелинейный | 36,9 | 11,3 | −2,3 | 4,6 | 40,18 5,3 | -0,6 | 5,6 | 14,1 | 6,9 | |
| Все | 25,0 | 15,4 | 1,6 | 12,0 | 30.4 | 30,6 |
| Задачи / схемы | Аморфный | Маленький мир | Градусный | Градусный без специфичности входа или выхода | Случайная краткосрочная динамика | Случайная краткосрочная динамика Статические синапсы | |||||
| Память | 32,6 | 41,6 | 12,0 | 35,8 | 48,3 | 65,7 | |||||
| Нелинейный | 36.9 | 11,3 | -2,3 | 4,6 | 40,1 | 38,7 | |||||
| Прочие | 12,2 | 5,3 | -0,6 | 5,6 | 1691 | ,0 | 15,4 | 1,6 | 12,0 | 30,4 | 30,6 |
Всего остается 3 параметра, значения которых необходимо выбрать, чтобы получить функциональные компьютерные модели кортикальных микросхем.Эти параметры S RW , S I1 , S I2 масштабируют (в виде мультипликативных множителей) амплитуды PSP для всех синаптических связей в цепи (рекуррентные веса), амплитуды PSP из входного потока 1 и амплитуды EPSP из входного потока 2. Они должны быть выбраны таким образом, чтобы они учитывали разницу в масштабе между нашими смоделированными микросхемами и биологическими кортикальными микросхемами.Значения этих трех параметров не могут быть считаны из ранее упомянутых данных, и нужно подозревать, что адекватные значения зависят также от вида, от конкретной корковой микросхемы in vivo, которую человек хочет смоделировать, от текущего состояния различных гомеостатических процессов. , от текущего поведенческого состояния (включая внимание) организма и от интенсивности текущего афферентного воздействия.
Параметр S I1 был выбран таким образом, чтобы афферентный входной поток 1 (состоящий из 40 последовательностей импульсов Пуассона с частотой 20 Гц) вызывал (без входного потока 2 и без повторяющихся соединений, т.е.е., S RW установлен на 0) средняя частота возбуждения 15 Гц в слое 4. Параметр S I2 был выбран аналогичным образом, так что афферентный входной поток 2 (сгенерированный как входной поток 1) вызвал средняя частота стрельбы 10 Гц в слоях 2/3. В любом случае был активирован только один из 2 входных потоков. С помощью этой процедуры мы получили S I1 = 14 и S I2 = 33. Для моделирования с входными потоками, состоящими из 4 последовательностей импульсов Пуассона, мы умножили эти значения на 10.Входные синапсы были выбраны статическими, то есть синаптические параметры были установлены на U = 1, D = 0 и F = 0, а их максимальные проводимости были выбраны из гауссовского распределения с SD. 70% от его среднего (с заменой отрицательных значений значениями, выбранными из равномерного распределения между нулем и двукратным средним).
Параметр S RW учитывает среднюю мощность синаптических входов в нейрон от других нейронов в цепи (кроме глобально смоделированного фонового синаптического входа, см. Выше), и, следовательно, разницу в размере цепи между наши смоделированные модели микросхем и настоящая кортикальная микросхема.Оказалось, что значение 60000 / (количество нейронов в моделируемой цепи) для S RW произведено в слое 5 моделируемой схемы для стандартных значений S I1 и S I2 представляет собой реалистичную низкую, но значительную активность возбуждения 8,5 Гц (см. Рис. 2), поэтому мы использовали это значение в качестве стандартного значения для S RW . Это значение увеличивает среднее число 76 пресинаптических нейронов в цепи из 560 нейронов до 107 раз, давая, таким образом, в среднем 8132 пресинаптических нейрона.Это число согласуется с оценками общего числа синапсов на нейроне, данными Бинцеггером и другими (2004), которые варьируются от 2981 до 13 075 для разных типов клеток зрительной коры головного мозга кошек. Некоторый дополнительный синаптический ввод моделировался фоновым синаптическим вводом (см. Выше).
Рисунок 2
( A ) Два входных потока, каждый из которых состоит из 40 последовательностей пиков Пуассона (вход в слои 2/3 начинается здесь на 100 мс позже). ( B ) Спайк-растр модели кортикальной микросхемы на основе данных (состоящей из 560 нейронов) для 2 входных потоков, показанных в ( A ).Вертикальное измерение масштабируется в соответствии с количеством нейронов в каждом слое. Шипы тормозных нейронов показаны серым цветом. (Показатели активизации населения в разных слоях в некоторой степени, но не полностью коррелированы. Максимальный коэффициент корреляции между темпами увольнения населения в слоях 2/3 и 4, слоях 2/3 и 5, а также в слое 4 и 5 [для t > 150 мс, размер интервала 1 мс и произвольная задержка] составляет 0,62, 0,65 и 0,56 соответственно.) ( C ) Распределение скоростей стрельбы в ( B ) (после начала ввода в слои 2 / 3 и слой 4), показывая автоматически возникающее экспоненциальное распределение скоростей возгорания в слое 5.Средняя частота стрельбы: 8,5 Гц. ( D ) Увеличение начального временного сегмента, показывающее распространение возбуждения от слоя 4 к поверхностным и глубоким слоям, что качественно совпадает с данными Армстронга-Джеймса и других (1992).
Рисунок 2
( A ) Два входных потока, каждый из которых состоит из 40 последовательностей пиков Пуассона (вход в слои 2/3 начинается здесь на 100 мс позже). ( B ) Спайк-растр модели кортикальной микросхемы на основе данных (состоящей из 560 нейронов) для 2 входных потоков, показанных в ( A ).Вертикальное измерение масштабируется в соответствии с количеством нейронов в каждом слое. Шипы тормозных нейронов показаны серым цветом. (Показатели активизации населения в разных слоях в некоторой степени, но не полностью коррелированы. Максимальный коэффициент корреляции между темпами увольнения населения в слоях 2/3 и 4, слоях 2/3 и 5, а также в слое 4 и 5 [для t > 150 мс, размер интервала 1 мс и произвольная задержка] составляет 0,62, 0,65 и 0,56 соответственно.) ( C ) Распределение скоростей стрельбы в ( B ) (после начала ввода в слои 2 / 3 и слой 4), показывая автоматически возникающее экспоненциальное распределение скоростей возгорания в слое 5.Средняя частота стрельбы: 8,5 Гц. ( D ) Увеличение начального временного сегмента, показывающее распространение возбуждения от слоя 4 к поверхностным и глубоким слоям, что качественно совпадает с данными Армстронга-Джеймса и других (1992).
Эти стандартные значения параметров S RW , S I1 , S I2 использовались во всех наших компьютерных экспериментах, за исключением результатов, представленных на рисунках 10 и 11, где мы анализировали влияние этих параметров на отчетные результаты.Там мы смоделировали модели схем со случайно выбранными значениями из независимых равномерных распределений в интервале [0,1 × стандартное значение, 3,1 × стандартное значение] для всех 3 параметров.
Наше компьютерное моделирование проверило, сколько информации о каждом предыдущем временном сегменте (длиной 30 мс) каждого из двух входных потоков было доступно гипотетическому проекционному нейрону в слоях 2/3 и гипотетическому проекционному нейрону в слое 5.. возбуждающие и тормозные пресинаптические нейроны для такого гипотетического считывающего нейрона выбирались случайным образом таким же образом, как и для любого другого возбуждающего нейрона в этом слое (т.е.е., согласно рис. 1), но никаких синаптических соединений от считывающего нейрона обратно в схему не было. Это составило в среднем 84 пресинаптических нейрона для считывающего нейрона в слоях 2/3 и 109 пресинаптических нейронов для считывающего нейрона в слое 5. Вес синаптических связей от этих пресинаптических нейронов был оптимизирован для конкретных задач. В отличие от моделирования, описанного в Maass и др. (2002), результирующее количество входов в такой считывающий нейрон было намного меньше, чем размер схемы.Сами нейроны проекции или считывания были смоделированы как линейные нейроны, то есть их выход представлял собой взвешенную сумму спайков, отфильтрованных нижними частотами (экспоненциальный спад с постоянной времени 15 мс, моделирование постоянных времени синаптических рецепторов и мембраны считывающего устройства). нейрон). Были приняты меры для того, чтобы веса возбуждающих (тормозных) пресинаптических нейронов не могли стать отрицательными (положительными). Для этого мы использовали линейный метод наименьших квадратов с ограничениями неотрицательности (Lawson and Hanson 1974), чтобы оптимизировать веса для конкретной задачи.Это контрастирует с линейной регрессией, которая использовалась в Maass и др. (2002). Для каждого обучающего или тестового примера, который состоял из входа и целевого значения для считывающего нейрона, мы выполняли моделирование модели микросхемы. Каждый вход для считывающего нейрона был сгенерирован путем сбора отфильтрованной нижними частотами версии пресинаптических шиповых последовательностей для считывающего нейрона в момент времени t = 450 мс. Каждое соответствующее целевое значение было рассчитано для различных задач, описанных ниже (см. Результаты).Чтобы правильно применить линейный метод наименьших квадратов с ограничениями неотрицательности, последовательности импульсов тормозных (возбуждающих) нейронов были свернуты с отрицательными (положительными) экспоненциальными ядрами, а соответствующие веса считывания были умножены на -1 (+1) после обучения. Для задач классификации нейрон с линейным считыванием был обучен выводить метки классов, то есть 0 или 1, тогда как классификация была получена путем установления порога вывода на 0,5 (аналогично порогу срабатывания реального коркового нейрона).Этот алгоритм дает весовой вектор < w i ,… w n > с w i ≥ 0, если i -й пресинаптический нейрон считывания является возбуждающим. и w i ≤ 0, если i -й пресинаптический нейрон является тормозящим. В рамках этих (линейных) ограничений эта ограниченная форма линейной регрессии минимизирует ошибку считывания в обучающих примерах.(В MATLAB можно выполнить этот алгоритм оптимизации с помощью команды LSQNONNEG.) Это обычно приводило к присвоению веса 0 (соответствующего молчащему синапсу в биологической цепи) примерно 2/3 этих синапсов. Следовательно, типичный считывающий нейрон имел менее 40 ненулевых весов и, следовательно, гораздо меньшую способность извлекать информацию по сравнению с моделью, рассмотренной в Maass и др. (2002).
Для каждого компьютерного моделирования было создано не менее 10 схем.Для экспериментов, показанных на рисунках 5 и 8, мы использовали 20 схем. Мы также генерировали новые шаблоны всплесков каждый раз, когда рисовалась новая схема, чтобы избежать случайной зависимости от свойств конкретных шаблонов всплесков. Для обучения считывающих нейронов мы выполнили 1500 симуляций со случайно выбранными пуассоновскими входами в течение 450 мс, а для тестирования были использованы 300 симуляций с новыми случайно сгенерированными входами. Планки погрешностей на рисунках обозначают стандартные ошибки. Все результаты производительности в этой статье (за исключением некоторых результатов диагностики, представленных на рис.8, см. Легенду) предназначены для тестовых входов, которые не использовались для обучения считыванию, и недавно сгенерированных случайных начальных условий и фонового шума для всех нейронов в цепи.
Все моделирование проводилось с помощью программного обеспечения CSIM (Natschläger и др., 2003) в сочетании с MATLAB.
Результаты
Инжекция 2 входных потоков, состоящих из цугов пуассоновских спайков, в слой 4 и слои 2/3 модели микросхемы вызвала отклик (см.рис.2), чье последовательное начало в разных слоях качественно совпадает с данными о корковых микросхемах in vivo (Armstrong-James и др., 1992). Кроме того, частота воспламенения в слое 5 автоматически приобретает биологически реалистичное экспоненциальное распределение (см., Например, v. Vreeswijk and Sompolinsky 1996, 1998; Amit and Brunel 1997; Baddeley and others 1997). (Распределение скоростей воспламенения в слоях 2/3 и 4 отражает типичное распределение скоростей последовательностей импульсов Пуассона, которые были вызваны пуассоновским воздействием на эти слои.Рисунок 3 дает представление о довольно большой вариабельности активности зажигания внутри схемы для одних и тех же схем ввода пиков, что является результатом дрожания на входе пиков (верхний ряд) и внутреннего шума (нижний ряд) из-за введение случайно изменяющихся фоновых входных токов ко всем нейронам для моделирования условий in vivo (см. Методы). (Кроме того, для всех рассматриваемых впоследствии вычислительных задач независимо выбранные паттерны спайков были ранее введены в качестве афферентных входов, вызывая довольно большую вариацию начальных состояний динамических синапсов.Таким образом, смоделированные схемы качественно отражают обычно наблюдаемую большую вариабельность нервных реакций in vivo от испытания к испытанию на повторение одного и того же стимула.
Рисунок 3
Влияние временного дрожания входных пиков (гауссово распределение со средним значением 0 и SD 1 мс) и фонового шума нейронов в цепи на отклик схемы (см. Методы). Строки в середине и вверху показывают растры пиков, полученные в результате 2 испытаний с идентичным фоновым шумом и с образцами пиков на входе, которые были идентичны, за исключением их временного дрожания.В нижнем ряду показано, насколько растр пиков для испытания с новым фоновым шумом и идентичными шаблонами пиков на входе отличается от такового для испытания, показанного в средней строке. Это иллюстрирует, что смоделированные цепи (которые были подвержены обоим источникам шума) качественно отражают обычно наблюдаемую большую вариабельность нервных реакций in vivo от испытания к испытанию на повторение одного и того же стимула.
Рисунок 3
Влияние временного дрожания входных всплесков (гауссово распределение со средним значением 0 и SD 1 мс) и фонового шума нейронов в цепи на отклик схемы (см. Методы).Строки в середине и вверху показывают растры пиков, полученные в результате 2 испытаний с идентичным фоновым шумом и с образцами пиков на входе, которые были идентичны, за исключением их временного дрожания. В нижнем ряду показано, насколько растр пиков для испытания с новым фоновым шумом и идентичными шаблонами пиков на входе отличается от такового для испытания, показанного в средней строке. Это иллюстрирует, что смоделированные цепи (которые были подвержены обоим источникам шума) качественно отражают обычно наблюдаемую большую вариабельность нервных реакций in vivo от испытания к испытанию на повторение одного и того же стимула.
Мы протестировали эти модели микросхем на множестве общих задач обработки информации, которые, вероятно, будут связаны с реальными вычислительными задачами нейронных микросхем в коре головного мозга: для задач обработки информации с паттернами спайков мы случайным образом сгенерировали шаблоны паттернов спайков, состоящие из 30 мс сегментов 40 последовательностей спайков Пуассона на частоте 20 Гц (см. рис. 4). Точнее, последовательности пиков каждого из двух входных потоков имели длину 450 мс и состояли из 15 последовательных временных сегментов длиной 30 мс.Для каждого сегмента случайным образом были сгенерированы 2 шаблона паттернов шипов. Для фактического ввода один из 2 шаблонов каждого временного сегмента был выбран случайным образом (с равной вероятностью) и его вариация с шумом, где каждый всплеск перемещался на величину, взятую из гауссовского распределения со средним значением 0 и SD 1 мс (см. панель с правой стороны рис. 4) была введена в схему. Такой временной джиттер на входе пиков вызывает значительные изменения в отклике схемы (см. Рис. 3), и для считывающих нейронов классифицировать паттерны спайков является нетривиальной задачей, несмотря на эту довольно большую вариабельность отклика схемы от испытаний к испытаниям. .Мы также протестировали ретроактивную классификацию предыдущих паттернов спайков, которые были введены за 30 мс до этого и, следовательно, были «перезаписаны» независимо выбранными последующими паттернами спайков. Кроме того, было исследовано нелинейное вычисление исключающего ИЛИ (исключающее ИЛИ) на образцах пиков в 2 параллельных входных потоках, чтобы проверить способность схемы извлекать и комбинировать информацию из обоих входных потоков нелинейным образом. Задача состоит в том, чтобы вычислить XOR (XOR выводит 1, если ровно один из его 2 входных битов имеет значение 1, он выводит 0, если входные биты равны 00 или 11) двух битов, которые представляют метки двух шаблонов, из которых были сгенерированы самые последние образцы всплесков в 2 входных потоках (например,g., его целевой выход равен 1 для обоих временных сегментов для входа, показанного в правой части рис. 4). Обратите внимание, что это вычисление включает в себя операцию нелинейной привязки к шаблонам пиков, потому что оно должно давать низкое выходное значение тогда и только тогда, когда либо шумные версии шаблонов пиков с меткой 1 появляются как во входных потоках 1 и 2, либо если шумная версия шаблоны всплесков с меткой 0 появились как во входном потоке 1, так и во входном потоке 2.
классификация шаблонов всплесков в любом из двух афферентных входных потоков (требующая инвариантности к шуму и всплеску входного сигнала из другого входного потока)
временная интеграция информации, содержащейся в таких шаблонах всплесков
Слияние информации из шаблонов всплесков в обоих входных потоках нелинейным образом (связанное с задачами «привязки»)
вычислений в реальном времени для скоростей активации из обоих входных потоков.
Рисунок 4
Распределения входных данных для классификации паттернов спайков и задач XOR. Задача состоит в том, чтобы вычислить XOR двух битов, которые представляют метки 2 шаблонов, из которых были сгенерированы самые последние шаблоны всплесков в 2 входных потоках, то есть классифицировать как 1, если их метки шаблонов различны, и 0 в противном случае. . Последовательности пиков каждого из 2 входных потоков имели длину 450 мс и состояли из 15 временных сегментов длиной 30 мс. Для каждого сегмента случайным образом были сгенерированы 2 шаблона (40 последовательностей пуассоновских спайков при 20 Гц).Фактические последовательности пиков каждого входа длиной 450 мс, используемые для обучения или тестирования, были сгенерированы путем выбора для каждого сегмента одного из 2 ранее выбранных связанных шаблонов, а затем создания версии с дрожанием путем перемещения каждого пика на величину, взятую из распределения Гаусса с среднее значение 0 и стандартное отклонение 1 мс (образец показан на панели справа).
Рисунок 4
Входные распределения для классификации шаблонов пиков и задач XOR. Задача состоит в том, чтобы вычислить XOR двух битов, которые представляют метки 2 шаблонов, из которых были сгенерированы самые последние шаблоны всплесков в 2 входных потоках, то есть классифицировать как 1, если их метки шаблонов различны, и 0 в противном случае. .Последовательности пиков каждого из 2 входных потоков имели длину 450 мс и состояли из 15 временных сегментов длиной 30 мс. Для каждого сегмента случайным образом были сгенерированы 2 шаблона (40 последовательностей пуассоновских спайков при 20 Гц). Фактические последовательности пиков каждого входа длиной 450 мс, используемые для обучения или тестирования, были сгенерированы путем выбора для каждого сегмента одного из 2 ранее выбранных связанных шаблонов, а затем создания версии с дрожанием путем перемещения каждого пика на величину, взятую из распределения Гаусса с среднее значение 0 и стандартное отклонение 1 мс (образец показан на панели справа).
Кроме того, мы проанализировали линейные и нелинейные вычисления для изменяющихся во времени скоростей срабатывания 2 входных потоков. Последовательности пиков каждого из 2 входных потоков имели длину 450 мс и состояли из 15 последовательных временных сегментов длиной 30 мс. Для каждого входного потока и каждого временного сегмента были сгенерированы 4 последовательности пиков Пуассона со случайно выбранной частотой от 15 до 25 Гц. Фактические скорости воспламенения, использованные для вычислений этих входных скоростей воспламенения, были рассчитаны на основе этих последовательностей импульсов со скользящим окном шириной 15 мс.Для этих тестов мы использовали входные потоки, состоящие всего из 4 последовательностей импульсов, поскольку производительность как схем на основе данных, так и схем управления была довольно низкой, если входные скорости были представлены 40 последовательностями импульсов.
Новые вычислительные свойства моделей микросхем на основе данных показаны на рисунке 5 (серые столбцы). Производительность обученных считываний для тестовых входных данных (которые генерируются из того же распределения, что и обучающие примеры, но не показаны во время обучения) была измерена для всех задач двоичной классификации с помощью коэффициента каппа, который находится в диапазоне [-1, 1], и принимает значение ≥0 тогда и только тогда, когда результирующая классификация тестовых примеров дает меньше ошибок, чем случайное предположение.(Коэффициент каппа измеряет процент соответствия между двумя классами, ожидаемыми сверх случайности, и определяется как ( P o – P c ) / (1 – P c ), где P o – наблюдаемое совпадение, а P c – случайное совпадение. Таким образом, для классификации на 2 одинаково часто встречающихся класса получается P c = 0,5.) Для задач, требующих аналогового вывода. значение, производительность обученного считывания измерялась на тестовых примерах по его коэффициенту корреляции с аналоговым целевым выходом.Точность вычислений, достигаемая обученными нейронами считывания из моделей микросхем с ламинарной структурой на основе данных, сравнивается с точностью, достигаемой обученными нейронами считывания из схем управления (черные полосы на рис. 5), чья ламинарная структура связности на основе данных была зашифрована. путем замены исходных и целевых нейронов каждого синаптического соединения в цепи на случайно выбранные нейроны того же типа, то есть возбуждающие или тормозящие нейроны, при условии, что синаптическое соединение не возникает дважды (мы обычно будем называть эти модели схем как аморфные схемы).Обратите внимание, что эта процедура не изменяет общее количество синапсов, выравнивание типа синапса в отношении пре- и постсинаптического типа нейрона, глобальное распределение синаптических весов или других синаптических параметров или наборы нейронов, которые получают афферентные входы или обеспечивают вывод на считывающие нейроны. Структура связности аморфных цепей (за исключением различных вероятностей связи между популяциями возбуждающих и тормозных нейронов и внутри них) идентична структуре графов, изучаемых в классической теории случайных графов (Bollobas 1985) (с 4 вероятностями связи для этих двух популяций). взяты из схем на основе данных).
Рисунок 5
Производительность обученных нейронов с линейным считыванием на уровнях 2/3 и уровне 5 (см. Методы) для различных задач классификации по шаблонам пиков и вычислений, выполняемых на скоростях двух входных потоков, как для моделей ламинарных микросхем на основе данных (серые полосы) и для цепей управления, в которых ламинарная структура была зашифрована (черные полосы). tcl 1/2 ( t ) обозначает ретроактивную классификацию зашумленных паттернов всплесков (входные данные были сгенерированы, как показано на рис.4) во входных потоках 1 или 2, которые были введены в течение предыдущего временного интервала [ t – 30 мс, t ] в 2 класса в соответствии с шаблоном, из которого был сгенерирован каждый образец пиков. tcl 1/2 ( t – Δ t ) относится к более сложной задаче по классификации во время t пикового шаблона перед последним, который был введен в течение временного интервала [ t – 60 мс, t – 30 мс]. Для классификации XOR задача состоит в том, чтобы вычислить в момент времени t = 450 мс XOR меток шаблона (0 или 1) обоих входных потоков, введенных в течение предыдущего временного сегмента [420 мс, 450 мс].Справа результаты вычислений в реальном времени для изменяющихся во времени скоростей стрельбы r 1 ( t ) входного потока 1 и r 2 ( t ) из входной поток 2 (оба состоят из 4 последовательностей импульсов Пуассона с независимо изменяющимися скоростями воспламенения в 2 входных потоках). Белые полосы показывают результаты производительности для 2 целевых функций r 1 ( t ) / r 2 ( t ) и ( r 1 ( t ) – r 2 ( t )) 2 , а полужирные столбцы – производительность нелинейных компонентов этих вычислений в реальном времени в любое время t (на фактических скоростях срабатывания в обоих входных потоках в течение последнего 30 мс).
Рисунок 5
Производительность обученных нейронов с линейным считыванием на уровнях 2/3 и уровне 5 (см. Методы) для различных задач классификации по шаблонам пиков и вычислений, выполняемых на скоростях двух входных потоков, как для ламинарной микросхемы на основе данных модели (серые полосы) и для цепей управления, в которых ламинарная структура была зашифрована (черные полосы). tcl 1/2 ( t ) обозначает ретроактивную классификацию зашумленных паттернов всплесков (входные данные были сгенерированы, как показано на рис.4) во входных потоках 1 или 2, которые были введены в течение предыдущего временного интервала [ t – 30 мс, t ] в 2 класса в соответствии с шаблоном, из которого был сгенерирован каждый образец пиков. tcl 1/2 ( t – Δ t ) относится к более сложной задаче по классификации во время t пикового шаблона перед последним, который был введен в течение временного интервала [ t – 60 мс, t – 30 мс]. Для классификации XOR задача состоит в том, чтобы вычислить в момент времени t = 450 мс XOR меток шаблона (0 или 1) обоих входных потоков, введенных в течение предыдущего временного сегмента [420 мс, 450 мс].Справа результаты вычислений в реальном времени для изменяющихся во времени скоростей стрельбы r 1 ( t ) входного потока 1 и r 2 ( t ) из входной поток 2 (оба состоят из 4 последовательностей импульсов Пуассона с независимо изменяющимися скоростями воспламенения в 2 входных потоках). Белые полосы показывают результаты производительности для 2 целевых функций r 1 ( t ) / r 2 ( t ) и ( r 1 ( t ) – r 2 ( t )) 2 , а полужирные столбцы – производительность нелинейных компонентов этих вычислений в реальном времени в любое время t (на фактических скоростях срабатывания в обоих входных потоках в течение последнего 30 мс).
Рисунок 5 показывает, что схемы на основе данных работают значительно лучше для большинства рассмотренных задач обработки информации, за исключением задач скорости для считывающего нейрона на уровне 5 и задачи tcl 2 ( t −Δ t ) для считывающего нейрона в слоях 2/3 (для которых повышение производительности не было значительным). В частности, потенциальные проекционные нейроны в слоях 2/3 и уровне 5 имеют в ламинарной схеме на основе данных лучший доступ к информации, содержащейся в текущем и предшествующем паттернах спайков из любого афферентного входного потока.Результаты показывают, что потенциальные считывающие нейроны могут классифицировать паттерны спайков от любого афферентного входного потока независимо от одновременно вводимого паттерна спайков в другом входном потоке (и независимо от довольно высокой вариабельности от испытания к испытанию, показанной на рис. 3). Одна интересная деталь может быть замечена для двух задач, связанных с расчетами скоростей стрельбы. Здесь производительность схем, основанных на данных, и схем управления примерно одинакова (см. Белые полосы на рис.5), но показания со слоев 2/3 работают для схем на основе данных значительно лучше на нелинейной составляющей этих вычислений (см. Жирные полосы перед белыми полосами на рис.5). (Эта нелинейная составляющая целевых функций r 1 / r 2 и ( r 1 – r 2 ) 2 в результате вычитания из этих функций [для рассмотренное распределение r 1 , r 2 ] оптимально подобранная линейная функция).
Фактическая производительность, достигаемая с помощью обученных считываний с моделей микросхем, зависит от размера схемы (теоретические результаты предсказывают, что она автоматически улучшится при увеличении размера схемы [Maass and others 2002]).Это показано на рисунке 6 для одной из вычислительных задач, рассмотренных на рисунке 5 (XOR меток шаблонов всплесков из 2 афферентных входных потоков), как для схем на основе данных, так и для схем управления. Рисунок 6 также показывает, что превосходные характеристики схем, основанных на данных, не зависят от размера схемы. Улучшение производительности схем, состоящих из 1000 нейронов, по сравнению со схемами, состоящими из 160 нейронов, несколько меньше для задач скорости. Например, производительность считывающего нейрона в слоях 2/3 или 5, обученных для 2 задач скорости r 1 / r 2 и ( r 1 – r 2 ) 2 увеличивается в среднем только на 25% для схем на основе данных и на 20% для аморфных схем.
Рисунок 6
Производительность (см. Методы) проекционных нейронов в слоях 2/3 и 5 для различных размеров схем, с ламинарной структурой на основе данных и без нее, для вычисления задачи XOR. Эти результаты показывают, что превосходные характеристики схем, основанных на данных, не зависят от размера схемы.
Рисунок 6
Производительность (см. Методы) проекционных нейронов на уровнях 2/3 и уровне 5 для различных размеров схемы, с ламинарной структурой на основе данных и без нее, для вычисления задачи XOR.Эти результаты показывают, что превосходные характеристики схем, основанных на данных, не зависят от размера схемы.
Предыдущие результаты показывают, что микросхемы с ламинарной структурой на основе данных обладают превосходными вычислительными возможностями для большого разнообразия вычислительных задач. Возникает вопрос «почему» это так. Мы подходим к этому вопросу с двух разных точек зрения. Сначала мы исследуем, какие аспекты структуры схемы на основе данных важны для их превосходной производительности.Очевидно, что наша процедура создания аморфных схем разрушает не только слоистую структуру схем на основе данных, но и другие структурные свойства, такие как распределение степеней узлов в нижележащем графе связности и его кластерная структура. Поэтому мы вводим 3 дополнительных типа цепей управления, чтобы проанализировать влияние конкретных конструктивных особенностей на производительность. Во-вторых, мы демонстрируем характерную особенность внутренней динамики этих различных типов схем, которая коррелирует с их вычислительной производительностью.
Мы сначала изучили вычислительное влияние свойств малого мира схем, основанных на данных. Сети малого мира были охарактеризованы Уоттсом и Строгатцем (1998) с помощью двух свойств. У них более высокий коэффициент кластеризации (измеряемый долей непосредственных соседей узлов в графе, которые связаны ссылкой), чем у аморфных цепей, при сохранении сопоставимой средней длины кратчайшего пути. (Обратите внимание, что оба свойства относятся к структуре нижележащего «неориентированного» графа, в котором направленные ребра заменены неориентированными связями.) Основанные на данных модели кортикальных микросхем на самом деле имеют свойства маленького мира в соответствии с этим определением, потому что их коэффициент кластеризации (который имеет значение 36%) на 38% выше, чем в аморфных схемах, тогда как их средняя длина кратчайшего пути примерно такая же. (1,75 ссылки). (По данным Hilgetag и др. [2000], дальнодействующие корковые связи мозга кошек и макак имеют коэффициенты кластеризации 55% и 46% соответственно). Чтобы решить, достаточно ли этих свойств небольшого мира для обеспечения превосходных вычислительных свойств схем на основе данных, мы создали схемы управления, которые имеют такой же размер, коэффициент кластеризации и среднюю кратчайшую длину пути, что и схемы на основе данных по пространственному алгоритм роста, описанный в Kaiser and Hilgetag (2004) (с параметрами α = 4, β = 1.32 и 560 узлов). Впоследствии эти неориентированные графы были преобразованы в ориентированные графы путем случайной замены каждого ребра синапсом (в случайной ориентации) или взаимной синаптической связью с вероятностью, так что общее количество синаптических связей и взаимных синаптических связей идентично соответствующему количеству для схемы на основе данных. (Следует отметить, что эта процедура не воспроизводит такую же долю типов синапсов, как для схем на основе данных и аморфных схем).
Для 3-го типа схемы управления мы рассмотрели схемы, которые имеют такое же распределение входных и исходящих степеней для нейронов, что и схемы, основанные на данных. Входящая и исходящая степень нейрона определяется как общее количество входящих и исходящих синаптических соединений, соответственно.
Для этой цели мы создали схемы на основе данных и впоследствии обменивались целевыми нейронами случайно выбранных пар синапсов с пре- и постсинаптическими нейронами той же категории (возбуждающие или тормозящие), пока не исчезла ламинарно-специфическая структура связности (нет обмен был произведен, если какое-либо из двух образовавшихся новых соединений уже существовало).Этот тип схемы также имеет свойства маленького мира, но средний коэффициент кластеризации был меньше, чем для схем, основанных на данных (всего на 27% выше, чем в аморфных сетях). Мы называем этот тип цепей цепями с регулируемой степенью.
Цепи, управляемые степенями, сохраняют распределение степеней между нейронами, которые получают внешний ввод или обеспечивают ввод для считывающего нейрона. Поэтому мы добавили схемы управления, называемые схемами с контролируемой степенью без специфичности ввода или вывода, путем случайного обмена нейронами на разных уровнях схем с регулируемой степенью.Распределение степеней нейронов для всех 5 типов цепей показано на рисунке 7.
Рисунок 7
Распределение нейронов в разных популяциях для 5 типов цепей (усредненное по 100 цепям). Все схемы имеют распределения степеней, которые могут быть лучше аппроксимированы суммами гауссиан, чем степенной закон вида P ( d ) ∝ d −γ для нейронов степени d и положительная константа γ ( я.е., схемы не свободны от масштаба). Коэффициенты корреляции для наименьших квадратов для сумм гауссианов и степенных распределений составляют> 0,96 и <0,08, соответственно. Таким образом, ни одна из этих схем не является безмасштабной, что показывает, что их различие в производительности не может быть объяснено на основе этой концепции. Вычислительный анализ (см. Таблицу 2) подразумевает, что различные местоположения пиков для разных уровней схем, основанных на данных, важны для их превосходных вычислительных характеристик.
Рисунок 7
Распределение нейронов в разных популяциях для 5 типов цепей (усредненное по 100 цепям). Все схемы имеют распределения степеней, которые могут быть лучше аппроксимированы суммами гауссиан, чем степенной закон вида P ( d ) ∝ d −γ для нейронов степени d и положительная константа γ ( т.е. схемы не свободны от масштаба). Коэффициенты корреляции для наименьших квадратов для сумм гауссианов и степенных распределений> 0.96 и <0,08 соответственно. Таким образом, ни одна из этих схем не является безмасштабной, что показывает, что их различие в производительности не может быть объяснено на основе этой концепции. Вычислительный анализ (см. Таблицу 2) подразумевает, что различные местоположения пиков для разных уровней схем, основанных на данных, важны для их превосходных вычислительных характеристик.
Важной структурной особенностью всех типов цепей, рассмотренных до сих пор, является выравнивание типа синапса относительно пре- и постсинаптического типа нейрона согласно таблице 1.Чтобы проанализировать влияние выравнивания динамических синапсов на производительность, мы случайным образом обменяли триплеты синаптических параметров, то есть U, D и F , которые определяют краткосрочную пластичность между всеми синапсами. В последнем типе схемы управления мы заменили все динамические синапсы статическими синапсами (с измененными весами, чтобы средняя частота срабатывания в слое 5 оставалась неизменной).
Сводные характеристики всех 7 различных типов цепей управления показаны в таблице 2.Свойство малого мира в некоторой степени увеличивает производительность аморфных схем, но более важной структурной особенностью является распределение степеней, определяемое схемами на основе данных. Если это распределение степеней соответствует распределению степеней схем на основе данных для каждого отдельного уровня и, следовательно, совпадает также с конкретной топологией ввода и вывода схем на основе данных, средняя производительность сопоставима с производительностью схем на основе данных. Таблица 2 также показывает (см. Столбец 5), что назначение типов синапсов на основе данных (в соответствии с таблицей 1) важно для хорошей вычислительной производительности.Последний столбец показывает, что схемы со статическими синапсами также имеют худшие вычислительные свойства.
Чтобы выяснить взаимосвязь между внутренними свойствами динамики схемы и вычислительной производительностью, мы более подробно изучили одну фундаментальную, но относительно простую задачу обработки информации: ретроактивную классификацию паттернов спайков на 2 класса, несмотря на шум. Точнее, задача заключалась в том, чтобы классифицировать входные последовательности пиков 2 × 4, сгенерированные из 2 шаблонов (как на рис.4) на 2 класса, несмотря на последующий период ожидания в 100 мс (в течение которого в каждом случае вводились идентичные последовательности пиков), и несмотря на сильно различающиеся начальные условия (вызванные разными предшествующими входными пиками) и относительно высокий внутренний шум, который моделирует бомбардировку несвязанным фоновым синаптическим входом в состоянии с высокой проводимостью (сравните средний и нижний ряды на рис. 3, чтобы увидеть стохастические изменения в спайковой реакции, вызванной последним). Эта задача проверяет способность схем поддерживать информацию о паттернах всплесков, которые были введены более 100 мс назад.Эта информация уменьшается из-за шума, возникающего из-за внутреннего шума в нейронах и изменения начальных условий (внутриклассовая дисперсия). Пунктирные серые линии на рисунке 8 показывают, что считывающие нейроны в слоях 2/3 и уровне 5 схем, основанных на данных, могут довольно быстро обучаться на относительно небольшом количестве обучающих примеров, чтобы угадать, какой из двух фиксированных паттернов спайков был ранее введен. Сравнение с черными линиями показывает, что их ошибка на новых примерах этой задачи (ошибка теста) значительно меньше, чем у считывающих нейронов в аморфных схемах управления.Более того, это преимущество не уменьшается, когда становится доступным больше обучающих примеров. Превосходство считывания из схем на основе данных является результатом как лучшего соответствия обучающим данным (сплошные кривые на рис. 8), так и меньшей ошибки обобщения (= расстояние между сплошной и пунктирной кривой). (Обратите внимание, что все типы схем имеют для меньшего числа обучающих примеров меньшую ошибку на обучающем наборе, но большую ошибку на тестовом наборе из-за хорошо известного эффекта переобучения, который широко изучается в статистической теории обучения [Vapnik 1998] ).
Рисунок 8
Погрешность обучения и тестирования считываний из основанных на данных и аморфных моделей микросхем в зависимости от размера обучающей выборки. Задача заключалась в обратной классификации 2 случайно созданных паттернов спайков длиной 100 мс (состоящих из 4 последовательностей спайков Пуассона с частотой 20 Гц) после того, как идентичный ввод длительностью 100 мс был впоследствии введен в схему, несмотря на различные начальные условия (вызванные независимыми генерируются предыдущие входные сигналы пуассоновских всплесков того же типа) и шумные фоновые входные токи, как и раньше.На верхней панели показаны характеристики потенциального считывающего нейрона в слоях 2/3 с 84 пресинаптическими нейронами, обученными с помощью линейной регрессии с ограничениями по знакам (см. Методы). На нижней панели показаны характеристики потенциального считывающего нейрона в слое 5 со 109 пресинаптическими нейронами. Оба типа считывания лучше работают для ламинарной схемы на основе данных как на обучающем наборе (с новыми случайными рисунками начальных условий и фонового шума), так и на тестовом наборе. Это справедливо для всех рассматриваемых размеров обучающей выборки.
Рисунок 8
Ошибка обучения и тестирования считываний из моделей на основе данных и моделей аморфных микросхем в зависимости от размера обучающей выборки. Задача заключалась в обратной классификации 2 случайно созданных паттернов спайков длиной 100 мс (состоящих из 4 последовательностей спайков Пуассона с частотой 20 Гц) после того, как идентичный ввод длительностью 100 мс был впоследствии введен в схему, несмотря на различные начальные условия (вызванные независимыми генерируются предыдущие входные сигналы пуассоновских всплесков того же типа) и шумные фоновые входные токи, как и раньше.На верхней панели показаны характеристики потенциального считывающего нейрона в слоях 2/3 с 84 пресинаптическими нейронами, обученными с помощью линейной регрессии с ограничениями по знакам (см. Методы). На нижней панели показаны характеристики потенциального считывающего нейрона в слое 5 со 109 пресинаптическими нейронами. Оба типа считывания лучше работают для ламинарной схемы на основе данных как на обучающем наборе (с новыми случайными рисунками начальных условий и фонового шума), так и на тестовом наборе. Это справедливо для всех рассматриваемых размеров обучающей выборки.
Более внутреннее объяснение лучшей вычислительной производительности ламинарных схем на основе данных дает теория вычислений в динамических системах (для обзора см. Legenstein and Maass 2005). На рисунке 9 показано, что схема на основе данных работает в значительно менее хаотическом режиме, чем аморфная схема управления. Его чувствительность к незначительным различиям в начальных условиях также меньше, чем у других трех типов схем управления, которые сохраняют выбранные аспекты структуры сети на основе данных.Менее хаотическая динамика подразумевает лучшую способность обобщения новых входных данных для многих различных типов динамических систем. Это наблюдение представляет интерес, поскольку можно предположить, что количество синапсов на нейрон является важным параметром, определяющим степень хаоса в цепи. Но все схемы, результаты которых представлены на рисунке 9, имеют одинаковое количество синапсов.
Рисунок 9
Евклидовы расстояния в траекториях состояний схемы (точнее: от входных векторов до считываемых нейронов в слоях 2/3 и в слое 5) в результате перемещения одиночного входного пика (на 100 мс) на 0.5 мс. Показано среднее значение моделирования 400 случайно сгенерированных цепей с различными начальными условиями и фоновым шумом. Начальные условия и внутренний шум были выбраны идентичными в обоих испытаниях каждого моделирования, как и в стандартных тестах для оценки показателя Ляпунова (детерминированных) динамических систем, см. Legenstein and Maass (2005). Кривые показывают устойчивые различия в евклидовом расстоянии между состояниями цепей, которые примерно в два раза больше для аморфных цепей, тем самым указывая на более хаотическую динамику, чем в ламинарных цепях с тем же количеством нейронов и синапсов.
Рисунок 9
Евклидовы расстояния в траекториях состояний схемы (точнее: от входных векторов до считывающих нейронов в слоях 2/3 и в слое 5) в результате перемещения одиночного входного пика (на 100 мс) на 0,5 мс. Показано среднее значение моделирования 400 случайно сгенерированных цепей с различными начальными условиями и фоновым шумом. Начальные условия и внутренний шум были выбраны идентичными в обоих испытаниях каждого моделирования, как и в стандартных тестах для оценки показателя Ляпунова (детерминированных) динамических систем, см. Legenstein and Maass (2005).Кривые показывают устойчивые различия в евклидовом расстоянии между состояниями цепей, которые примерно в два раза больше для аморфных цепей, тем самым указывая на более хаотическую динамику, чем в ламинарных цепях с тем же количеством нейронов и синапсов.
Сплошные кривые на рисунке 8 показывают, что еще одна причина лучшей вычислительной производительности моделей схем на основе данных заключается в том, что синаптические веса их считывающих нейронов могут лучше соответствовать обучающим данным. Этот факт можно объяснить с точки зрения «классовой дисперсии» состояний схемы большой размерности, вызванной изменением начальных условий и внутреннего шума (для повторных испытаний с одним и тем же импульсом на входе схемы).Корреляция между этой классовой дисперсией обучающих данных и ошибкой классификации обученных считываний тестовых данных для задачи, рассмотренной на рисунке 8, составляла 0,80 для показаний из слоев 2/3 и 0,72 для показаний из уровня 5 для ламинарных схем на основе данных. .
На рисунке 10 показано, что для аморфных цепей управления это внутриклассовое отклонение обычно больше. Кроме того, на рисунке 10 показано, что эта функция подавления шума динамики в ламинарных схемах на основе данных не является случайным свойством фиксированной настройки трех параметров S RW , S I1 , S I2 (который масштабирует веса повторяющихся синаптических соединений, амплитуды входного потока 1 и амплитуды входного потока 2), который мы использовали для моделирования, о котором сообщалось до сих пор (см. Методы).На рисунке 10 показано, что эта функция подавления шума появляется также для всех других (случайно выбранных) настроек этих параметров, которые были протестированы.
Рисунок 10
Структура схемы на основе данных снижает влияние шума. Для каждого из 2 типов схем, то есть для схем на основе данных и аморфных схем, было сгенерировано 50 схем, и 500 идентичных входов, сгенерированных для задачи на рисунке 8, были введены в каждую из них. Разница в дисперсии откликов цепей (в среднем по 50 цепям) оценивалась с точки зрения считывающих нейронов.Точнее, мы измерили дисперсию входных данных для считывающих нейронов после 500-кратного ввода того же входа в схему и проанализировали, насколько эта дисперсия уменьшилась для структуры схемы на основе данных (выраженной в процентах изменения по сравнению с аморфной структурой). схемы управления). Меньшая дисперсия для схем, основанных на данных, показывает, что на их динамику меньше влияют внутренний шум и различные начальные условия, тем самым обеспечивая лучшие возможности обобщения обученных считываний.Этот эксперимент был повторен для 30 случайно выбранных настроек параметров масштабирования S RW , S I1 , S I2 (см. Методы), и на этом рисунке показано, сколько из этих настроек параметра a определенное снижение шума было достигнуто для схем, основанных на данных. Среднее отклонение в классе для всех настроек параметров составило 0,58, а стандартное отклонение – 0,29 для схем на основе данных. Проценты для 5 из этих цепей не были введены в этот график, потому что их классовая дисперсия была ниже 0.01 из-за того, что они почти не ответили на ввод. Процент изменения классовой дисперсии для стандартной настройки параметров масштабирования составил -38 для показаний уровня 2/3 и -41 для показаний уровня 5. Эти результаты показывают, что способность схем на основе данных снижать уровень шума не была случайным свойством стандартной настройки параметров масштабирования.
Рисунок 10
Структура схемы на основе данных снижает влияние шума. Для каждого из 2 типов схем, то есть для схем на основе данных и аморфных схем, было сгенерировано 50 схем, и 500 идентичных входов, сгенерированных для задачи на рисунке 8, были введены в каждую из них.Разница в дисперсии откликов цепей (в среднем по 50 цепям) оценивалась с точки зрения считывающих нейронов. Точнее, мы измерили дисперсию входных данных для считывающих нейронов после 500-кратного ввода того же входа в схему и проанализировали, насколько эта дисперсия уменьшилась для структуры схемы на основе данных (выраженной в процентах изменения по сравнению с аморфной структурой). схемы управления). Меньшая дисперсия для схем, основанных на данных, показывает, что на их динамику меньше влияют внутренний шум и различные начальные условия, тем самым обеспечивая лучшие возможности обобщения обученных считываний.Этот эксперимент был повторен для 30 случайно выбранных настроек параметров масштабирования S RW , S I1 , S I2 (см. Методы), и на этом рисунке показано, сколько из этих настроек параметра a определенное снижение шума было достигнуто для схем, основанных на данных. Среднее отклонение в классе для всех настроек параметров составило 0,58, а стандартное отклонение – 0,29 для схем на основе данных. Проценты для 5 из этих цепей не были введены в этот график, потому что их классовая дисперсия была ниже 0.01 из-за того, что они почти не ответили на ввод. Процент изменения классовой дисперсии для стандартной настройки параметров масштабирования составил -38 для показаний уровня 2/3 и -41 для показаний уровня 5. Эти результаты показывают, что способность схем на основе данных снижать уровень шума не была случайным свойством стандартной настройки параметров масштабирования.
Рисунок 11 показывает, что превосходная вычислительная производительность моделей ламинарных микросхем на основе данных не является случайным следствием конкретного выбора этих трех параметров, но сохраняется для большинства их потенциальных значений.Этот факт демонстрируется здесь для XOR на паттернах спайков, которые уже обсуждались ранее. (Стандартное отклонение производительности считываний от схем управления для различных настроек параметров составляло 0,15 для показаний со слоев 2/3 и 0,18 для показаний со слоя 5. Улучшение производительности для ламинарных схем на основе данных в некоторой степени коррелировало с производительностью [коэффициент корреляции 0,16 для слоев 2/3, 0,59 для показаний слоя 5]). Это говорит о том, что ламинарная схема имеет для некоторых задач превосходные вычислительные возможности для довольно большого изменения динамических режимов.Это представляет интерес, потому что разные поведенческие состояния, разные состояния гомеостатических процессов или разные входные интенсивности могут привести к множеству различных динамических режимов корковых микросхем.
Рисунок 11
Процент улучшения производительности считывания из ламинарных цепей для различных значений трех параметров масштабирования для той же задачи XOR, которая обсуждается на рисунках 4 и 5 (но только с 2 × 4 последовательностями входных пиков). Процент улучшения был измерен для 16 случайно выбранных настроек параметров масштабирования S RW , S I1 и S I2 .Две из этих настроек дали чрезвычайно низкую производительность как для схем, основанных на данных, так и для аморфных схем (ниже 0,08, следовательно, ниже SD данных производительности для всех 16 настроек параметров) и поэтому были исключены из графика. Улучшение производительности при стандартной настройке этих параметров составило 74% для показаний слоев 2/3 и 64% для показаний уровня 5. Это говорит о том, что ламинарная схема обладает превосходными вычислительными возможностями для большинства настроек параметров, следовательно, для широкого разнообразия интенсивности стимулов и регуляторных состояний нейронных систем in vivo.
Рисунок 11
Процент повышения производительности считывания из ламинарных цепей для различных значений трех параметров масштабирования для той же задачи XOR, как показано на рисунках 4 и 5 (но только с 2 × 4 последовательностями входных пиков). Процент улучшения был измерен для 16 случайно выбранных настроек параметров масштабирования S RW , S I1 и S I2 . Две из этих настроек дали чрезвычайно низкую производительность как для схем на основе данных, так и для аморфных схем (ниже 0.08, следовательно, ниже стандартного отклонения рабочих характеристик для всех 16 значений параметров) и поэтому не были включены в график. Улучшение производительности при стандартной настройке этих параметров составило 74% для показаний слоев 2/3 и 64% для показаний уровня 5. Это говорит о том, что ламинарная схема обладает превосходными вычислительными возможностями для большинства настроек параметров, следовательно, для широкого разнообразия интенсивности стимулов и регуляторных состояний нейронных систем in vivo.
Обсуждение
Мы продемонстрировали, что ламинарная структура связности на основе данных расширяет возможности обработки информации в моделях кортикальных микросхем.В частности, мы показали, что такая модель схемы на основе данных может накапливать, удерживать и объединять информацию, содержащуюся в двух афферентных входных потоках пиков. Следует отметить, что вычисления, которые были проанализированы в наших компьютерных экспериментах, были биологически реалистичными вычислениями в реальном времени для динамически изменяющихся входных потоков, а не статическими вычислениями для пакетных входных данных, которые обычно рассматриваются в исследованиях моделирования. В отличие от схемных моделей из Buonomano и Merzenich (1995) и Maass и др. (2002), схемные модели, которые были проанализированы в этой статье, не только имеют биологически реалистичную ламинарную структуру, но также состоят из нейронов типа Ходжкина-Хаксли (с дополнительными фоновый ввод основан на данных, которые предположительно являются репрезентативными для состояния высокой проводимости корковых цепей in vivo [Destexhe and others 2003]).Кроме того, моделирование, обсуждаемое в этой статье, учитывает существенно большую вариабельность от испытания к испытанию. Кроме того, информация извлекалась не из всех нейронов, как у Buonomano и Merzenich (1995) и Maass и др. (2002), а из гораздо меньшего подмножества нейронов, которое представляет собой типичный набор пресинаптических нейронов для проекционного нейрона в слоях 2/3 или 5. Кроме того, получение информации такими проекционными нейронами впервые было связано с ограничением, согласно которому знаки веса входящих синапсов не могут выбираться произвольно в биологической цепи, а определяются типом (возбуждающим или тормозящим) каждый пресинаптический нейрон.Хотя это означает, что не вся мощность линейной регрессии (или правила обучения персептрона) может быть использована для оптимизации таких более реалистичных результатов, мы показываем, что даже в этих биологически более реалистичных условиях значительный объем информации может быть извлечен проекционными нейронами. в слоях 2/3 или уровне 5. Кроме того, результаты на Рисунке 6 показывают, что их производительность увеличивается с увеличением размера схемы, что позволяет предположить, что почти идеальная производительность будет достигнута с помощью модели схемы, которая достаточно велика, так что количество пресинаптические нейроны приближаются к реальным значениям в несколько тысяч.
Мы продемонстрировали на рисунке 5, что модели ламинарных микросхем на основе данных работают значительно лучше, чем схемы управления (которые не имеют ламинарных структур, но в остальном идентичны в отношении своих компонентов и общей статистики соединений) для широкого спектра фундаментальной информации: задачи обработки. Это превосходство сохраняется для большинства настроек параметров, которые масштабируют глобальную силу афферентных входов и повторяющихся связей, соответствующих широкому диапазону интенсивности стимулов и регуляторных состояний нейронных систем in vivo (рис.11). Мы также проанализировали, какой аспект структуры связности ламинарных схем на основе данных отвечает за их лучшую вычислительную производительность. Мы пришли (на основе результатов, представленных в Таблице 2) к выводу, что их особое распределение степеней узлов (относительно входов схемы и проекционных нейронов) является основной причиной, в большей степени, чем свойство ограниченного мира данных – на основе схем. Мы предполагаем, что это вычислительное превосходство ламинарных схем можно понять с точки зрения свойств динамической системы, которая определяется такими моделями микросхем.Мы показали на рисунках 9 и 10, что динамика ламинарных цепей несколько меньше зависит от внутреннего шума и шума на входе, тем самым обеспечивая лучшие возможности обобщения обученных считываний и лучшее соответствие обучающим данным из-за меньшей дисперсии в ответах цепей. .
По-видимому, модели нейронных цепей, рассмотренные в этой статье, представляют собой наиболее подробные модели корковых микросхем, основанные на данных, возможности обработки информации которых были проанализированы до сих пор.Результаты этого анализа показывают, что с помощью обширного компьютерного моделирования можно продемонстрировать конкретные вычислительные последствия их ламинарной структуры, тем самым создавая связь между подробными анатомическими и нейрофизиологическими данными и их вероятными вычислительными последствиями. Мы ожидаем, что эта программа может быть продолжена для выяснения также функциональных последствий дальнейших деталей кортикальных микросхем, таких как описанные, например, у Гупты и др. (2000), Штайгера и др. (2000), Шуберта и др. (2001), Бинцеггера. и другие (2004 г.), Каллавей (2004 г.), Маркрам и другие (2004 г.) и Йошимура и другие (2005 г.).
Мы хотели бы поблагодарить Эда Каллэуэя, Родни Дугласа, Рольфа Кеттера, Генри Маркрама и Алекса Томсона за полезные обсуждения исследований, связанных с этой статьей. Работа частично поддержана австрийским научным фондом Fonds zur Förderung der Wissenschaftlichen Forschung (FWF), проект # P15386 и # S9102-N04, проект анализа паттернов, статистического моделирования и вычислительного обучения # IST2002-506778 и Fast Analog Computing with Emergent Проект “Переходные государства” № FP6-015879 Европейского Союза.
Список литературы
,.Модель глобальной спонтанной активности и локальной структурированной активности в периоды задержки в коре головного мозга
,Cereb Cortex
,1997
, vol.7
3
(стр.237
–252
),,,,.Зависимость от стимула двухуровневых колебаний мембранного потенциала в зрительной коре головного мозга кошки
,Nat Neurosci
,2000
, vol.3
6
(стр.617
–621
),,.Поток возбуждения в коре головного мозга крысы при столкновении с одиночной вибриссой
,Дж. Нейрофизиол
,1992
, т.68
4
(стр.1345
–1358
),,,,,,.Ответы нейронов первичной и нижней височной зрительной коры на естественные сцены
,Proc R Soc Lond B Biol Sci
,1997
, vol.264
1389
(стр.1775
–1783
),,.Количественная карта цепи первичной зрительной коры кошки
,J Neurosci
,2004
, vol.24
39
(стр.8441
–8453
). ,Случайные графики
,1985
Лондон
Academic Press
,,.Визуальный ввод вызывает кратковременное и сильное подавление шунтирования в зрительных корковых нейронах
,Nature
,1998
, vol.393
6683
(стр.369
–373
),.Временная информация, преобразованная в пространственный код нейронной сетью с реалистичными свойствами
,Science
,1995
, vol.267
(стр.1028
–1030
).Прямая связь, обратная связь и тормозящие связи в зрительной коре головного мозга приматов
,Neural Netw
,2004
, vol.17
5–6
(стр.625
–632
),.Настройка силы и ориентации таламического входа к простым клеткам, выявленная электрически вызванным кортикальным подавлением
,Neuron
,1998
, vol.20
(стр.1177
–1189
),.Ламинарные источники синаптического входа в корковые тормозные интернейроны и пирамидные нейроны
,Nat Neurosci
,2000
, vol.3
7
(стр.701
–707
),.Пластичность в вычислениях одиночных нейронов и схем
,Nature
,2004
, vol.431
(стр.789
–795
),.Влияние сетевой активности на интегративные свойства пирамидных нейронов неокортекса in vivo
,J Neurophysiol
,1999
, vol.81
4
(стр.1531
–1547
),,,.Колеблющиеся синаптические проводимости воссоздают in vivo-подобную активность нейронов неокортекса
,Neuroscience
,2001
, vol.107
1
(стр.13
–24
),,.Состояние высокой проводимости нейронов неокортекса in vivo
,Nat Rev Neurosci
,2003
, vol.4
9
(стр.739
–751
),,,,.Рекуррентное возбуждение в контурах неокортекса
,Science
,1995
, vol.269
5226
(стр.981
–985
),.Нейронные цепи неокортекса
,Annu Rev Neurosci
,2004
, vol.27
(стр.419
–451
).Происхождение ориентационно-селективных ВПСП в простых клетках зрительной коры головного мозга кошек
,J Neurosci
,1987
, vol.7
6
(стр.1780
–1791
),,.Принципы организации разнообразных ГАМКергических интернейронов и синапсов в неокортексе
,Science
,2000
, vol.287
(стр.273
–278
),,,.Анатомическая взаимосвязь определяет организацию кластеров кортикальных областей у макак и кошек.
,Philos Trans R Soc Lond B Biol Sci
,2000
, vol.355
(стр.91
–110
),,,.Синаптическая интеграция в полосатых простых клетках коры
,J Neurosci
,1998
, vol.18
22
(стр.9517
–9528
),,,.K + канал регуляции распространения сигнала в дендритах пирамидных нейронов гиппокампа
,Nature
,1997
, vol.387
6636
(стр.869
–875
),.Пространственный рост реальных сетей
,Phys Rev E
,2004
, vol.69
стр.036103
,,.Неокортикальная микросхема как tabula rasa
,Proc Natl Acad Sci USA
,2005
, vol.102
3
(стр.880
–885
),.Линейный метод наименьших квадратов с ограничениями линейного неравенства
,Решение задач наименьших квадратов
,1974
Энглвуд Клиффс (Нью-Джерси)
Прентис-Холл
стр.161
,. ,,,.Что делает динамическую систему вычислительно мощной?
,Новые направления в статистической обработке сигналов: от систем к мозгу
,2005
MIT Press
,,.Принципы вычислений в реальном времени с обратной связью, применяемые к моделям кортикальных микросхем
,Proceedings of Advances in Neural Information Processing Systems
,2005
MIT Press
,.Синапсы как буферы динамической памяти
,Neural Netw
,2002
, vol.15
(стр.155
–161
),,.Вычисления в реальном времени без стабильных состояний: новая структура для нейронных вычислений на основе возмущений
,Neural Comput
,2002
, vol.14
11
(стр.2531
–2560
),,,.Электрическая и кальциевая передача сигналов в дендритах пирамидных нейронов гиппокампа
,Annu Rev Physiol
,1998
, vol.60
(стр.327
–346
),.Характеристика одиночных потенциалзависимых каналов Na + и Ca2 + в апикальных дендритах пирамидных нейронов CA1 крысы
,J Physiol
,1995
, vol.487
Pt 1
(стр.67
–90
),,,.Модель инициирования спайков в пирамидных нейронах неокортекса
,Neuron
,1995
, vol.15
6
(стр.1427
–1439
),,,,,.Интернейроны неокортикальной тормозной системы
,Nat Rev Neurosci
,2004
, vol.5
10
(стр.793
–807
),,.Дифференциальная передача сигналов через один и тот же аксон пирамидных нейронов неокортекса
,Proc Natl Acad Sci USA
,1998
, vol.95
(стр.5323
–5328
). ,Перцептивная нейробиология: кора головного мозга
,1998
Кембридж, Массачусетс
Harvard University Press
,,. .Компьютерные модели и средства анализа нейронных микросхем
,Базы данных нейронауки. Практическое руководство
,2003
Boston
Kluwer Academic Publishers
(стр.123
–138
).Корковые микросхемы: разнообразны или каноничны?
,Нейрон
,2002
, т.36
1
(стр.19
–27
),.К теории ламинарной архитектуры коры головного мозга: вычислительные ключи от зрительной системы
,Cereb Cortex
,2003
, vol.13
1
(стр.100
–113
),,,,,.Слоистая внутриколоночная и трансколонарная функциональная связность пирамидных клеток слоя v в коре головного мозга крысы
,J Neurosci
,2001
, vol.21
10
(стр.3580
–3592
),,.Стереотипия в микросхемах неокортекса
,Trends Neurosci
,2002
, vol.25
5
(стр.227
–230
),,,.Ламинарные характеристики функциональной связности в коре головного мозга крысы, выявленные при стимуляции глутаматом в клетке
,Neurosci Res
,2000
, vol.37
1
(стр.49
–58
),.Активное распространение соматических потенциалов действия в дендриты пирамидных клеток неокортекса
,Nature
,1994
, vol.367
6458
(стр.69
–72
).Кросс-корреляционный анализ взаимоотношений геникулостриатных нейронов у кошек
,J Neurophysiol
,1983
, vol.49
(стр.1303
–1318
).Пресинаптическая частотно-зависимая фильтрация и шаблон
,J Comput Neurosci
,2003
, vol.15
(стр.159
–202
),,,.Синаптические связи и небольшие цепи с участием возбуждающих и тормозных нейронов в слоях 2–5 неокортекса взрослых крыс и кошек: тройная внутриклеточная запись и мечение биоцитином in vitro
,Cereb Cortex
,2002
, vol.12
9
(стр.936
–953
),. ,Нейронные сети гиппокампа
,1991
Кембридж, Массачусетс
Cambridge University Press
.Вычислительные ограничения, которые могли способствовать расслоению сенсорной коры
,J Comput Neurosci
,2003
, vol.14
(стр.271
–282
). ,Статистическая теория обучения
,1998
Нью-Йорк
Джон Вили
,.Хаос в нейронных сетях со сбалансированной возбуждающей и тормозной активностью
,Science
,1996
, vol.274
(стр.1724
–1726
),.Хаотическое сбалансированное состояние в модели корковых цепей
,Neural Comput
,1998
, т.10
6
(стр.1321
–1371
),.Коллективная динамика сетей «маленького мира»
,Nature
,1998
, vol.393
(стр.440
–442
). ,Кортикальные контуры
,1989
Boston
Birkhaeuser
,,.Возбуждающие корковые нейроны образуют мелкомасштабные функциональные сети
,Nature
,2005
, vol.433
7028
(стр.868
–873
)Заметки автора
© Автор, 2006. Опубликовано Oxford University Press. Все права защищены. Для получения разрешений обращайтесь по электронной почте: [email protected]
Онлайн-версия этой статьи была опубликована в рамках модели открытого доступа. Пользователи имеют право использовать, воспроизводить, распространять или отображать версию этой статьи в открытом доступе в некоммерческих целях при условии, что: исходное авторство правильно и полностью указано; Журнал и Издательство Оксфордского университета указываются как место первоначальной публикации с указанием правильных сведений о цитировании; если статья впоследствии воспроизводится или распространяется не полностью, а только частично или как производная работа, это должно быть четко указано.По вопросам коммерческого повторного использования обращайтесь по адресу [email protected].
Модуляция делительного усиленияобеспечивает гибкое и быстрое вовлечение в неокортикальную микросхему модели
Модель неокортикальной микросхемы.
Модель, использованная в этом исследовании, является расширенной версией пространственно локализованной модели Уилсона – Коуэна (Wilson and Cowan, 1972). Эта модель неокортикальной микросхемы включает одну возбуждающую популяцию, E , и две ингибирующие субпопуляции: одну нацеленную на дендриты, I dend и одну нацеленную на сому межнейронную популяцию, I сома (см.рис.1 А ). Две ингибирующие субпопуляции различаются типом ингибирующей модуляции, которую они обеспечивают возбуждающей популяции. I dend всегда обеспечивает субтрактивное торможение, то есть субтрактивную модуляцию функции ввода-вывода своей цели: сдвиг кривой к более высоким входным значениям. I soma может обеспечивать комбинацию вычитающего и разделяющего ингибирования, то есть модуляции делительного усиления, на возбуждающую популяцию: уменьшая наклон и максимальный выходной сигнал функции (см. Схемы модуляции на рис.1 А ).
Рис. 1. Модель неокортикальной микросхемы и меры увлечения. A : схематическая модель цепи неокортекса. Он включает одну возбуждающую ( E ) и две ингибирующие субпопуляции, одну нацеленную на дендриты ( I dend ) и одну межнейронную популяцию, нацеленную на сому ( I сома ), и веса ( w ). их связей. Обратите внимание, что характер подавления, передаваемого от I сома до E , определяется параметром делимости 0 ≤ q ≤ 1 (показано синим).Он варьируется от полностью вычитающей ( q = 0) до полностью разделяющей ( q = 1) модуляции. Также показаны схемы этих двух типов модуляции ввода-вывода. B : пример захвата. Первоначально схема получает постоянный входной сигнал, P ( t ) = P , и колеблется на своей собственной частоте, f ∗ [представленной активностью возбуждающего населения E ( t )]. Входной сигнал становится синусоидальной волной с амплитудой Λ и входной частотой f в .Микросхема успешно увлекается входной частотой. C : увлечение микросхемы может занять несколько циклов колебаний, как показано в этом примере. Время сходимости рассчитывается как время, необходимое для схождения активности до конечного предельного цикла. Затем рассчитывается индекс увлечения из спектра мощности сигнала E ( t ) после его схождения.
Исходная модель Уилсона – Коуэна имеет субтрактивную модуляцию, но, чтобы ввести в модель делительную модуляцию, нам нужно определить сигмоидальную функцию ввода-вывода, которая принимает три переменные: вход или привод, x ; субтрактивная модуляция,; и делительная модуляция, A:
Fe, ix, Θ, Α = αe, iαe, i + qΑ11 + exp − αe, ix − θe, i + Θ + 1 − qΑ − 11 + expαe, iθe, i, ( 1)
, где e обозначает возбуждающее, а i обозначает тормозящее.Функция определена со всеми входными данными, которые являются действительными неотрицательными значениями. Константа θ e, i – это минимальное смещение функции ввода-вывода по оси x для соответствующей совокупности, представляющее состояние по умолчанию, когда нет субтрактивного запрета, который смещает функцию дальше. Константа α e, i – это максимальное усиление сигмоидальной функции для соответствующей совокупности, представляющее состояние по умолчанию, когда нет разделяющего запрета, уменьшающего усиление функции.Обратите внимание, как параметр делимости q ∈ [0, 1] управляет эффектом делительной модуляции A. Только тогда, когда q = 1, модуляция является чисто разделительной. Более низкое значение приводит к комбинации вычитающей и делительной модуляции, и она становится чисто вычитающей с q = 0. Кроме того, обратите внимание, что когда A = 0, эта функция эквивалентна сигмоидальной функции, используемой в исходной модели Уилсона – Коуэна. модель (Уилсон и Коуэн, 1972). Определенная здесь функция ввода-вывода является разновидностью функции, используемой в Papasavvas et al.(2015), и мы объяснили его точное происхождение в Papasavvas and Wang (2019).С помощью функции ввода-вывода F e, i микросхема неокортекса описывается следующей системой обыкновенных дифференциальных уравнений:
τdEtdt = −Et + ke − EtFew1Et + Pt, w2Idendt, w3Isomat,
τdIdendtdt = −Idendt + ki − IdendtFiw4Et, 0,0,
τdIsomatdt = −Isomat + ki − IsomatFiw5Et, w6Idendt + w7Isomat, 0, (2)
где, аналог исходной модели Уилсона – Коуэна, limx → ∞Fe, ix, 0,0 = expαe, iθe, i1 + expαe, iθe, i.(3)Рис. 1 A показывает схему неокортикальной микросхемы с указанием возбуждающих и тормозных связей и их веса, w . Чтобы ограничить параметры этой модели, мы использовали данные, полученные в первичной зрительной коре Pfeffer et al. (2013). Во-первых, I dend подавляет I сома , но не взаимно. Во-вторых, для I сома существует самоторможение, но не для I dend .В-третьих, некоторые значения веса установлены относительно w 3 : w 7 = w 3 , w 2 = 0,54 w 3 и w 6 = 0,33 w 3 .
Параметры, используемые для функций ввода-вывода F e, i ( уравнение 1 ): θ e = 4, θ i = 3,7, α e = 1.3 и α i = 2. Это значения, использованные в исходной модели Уилсона – Коуэна (Wilson and Cowan, 1972). Постоянная времени была установлена на τ = 0,05. Это значение было выбрано на основе реакции модели неокортикальной микросхемы на мгновенный ввод (см. Дополнительный рис. S1; см. Https://doi.org/dkwh), который имеет характерный период полураспада t 1 / 2 = 39,8 мс, что приблизительно соответствует реакции электрически стимулированных микросхем неокортекса в электрофизиологических экспериментах (Alfonsa et al.2015).
Модель микросхемы была специально разработана для исследования влияния разделяющего ингибирования, которое доставляется популяцией ингибиторов нацеливания на сому I сома , на захват микросхемы. Параметр деления q включен в модель для создания континуума между двумя крайностями: от чисто субтрактивного ингибирования до чисто разделяющего ингибирования, обеспечиваемого I сома . Изменяя параметр q от 0 до 1, мы можем количественно исследовать, используя соответствующие меры (см. Ниже), как на увлечение влияет систематическое увеличение разделяющей способности тормозящей модуляции.Обратите внимание, что параметр деления управляет только модуляцией, передаваемой I сома на E , в то время как I dend всегда обеспечивает чисто субтрактивную модуляцию, и это связано с тем, что модуляция делительного усиления была связана с нацеливанием на сома. интернейроны в неокортексе (Atallah et al. (2012); Pouille et al. (2009); Wilson et al. (2012); однако см. также Seybold et al. (2015)). Несмотря на то, что разделение обеспечивается исключительно I сома , мы по-прежнему включаем I dend в модель, чтобы можно было сравнить с предыдущими результатами моделирования.
Модели нейронной массы могут работать во многих различных режимах, даже при постоянном вводе. Систематический анализ их динамики показал, что они могут находиться в (устойчивом) стабильном состоянии, либо в устойчивой фиксированной точке, либо в стабильном предельном цикле (Grimbert and Faugeras 2006; Spiegler et al. 2010; Wilson and Cowan 1972). Они также могут находиться в предельно стабильной фиксированной точке, то есть в критической точке между поглощающим состоянием и активированным состоянием, что отражает свидетельство критичности экспериментальных данных (Cowan et al.2016). Даже хаотические режимы могут быть достигнуты в модели нейронной массы как минимум с тремя переменными (Papasavvas et al. 2015; Spiegler et al. 2011). Однако в этом исследовании, как и в предыдущей работе (Spiegler et al. 2011), мы исследуем явление уноса только в режиме стабильного предельного цикла, то есть при существующем колебании перед любым колебательным движением. Это связано с тем, что эффект увлечения в биологическом смысле обычно рассматривается как управляемая вводом адаптация уже существующего колебания.
Выборка пространства параметров и поиск локальных оптимумов.
Чтобы исследовать влияние параметра делимости q на развлекательные мероприятия, мы должны были установить параметры микросхемы так, чтобы ее собственная колебательная активность [реакция на постоянный входной сигнал P ( t ) = P . ] при q = 0 и q = 1 сопоставимо. Мы разработали стратегию выборки пространства параметров для параметров { P , w 1 , w 3 , w 4 , w 5 } и создания коллекции допустимых наборов параметров.Другие параметры подключения { w 2 , w 6 , w 7 } устанавливаются относительно w 3 (см. Выше). Чтобы набор параметров считался допустимым для исследования, он должен соответствовать следующим условиям:
Он следует отношениям связи, полученным из Pfeffer et al. (2013).
Он вызывает колебательное поведение как для q = 0, так и для q = 1 с постоянным входом, P ( t ) = P .
Разница между размахом амплитуды (см. Рис.1) E ( t ) при q = 0 и q = 1 меньше 0,001.
Размах амплитуды I сома ( t ) по крайней мере такой же большой, как размах амплитуды E ( t ) на обоих q = 0 и q = 1.
Обратите внимание, что необходимо применить ограничение на амплитуду I сома ( t ) ( условие 4 ), потому что существует возможность подключения параметры, уменьшающие амплитуду I сом ( t ) при сохранении колебаний в двух других популяциях.В этом случае система сводится к классической двумерной системе E – I (Wilson and Cowan 1972). Аналогичное ограничение на I dend ( t ) не является необходимым, поскольку устойчивые колебания в микросхеме невозможны при пониженной активности в I dend ( t ) при q = 1. . Чтобы создать набор таких наборов параметров, мы случайным образом выбрали гиперкуб в пространстве параметров, определяемом интервалами: P ∈ [0, 5] и w k ∈ [0, 35] для k ∈ {1, 3, 4, 5}.После того, как мы сгенерировали остальные параметры на основе условие 1 , система была смоделирована, и были сохранены только те образцы, которые соответствовали условию 2 . Эти наборы затем использовались в качестве отправных точек для задачи оптимизации. Задача оптимизации заключалась в том, чтобы минимизировать разницу между размахом амплитуды E ( t ) при q = 0 и q = 1 ( условие 3 ) при сохранении размаха до пиковая амплитуда I сома ( t ) по крайней мере такая же, как размах амплитуды E ( t ) ( условие 4 ).Во время оптимизации использовался штрафной член, чтобы гарантировать, что условие 4 выполняется и что параметры не отклоняются от гиперкуба, определенного выше. Задача оптимизации решалась с помощью решателя нелинейного программирования fminsearch в MATLAB.
Действительный набор параметров был первоначально создан с помощью этого метода и использован в качестве примера набора для результатов на рис. 2 и 3. Набор примеров: { P = 1,428, w 1 = 24,368, w 3 = 9.677, w 4 = 27,249, w 5 = 30,913}. Другие параметры подключения устанавливаются относительно w 3 , то есть w 2 = 5,225, w 6 = 3,193 и w 7 = 9,677.
С целью обобщения результатов по различным областям пространства параметров мы создали коллекцию допустимых наборов после обширной выборки. Случайная выборка из 250 000 наборов параметров сгенерировала набор из 1227 наборов параметров, которые соответствуют условиям 1 и 2 .Из этих наборов 981 считался уникальным с допуском 0,05 (с использованием уникетола в MATLAB). Эти уникальные наборы затем использовались в качестве отправных точек для задачи оптимизации. Из 981 уникальных начальных точек 359 сходились к набору параметров, который соответствовал всем четырем условиям, и 138 из них были признаны уникальными с допуском 0,05.
Гибкие микросхемы печатных плат – производственные возможности
По мере развития технологий устройства становятся все меньше и требования к скорости передачи данных на печатных платах возрастают.Каждая гибкая печатная плата предназначена для выполнения определенной функции внутри отведенного пространства, но они также должны быть технологичными, иметь строгие механические допуски и быть рентабельными.
Поскольку размеры компонентов с каждым годом становятся все меньше, это позволяет очень маленьким гибким печатным платам включать значительную функциональность, а также быть пассивным компонентом для обеспечения связи в определенных приложениях.
Насколько маленькой может быть гибкая печатная плата?
Размер микросхемы зависит от области применения, но некоторые из ключевых элементов, которые важны для многих клиентов:
- Соблюдение очень жестких допусков по контуру.
- Значения импеданса, которые можно поддерживать на уровне +/- 5% даже в приложениях с большим объемом.
- Позолота для использования в клеммных колодках разъемов.
- Использование паяльной маски LPI на гибкой подложке, чтобы не повредить прокладки паяльной маски.
- Добавление небольших ребер жесткости, чтобы сделать один конец материала жестким. Микросхемы
- требуют очень тонких линий и пространств, что требует специального оборудования для травления.
- Собственные возможности по производству пробивных прессов и оснастки сокращают время выхода на рынок.
Возможности миниатюрной гибкой печатной платы
В следующей таблице представлены общие возможности гибких микросхем. Однако каждое приложение индивидуально, поэтому, пожалуйста, свяжитесь с нами, если у вас возникнут вопросы по дизайну, чтобы мы могли проверить параметры вашего приложения.
| Наименьший размер печатной платы | 5 мм x 5 мм |
| Ширина строки и интервал | 0,002 дюйма (0,05 мм) |
| Толщина меди | 1/4 унции, 1/3 унции, 1/2 унции 1 унция, 2 унции и выше |
| Размер переходного отверстия / сверла | |
| Минимальный диаметр сверла (механическое) | 0.004 “(0,1 мм) |
| Минимальный размер переходного отверстия (лазер) | 2 мил (0,05 мм) |
| Минимальный размер микроперехода (лазер) | 3 мил (0,07 мм) |
| Паяльная маска / покрытие | |
| Паяльная маска Мост между плотиной | 3 мил (0.08 мм) |
| Допуск регистрации паяльной маски | 2 мил (0,05 мм) |
| Регистрация обложки | 8 мил (0,20 мм) |
| Регистрация PIC | 7 мил (0,18 мм) |
| Ребро жесткости | |
| Регистрация ребра жесткости | 8 мил (0.28 мм) |
| Допуск по толщине | 10% |
| Наименьший размер ребра жесткости | 0 мм x 0 мм |
| Импеданс | |
| Импеданс | +/- 8% |
| Стальная линейчатая матрица (SRD)) | |
| Допуск контура | 4 мил (0.1 мм) |
| Минимальный радиус | 4 мил (0,1 мм) |
| Внутренний радиус | 0,51 мм (20 мил) |
| Минимальный размер отверстия пуансона | 28 мил (0,7 мм) |
| Допуск размера перфорационного отверстия | +/- 2 мил (0.051 мм) |
| Ширина слота | 0,51 мм (20 мил) |
| Допуск отверстия до контура | +/- 2 мил (0,05 мм) |
| Допуск края отверстия до контура | +/- 3 мил (0,07 мм) |
| Минимум от трассы до контура | 8 мил (0.20 мм) |
| Электрические испытания | |
| Испытательное напряжение | 50 – 300 В |
| Доступная отделка поверхности | |
| Жесткое золото, ENIG, ENEPIG, OSP, иммерсионное серебро, иммерсионное олово, плакированное олово | |
Приложения для гибких микросхем
Непрерывная миниатюризация устройств привела к тому, что печатные платы стали меньше и плотнее и требовали больше возможностей.Применения, которые мы поставляем гибкие мини-печатные платы, включают:
- Высокоскоростные цифровые / ВЧ / СВЧ-разъемы
- Промышленные датчики
- Расходные материалы для КИП
- Медицинские изделия – носимые устройства
- Слуховые аппараты
- Упаковка для полупроводников
- RFID метки / антенны
- И многое другое…
Epec – ведущий опытный разработчик и производитель миниатюрных печатных плат, уделяющий внимание отраслевым допускам и обслуживанию клиентов. Мы предлагаем полный спектр услуг по проектированию и производству, которые могут быстро адаптироваться к любым вашим требованиям при больших или малых объемах.
Компьютерное моделирование парадигм электроэнцефалографии и функциональной магнитно-резонансной томографии указывает на постоянную потерю синаптического усиления пирамидных клеток при шизофрении
Доступно в Интернете 10 августа 2021 г.
https: // doi.org / 10.1016 / j.biopsych.2021.07.024Получить права и контентAbstract
Предпосылки
Снижение синаптического усиления – чувствительности постсинаптических ответов к нервным сигналам – может быть фундаментальной синаптической патологией при шизофрении. Однако доказательства этого косвенные. Кроме того, неясно, затронуты ли пирамидные клетки или интернейроны (или и то, и другое), и как эти дефициты связаны с симптомами.
Методы
Людям с диагнозом шизофрения (PScz) ( n = 108), их родственникам ( n = 57) и контрольным субъектам ( n = 107) были выполнены 3 парадигмы электроэнцефалографии (ЭЭГ) – в состоянии покоя, Отрицательность рассогласования и установившийся слуховой отклик 40 Гц – и функциональная магнитно-резонансная томография в состоянии покоя.Динамическое причинно-следственное моделирование использовалось для количественной оценки синаптических связей в корковых микросхемах.
Результаты
Были воспроизведены классические групповые различия в характеристиках ЭЭГ между PScz и контрольными субъектами, включая увеличение тета и другие спектральные изменения (ЭЭГ покоя), снижение негативности рассогласования и снижение мощности 40 Гц. Во всех четырех парадигмах характерные особенности данных PScz лучше всего объяснялись моделями с большим самоингибированием (уменьшенным синаптическим усилением) в пирамидных клетках.Кроме того, растормаживание слуховых областей предсказало аномальное слуховое восприятие (и положительные симптомы) при PScz в трех парадигмах.
Выводы
Во-первых, характерные изменения ЭЭГ в PScz в трех классических парадигмах можно отнести к одному и тому же изменению основного параметра: большему самоторможению в пирамидных клетках. Во-вторых, психотические симптомы при PScz связаны с растормаживанием нервных цепей. Эти находки более соизмеримы с гипотезой о том, что при PScz первичная потеря синаптического усиления пирамидных клеток затем компенсируется подавлением интернейронов (скорее, чем наоборот).Они также предполагают, что психотические симптомы связаны с этим вторичным подавлением.
Ключевые слова
Устойчивое слуховое состояние
Динамическая каузальная модель
Отрицательность несоответствия
Психоз
Состояние покоя
Шизофрения
Рекомендуемые статьиЦитирующие статьи (0)
© 2021 Psyological Society of Bichiatry Society.
Рекомендуемые статьи
Ссылки на статьи
Прогнозирование синаптических подключений для моделирования крупномасштабных микросхем с использованием Snudda
Akar, N.A. et al. (2019). Arbor – морфологически детализированная библиотека моделирования нейронных сетей для современных высокопроизводительных вычислительных архитектур. 2019 27-я Международная конференция Euromicro по параллельной, распределенной и сетевой обработке (PDP) , Павия, Италия, 2019 г., стр. 274–282. https://doi.org/10.1109/EMPDP.2019.8671560.
Акрам, М., Нанда, С., Маравер, П., Арманьянсас, Р., и Асколи, Г. А. (2018). Открытое хранилище одноклеточных реконструкций мозгового леса. Sci Data, 5 , 180006. https://doi.org/10.1038/sdata.2018.6.
Артикул PubMed PubMed Central Google ученый
Amunts, K., Knoll, AC, Lippert, T., Pennartz, CMA, Ryvlin, P., Destexhe, A., Jirsa, VK, D’Angelo, E., & Bjaalie, JG (2019) . Проект человеческого мозга – синергия между нейробиологией, вычислениями, информатикой и технологиями, вдохновленными мозгом. PLoS Biology, 17 (7), e3000344.https://doi.org/10.1371/journal.pbio.3000344.
CAS Статья PubMed PubMed Central Google ученый
Анвар Х., Риачи И., Шюрманн Ф. и Маркрам Х. (2009). Подход к регистрации морфологического разнообразия нейронов. В Вычислительная неврология: реалистичное моделирование для экспериментаторов . Де Шуттер Э., редактор. (Кембридж: MIT Press) 211–232. https://doi.org/10.7551/mitpress/
Бертет П., Линдаль М., Талли П. Дж., Хеллгрен-Коталески Дж. И Ланснер А. (2016). Функциональная значимость различных путей базальных ганглиев исследована в модели спайков с пластичностью, зависящей от вознаграждения. Frontiers in Neural Circuits., 10 , 53. https://doi.org/10.3389/fncir.2016.00053.
Артикул PubMed PubMed Central Google ученый
Билле, Ю. Н., Кай, Б., Gratiy, SL, Dai, K., Iyer, R., Gouwens, NW, Abbasi-Asl, R., Jia, X., Siegle, JH, Olsen, SR, Koch, C., Mihalas, S., & Архипов, А. (2020). Систематическая интеграция структурных и функциональных данных в многомасштабные модели первичной зрительной коры мыши. Neuron, 106 (3), 388–403.e18. https://doi.org/10.1016/j.neuron.2020.01.040.
CAS Статья PubMed Google ученый
Браун, К. М., Баррионуево, Г., Канти, А. Дж., Де Паола, В., Хирш, Дж. А., Джефферис, Г. С., Лу, Дж., Снайппе, М., Сугихара, И., и Асколи, Г. А. (2011). Наборы данных DIADEM: репрезентативные световые микроскопические изображения морфологии нейронов для автоматизации цифровых реконструкций. Нейроинформатика, 9 (2–3), 143–157. https://doi.org/10.1007/s12021-010-9095-5.
Артикул PubMed PubMed Central Google ученый
Кэннон, Р. К., Тернер, Д. А., Пьяпали, Г. К., и Уил, Х. В. (1998). Он-лайн архив реконструированных нейронов гиппокампа. Журнал методов неврологии, 84 (1-2), 49-54. https://doi.org/10.1016/s0165-0270(98)00091-0.
CAS Статья PubMed Google ученый
Карневейл, Т., и Хайнс, М. (2006). Книга НЕЙРОН (стр. 2006). Издательство Кембриджского университета. https://doi.org/10.1017/CBO11541612.
Казали, С., Маренци, Э., Медини, К., Казеллато, К., и Д’Анджело, Э. (2019). Реконструкция и моделирование каркасной модели мозжечковой сети. Frontiers in Neuroinformatics, 13 , 37. https://doi.org/10.3389/fninf.2019.00037.
Артикул PubMed PubMed Central Google ученый
Cizeron, M., Qiu, Z., Koniaris, B., Gokhale, R., Komiyama, N.H., Fransén, E., & Grant, S.Г. Н. (2020). Атлас синапсов на протяжении всей жизни мыши. Science., 369 (6501), 270–275. https://doi.org/10.1126/science.aba3163.
CAS Статья PubMed PubMed Central Google ученый
Коланджело, К., Шичкова, П., Келлер, Д., Маркрам, Х., и Рамасвами, С. (2019). Клеточные, синаптические и сетевые эффекты ацетилхолина в неокортексе. Границы в нейронных цепях, 13 , 24.https://doi.org/10.3389/fncir.2019.00024.
CAS Статья PubMed PubMed Central Google ученый
Cuntz, H., Forstner, F., Borst, A., & Häusser, M. (2010). Одно правило, чтобы вырастить их всех: общая теория ветвления нейронов и ее практическое применение. PLoS Computational Biology, 6 (8), e1000877. https://doi.org/10.1371/journal.pcbi.1000877.
CAS Статья PubMed PubMed Central Google ученый
Дай, К., Эрнандо, Дж., Билле, Ю.Н., Грати, С.Л., Планас, Дж., Дэвисон, А.П., Дура-Бернал, С., Глисон, П., Девресс, А., Дихтер, Б.К., Геваерт, М., Кинг , Дж. Г., ван Гейт, ВАХ, Поволоцкий, А. В., Мюллер, Э., Куркол, Дж. Д., и Архипов, А. (2020). Формат данных SONATA для эффективного описания крупномасштабных сетевых моделей. PLoS Computational Biology, 16 (2), e1007696. https://doi.org/10.1371/journal.pcbi.1007696.
CAS Статья PubMed PubMed Central Google ученый
Экономо, М.Н., Виннубст, Дж., Бас, Э., Феррейра, Т. А., и Чандрашекар, Дж. (2019). Реконструкция аксонов одиночных нейронов: поиск схемы соединений мозга. Журнал сравнительной неврологии, 527 (13), 2190–2199. https://doi.org/10.1002/cne.24674.
Артикул PubMed Google ученый
Einevoll, G. T., Destexhe, A., Diesmann, M., Grün, S., Jirsa, V., de Kamps, M., Migliore, M., Ness, T. V., Plesser, H.Э., и Шюрманн Ф. (2019). Научное обоснование моделирования мозга. Neuron., 102 (4), 735–744. https://doi.org/10.1016/j.neuron.2019.03.027.
CAS Статья PubMed Google ученый
Fuzik, J., Zeisel, A., Máté, Z., Calvigioni, D., Yanagawa, Y., Szabó, G., Linnarsson, S., & Harkany, T. (2016). Интеграция электрофизиологических записей с данными секвенирования РНК одиночных клеток позволяет идентифицировать подтипы нейронов. Nature Biotechnology, 34 (2), 175–183. https://doi.org/10.1038/nbt.3443.
CAS Статья PubMed Google ученый
Герфен, К. Р., Экономо, М. Н., и Чандрашекар, Дж. (2018). Проекции кортикальных пирамидных нейронов на большие расстояния. Journal of Neuroscience Research, 96 (9), 1467–1475. https://doi.org/10.1002/jnr.23978.
CAS Статья PubMed Google ученый
Глисон, П., Crook, S., Cannon, RC, Hines, ML, Billings, GO, Farinella, M., Morse, TM, Davison, AP, Ray, S., Bhalla, US, Barnes, SR, Dimitrova, YD, & Silver , РА (2010). NeuroML: язык для описания моделей нейронов и сетей, управляемых данными, с высокой степенью биологической детализации. PLoS Computational Biology, 6 (6), e1000815. https://doi.org/10.1371/journal.pcbi.1000815.
CAS Статья PubMed PubMed Central Google ученый
Гекче, О., Стэнли, Г. М., Трейтлейн, Б., Нефф, Н. Ф., Кэмп, Дж. Г., Маленка, Р. К., Ротвелл, П. Э., Фуччилло, М. В., Судхоф, Т. К., & Quake, С. Р. (2016). Клеточная таксономия полосатого тела мыши по данным одноклеточной RNA-Seq. Cell Reports, 16 (4), 1126–1137. https://doi.org/10.1016/j.celrep.2016.06.059.
CAS Статья PubMed Google ученый
Gratiy, S. L., Billeh, Y. N., Dai, K., Mitelut, C., Feng, D., Гоувенс, Н. В., Каин, Н., Кох, К., Анастасси, К. А., и Архипов, А. (2018). BioNet: интерфейс Python для NEURON для моделирования крупномасштабных сетей. PLoS ONE, 13 (8), e0201630. https://doi.org/10.1371/journal.pone.0201630.
CAS Статья PubMed PubMed Central Google ученый
Грилнер, С., Ип, Н., Кох, К., Корошец, В., Окано, Х., Полачек, М., Пу, М. М., и Сейновски, Т. Дж. (2016).Всемирные инициативы по продвижению исследований мозга. Nature Neuroscience, 19 (9), 1118–1122. https://doi.org/10.1038/nn.4371.
CAS Статья PubMed PubMed Central Google ученый
Герни К., Прескотт Т. Дж. И Редгрейв П. (2001). Вычислительная модель выбора действий в базальных ганглиях. II Анализ и моделирование поведения. Биологическая кибернетика, 84 (6), 411–423. https: // doi.org / 10.1007 / PL00007985.
CAS Статья PubMed Google ученый
Хеллвиг, Б. (2000). Количественный анализ локальной связи между пирамидными нейронами в слоях 2/3 зрительной коры головного мозга крысы. Биологическая кибернетика, 82, (2), 111–121. https://doi.org/10.1007/pl00007964.
CAS Статья PubMed Google ученый
Хепберн, И., Чен, В., Уилс, С., и Де Шуттер, Э. (2012). ШАГИ: Эффективное моделирование стохастических моделей реакции-диффузии в реалистичных морфологиях. BMC Systems Biology, 6 , 36. https://doi.org/10.1186/1752-0509-6-36.
Артикул PubMed PubMed Central Google ученый
Хайнс, М. Л., Дэвисон, А. П., и Мюллер, Э. (2009). НЕЙРОН и Python. Frontiers in Neuroinformatics, 3 , 1. https: // doi.орг / 10.3389 / нейро.11.001.2009.
Артикул PubMed PubMed Central Google ученый
Хьорт, Дж., Блэквелл, К. Т., и Коталески, Дж. Х. (2009). Щелевые соединения между интернейронами полосатого тела с быстрыми импульсами регулируют активность и синхронизацию в зависимости от активности коры. Журнал неврологии, 29 (16), 5276–5286. https://doi.org/10.1523/jneurosci.6031-08.2009.
CAS Статья PubMed PubMed Central Google ученый
Хьорт, Дж.JJ, Kozlov, A., Carannante, I., Frost Nylén, J., Lindroos, R., Johansson, Y., Tokarska, A., Dorst, MC, Suryanarayana, SM, Silberberg, G., Hellgren Kotaleski, J ., & Grillner, S. (2020). Микросхемы полосатого тела in silico. Proceedings of the National Academy of Sciences of the United States of America, 117 (17), 9554–9565. https://doi.org/10.1073/pnas.2000671117.
CAS Статья PubMed PubMed Central Google ученый
Хо, Х., Оба, доктор медицины, Синиард, А., Шарма, С., Нотвелл, Дж. Х., Уоллес, М., Леоне, Д. П., Нгуен, А., Чжао, Э., Ли, Х., Цвиллинг, Д., Томпсон, К.Р., Брейтуэйт, С.П., Хуэнтельман, М., и Портманн, Т. (2018). Руководство по одноклеточной транскриптомике в мозге взрослых грызунов: пересмотр транскриптома среднего шиповатого нейрона. Frontiers in Cellular Neuroscience, 12 , 159. https://doi.org/10.3389/fncel.2018.00159.
CAS Статья PubMed PubMed Central Google ученый
Хамфрис, М.Д., Вуд Р. и Герни К. (2009). Модулируемые дофамином динамические клеточные сборки, генерируемые ГАМКергической стриарной микросхемой. Нейронные сети, 22 (8), 1174–1188. https://doi.org/10.1016/j.neunet.2009.07.018.
Артикул PubMed Google ученый
Инсел, Т. Р., Лэндис, С. К., и Коллинз, Ф. С. (2013). Приоритеты исследований. Инициатива NIH BRAIN. Science., 340 (6133), 687–688. https: // doi.org / 10.1126 / science.1239276.
CAS Статья PubMed PubMed Central Google ученый
Джонс А. Р., Оверли К. С. и Санкин С. М. (2009). Атлас мозга Аллена: 5 лет и более. Обзоры природы. Неврология, 10 (11), 821–828. https://doi.org/10.1038/nrn2722.
CAS Статья PubMed Google ученый
Джордан, Дж., Гелиас, М., Дисманн, М., и Кункель, С. (2020). Эффективная коммуникация в распределенном моделировании нейронных сетей с щелевыми контактами. Frontiers in Neuroinformatics, 14 , 12. https://doi.org/10.3389/fninf.2020.00012.
Артикул PubMed PubMed Central Google ученый
Канари, Л., Рамасвами, С., Ши, Й., Моран, С., Мейстр, Дж., Перин, Р., Абделла, М., Ван, Й., Хесс, К., & Маркрам, Х.(2019). Объективная морфологическая классификация пирамидных клеток неокортекса. Кора головного мозга, 29 (4), 1719–1735. https://doi.org/10.1093/cercor/bhy339.
Артикул PubMed PubMed Central Google ученый
Кумбхар, П., Хайнс, М., Фурио, Дж., Овчаренко, А., Кинг, Дж., Делалондр, Ф., и Шюрманн, Ф. (2019). CoreNEURON: оптимизированный вычислительный движок для симулятора NEURON. Границы нейроинформатики, 13 , 63.https://doi.org/10.3389/fninf.2019.00063.
Артикул PubMed PubMed Central Google ученый
Линдал, М., и Хеллгрен Коталески, Дж. (2017). Распутывание динамики и функции сети базальных ганглиев: роль истощения и ингибирования дофамина, исследованная в модели сети пиков. eNeuro, 3 (6), ENEURO.0156-16.2016. https://doi.org/10.1523/ENEURO.0156-16.2016.
Маркрам, Х., Мюллер, Э., Рамасвами, С., Рейман, М.В., Абделла, М., Санчес, Калифорния, Айламаки, А., Алонсо-Нанкларес, Л., Антилл, Н., Арсевер, С., Каху, GAA, Бергер, Т.К., Бильгили, А., Бунчич, Н., Чалимурда, А., Чиндеми, Г., Куркол, Д.Д., Делалондр, Ф., Делатр, В., Друкманн, С., Дюмуск, Р., Дайнс, J., Eilemann, S., Gal, E., Gevaert, ME, Ghobril, JP, Gidon, A., Graham, JW, Gupta, A., Haenel, V., Hay, E., Heinis, T., Эрнандо, Дж. Б., Хайнс, М., Канари, Л., Келлер, Д., Кеньон, Дж., Хазен, Г., Ким, Ю., Кинг, Дж. Г., Кисвардей, З., Кумбхар, П., Лассер, С., Ле Бе, СП, Магальяйнс, BRC, Мерчан-Перес, А., Мейстр, Дж., Моррис, Б. Р., Мюллер, Дж., Муньос-Сеспедес, А., Муралидхар, С., Мутураса, К., Накбаур, Д., Ньютон, Т.Х., Нольте, М., Овчаренко, А., Паласиос, Дж., Пастор, Л. , Перин, Р., Ранджан, Р., Риачи, И., Родригес, Дж. Р., Таухид, Ф., Телефонт, М., Толедо-Родригес, М., Транклер, Т., Ван Гейт, В., Диас, Дж. В., Уокер, Р., Ван, Ю., Занинетта, С. М., ДеФелипе, Дж., Хилл, С. Л., Сегев, И., & Шюрманн, Ф. (2015). Реконструкция и моделирование микросхем неокортекса. Cell, 163 (2), 456–492. https://doi.org/10.1016/j.cell.2015.09.029.
CAS Статья PubMed Google ученый
Масоли, С., Тоннолина, М., Лафоренца, У., Мочча, Ф., и Д’Анджело, Э. (2020). Настройка параметров позволяет дифференцировать подтипы гранулярных клеток, улучшая свойства передачи на входной стадии мозжечка. Биология коммуникации, 3 (1), 222. https://doi.org/10.1038/s42003-020-0953-x.
CAS Статья PubMed PubMed Central Google ученый
Макдугал, Р. А., Морс, Т. М., Карневале, Т., Маренко, Л., Ван, Р., Мильоре, М., Миллер, П. Л., Шеперд, Г. М., и Хайнс, М. Л. (2017). Двадцать лет ModelDB и далее: создание необходимых инструментов моделирования для будущего нейробиологии. Journal of Computational Neuroscience, 42 (1), 1–10.https://doi.org/10.1007/s10827-016-0623-7.
Артикул PubMed Google ученый
Migliore, R., Lupascu, CA, Bologna, LL, Romani, A., Courcol, J.-D., Antonel, S., van Geit, WAH, Thomson, AM, Mercer, A., Lange , S., Falck, J., Rössert, CA, Shi, Y., Hagens, O., Pezzoli, M., Freund, TF, Kali, S., Muller, EB, Schürmann, F., Markram, H. , & Мильоре, М. (2018). Физиологическая вариабельность плотности каналов в пирамидных клетках и интернейронах гиппокампа CA1 исследована с использованием единого рабочего процесса моделирования на основе данных. PLoS Computational Biology, 14 (9), e1006423. https://doi.org/10.1371/journal.pcbi.1006423.
CAS Статья PubMed PubMed Central Google ученый
Muñoz-Manchado, AB, Bengtsson Gonzales, C., Zeisel, A., Munguba, H., Bekkouche, B., Skene, NG, Lönnerberg, P., Ryge, J., Harris, KD, Linnarsson , S., & Hjerling-Leffler, J. (2018). Разнообразие интернейронов в дорсальном полосатом теле, выявленное с помощью секвенирования одноклеточной РНК и PatchSeq. Cell Rep, 24 (8), 2179–2190.e7. https://doi.org/10.1016/j.celrep.2018.07.053.
CAS Статья PubMed PubMed Central Google ученый
Окано, Х., Мияваки, А., и Касаи, К. (2015). Мозг / РАЗУМ: проект по картированию мозга в Японии. Философские труды Лондонского королевского общества. Серия B, Биологические науки, 370 (1668), 20140310. https://doi.org/10.1098/rstb.2014.0310.
Артикул PubMed PubMed Central Google ученый
Плессер, Х., Дисманн, М., Гевалтиг, М., и Моррисон, А. (2015). Nest: инструмент нейронного моделирования. В D. Jaeger & R. Jung (Eds.), Энциклопедия вычислительной нейробиологии (стр. 1849–1852). Springer Нью-Йорк.
Подляски, В. Ф., Зехольцер, А., Грошнер, Л. Н., Мизенбек, Г., Ранджан, Р., и Фогельс, Т. П. (2017). Картирование функции нейрональных ионных каналов в модели и эксперименте. Elife., 6 , e22152. https://doi.org/10.7554/elife.22152.
Артикул PubMed PubMed Central Google ученый
Ранджан, Р., Хазен, Г., Гамбацци, Л., Рамасвами, С., Хилл, С. Л., Шюрманн, Ф., и Маркрам, Х. (2011). Channelpedia: интегрированная и интерактивная база данных по ионным каналам. Frontiers in Neuroinformatics, 5 , 36. https://doi.org/10.3389/fninf.2011.00036.
Артикул PubMed PubMed Central Google ученый
Рэй С. и Бхалла, США (2008). PyMOOSE: совместимость сценариев на Python для MOOSE. Границы нейроинформатики, 2 , 6.https://doi.org/10.3389/neuro.11.006.2008.
Артикул PubMed PubMed Central Google ученый
Рейманн М.В., Кинг Дж. Г., Мюллер Э. Б., Рамасвами С. и Маркрам Х. (2015). Алгоритм прогнозирования коннектома нейронных микросхем. Frontiers in Computational Neuroscience, 9 , 120. https://doi.org/10.3389/fncom.2015.00120.
Артикул PubMed PubMed Central Google ученый
Розен, Г.Д. и Уильямс Р. В. (2001). Комплексный анализ признаков полосатого тела мыши: независимые QTL модулируют объем и количество нейронов. BMC Neuroscience, 2 , 5. https://doi.org/10.1186/1471-2202-2-5.
CAS Статья PubMed PubMed Central Google ученый
Сантуй, А., Томас-Рока, Л., Родригес, младший, Гонсалес-Сориано, Дж., Чжу, Ф., Цю, З., Грант, SGN, ДеФелипе, Дж. И Мерчан-Перес , А. (2020).Оценка количества синапсов в гиппокампе и в головном мозге с помощью объемной электронной микроскопии и генетической маркировки. Scientific Reports, 10 , 14014. https://doi.org/10.1038/s41598-020-70859-5.
CAS Статья PubMed PubMed Central Google ученый
Сондерс, А., Макоско, Э.З., Высокер, А., Гольдман, М., Кринен, Ф.М., де Ривера, Х., Бьен, Э., Баум, М., Бортолин, Л., Ван, С., Гоева, А., Немеш, Дж., Камитаки, Н., Брамбо, С., Кулп, Д., и МакКэрролл, С. А. (2018). Молекулярное разнообразие и специализация клеток мозга взрослой мыши. Cell, 174 (4), 1015–1030.e16. https://doi.org/10.1016/j.cell.2018.07.028.
CAS Статья PubMed PubMed Central Google ученый
Скорчиони Р., Полаварам С. и Асколи Г. (2008). L-мера: доступный в Интернете инструмент для анализа, сравнения и поиска цифровых реконструкций морфологии нейронов. Протоколы природы, 3 , 866–876. https://doi.org/10.1038/nprot.2008.51.
CAS Статья PubMed PubMed Central Google ученый
Судхакар, С.К., Хонг, С., Райков, И., Публио, Р., Ланг, К., Клоуз, Т., Го, Д., Негрелло, М., и Де Шуттер, Э. ( 2017). Пространственно-временное сетевое кодирование физиологических входов мшистых волокон зернистым слоем мозжечка. PLoS Computational Biology, 13 , e1005754.https://doi.org/10.1371/journal.pcbi.1005754.
CAS Статья PubMed PubMed Central Google ученый
Торбен-Нильсен, Б. (2014). Эффективная и расширяемая библиотека Python для анализа морфологии нейронов. Нейроинформатика, 12 (4), 619–622. https://doi.org/10.1007/s12021-014-9232-7.
Артикул PubMed Google ученый
Ван Гейт, В., Геваерт, М., Чиндеми, Г., Рёссерт, К., Куркол, Дж., Мюллер, Э. Б., Шюрманн, Ф., Сегев, И., и Маркрам, Х. (2016). BluePyOpt: использование программного обеспечения с открытым исходным кодом и облачной инфраструктуры для оптимизации параметров модели в неврологии. Frontiers in Neuroinformatics, 10 , 17. https://doi.org/10.3389/fninf.2016.00017.
Артикул PubMed PubMed Central Google ученый
Wichert, I., Jee, S., De Schutter, E., & Хонг, С. (2020). Pycabnn: эффективное и расширяемое программное обеспечение для создания анатомической основы физиологически реалистичной модели нейронной сети. Frontiers in Neuroinformatics, 14 , 31. https://doi.org/10.3389/fninf.2020.00031.
Артикул PubMed PubMed Central Google ученый
Wilkinson, M. D., Dumontier, M., Aalbersberg, I.J., Appleton, G., Axton, M., Baak, A., Blomberg, N., Boiten, J.В., да Силва Сантос, Л. Б., Борн, П. Е., Боуман, Дж., Брукс, А. Дж., Кларк, Т., Кросас, М., Дилло, И., Думон, О., Эдмундс, С., Эвело, Коннектикут , Финкерс, Р., Гонсалес-Бельтран, А., Грей, AJG, Грот, П., Гобл, К., Грет, Дж. С., Херинга, Дж., ‘Т Хоэн, PAC, Хоофт, Р., Кун, Т. ., Kok, R., Kok, J., Lusher, SJ, Martone, ME, Mons, A., Packer, AL, Persson, B., Rocca-Serra, P., Roos, M., van Schaik, R. ., Sansone, SA, Schultes, E., Sengstag, T., Slater, T., Strawn, G., Swertz, MA, Thompson, M., ван дер Лей, Дж., ван Муллиген, Э., Велтероп, Дж., Ваагмеестер, А., Виттенбург, П., Вольстенкрофт, К., Чжао, Дж., и Монс, Б. (2016). Справедливые руководящие принципы для управления научными данными и рационального использования [опубликованные исправления опубликованы в Sci Data. 2019 марта 19; 6 (1): 6]. Sci Data, 3 , 160018. https://doi.org/10.1038/sdata.2016.18.
Артикул PubMed PubMed Central Google ученый
Уильямс, Т. Дж., Балакришнан, Р., Делалондре, Ф., Шерманн, Ф., Мюллер, Э., и Гевалтиг, М. О. (2018). Крупномасштабное моделирование тканей мозга, Blue Brain Project, EPFL . США: N. p., 2018. Web. https://doi.org/10.2172/1483995.
| #! / Usr / bin / env python | |
| # – * – кодировка: utf-8 – * – | |
| ” | |
| Пример сценария гибридной схемы LFP, применяющий методологию с моделью: | |
| Потянс, Т.и Дисманн, М. “Специфическая кортикальная микросхема клеточного типа: | |
| Связь структуры и активности в полномасштабной модели сети с пиками ». | |
| Cereb. Cortex (2014) 24 (3): 785-806. | |
| DOI: 10.1093 / cercor / bhs358 | |
| Краткое изложение основной процедуры моделирования: | |
| 1.Загрузка набора параметров | |
| а. параметры сети | |
| г. параметры для гибридной схемы | |
| 2. Настроить места назначения файлов для различных выходных данных моделирования | |
| 3. моделирование сети | |
| а. выполнить сетевое моделирование с помощью NEST (www.nest-simulator.org) | |
| г. объединить вывод шипа гнезда из разных рангов MPI | |
| 4. Создайте объектное представление, которое использует sqlite3 для всех выходных сигналов | |
| 5. Итерация по постсинаптическим популяциям: | |
| а. Создать объект Население с соответствующими параметрами для | |
| для каждой конкретной группы | |
| г.Выполните все вычисления для популяций | |
| г. Результат моделирования постобработки всех ячеек в популяции | |
| 6. Постобработка всех выходных данных для конкретных клеток и популяций | |
| 7. Создайте архив для всех неизбыточных выходных данных моделирования | |
| Полная симуляция может быть вызвана вызовом mpirun, например | |
| mpirun -np 64 python example_microcircuit.py | |
| , где число 64 – желаемое количество потоков MPI и ядер ЦП | |
| Учитывая размер сети и потребности в многокамерном LFP- | |
| прогнозов с использованием настоящей схемы, запуск модели в крупном масштабе | |
| Настоятельно рекомендуется использовать вычислительные ресурсы | . |
| ” | |
| из example_plotting import * | |
| импортировать matplotlib.pyplot как plt | |
| из example_microcircuit_params import multicompartment_params, \ | |
| point_neuron_network_params | |
| импорт ОС | |
| , если «ДИСПЛЕЙ» не в ОС.Окружающая среда: | |
| импорт matplotlib | |
| matplotlib.use (‘Agg’) | |
| импортировать numpy как np | |
| из времени импорта времени | |
| import нейрон # NEURON, скомпилированный с MPI, должен быть импортирован до NEST и mpi4py | |
| #, чтобы NEURON не знал о MPI. | |
| import nest # Импорт не используется, но выполняется для обеспечения правильного выполнения | |
| из hybridLFPy import PostProcess, Population, CachedNetwork | |
| из hybridLFPy import setup_file_dest, helpers | |
| от glob import glob | |
| импортный tarfile | |
| импорт lfpykit | |
| из mpi4py импортировать MPI | |
| # установить начальные значения | |
| SEED = 12345678 | |
| SIMULATIONSEED = 12345678 | |
| нп.random.seed (SEED) | |
| ################ Инициализация материалов MPI ########################### | |
| COMM = MPI.COMM_WORLD | |
| РАЗМЕР = COMM.Get_size () | |
| РАНГ = COMM.Get_rank () | |
| # если True, выполнить полную модель.Если False, делать только график. Результаты моделирования | |
| # должен существовать. | |
| rightrun = True | |
| # проверяем, загружен ли файл мода для модели синапса, указанной в expisyn.mod | |
| , если не hasattr (нейрон.ч, ‘ExpSynI’): | |
| , если РАНГ == 0: | |
| os.system (‘nrnivmodl’) | |
| Барьер связи () | |
| нейрон.load_mechanisms (‘.’) | |
| ########################################################################### ######################## | |
| # ПАРАМЕТРЫ | |
| ########################################################################### ######################## | |
| # Полный набор параметров, включая параметры сети | |
| params = multicompartment_params () | |
| ########################################################################### ######################## | |
| # Объявление (я) функции | |
| ########################################################################### ######################### | |
| def merge_gdf (параметры_модели, | |
| raw_label = ‘шипы_’, | |
| file_type = ‘gdf’, | |
| fileprefix = ‘шипы’, | |
| скипроузов = 0): | |
| ” | |
| NEST создает один файл для каждого виртуального процесса, содержащий выходные данные записывающего устройства. | |
| Эта функция собирает и объединяет их в один файл на | |
| жителей сети. | |
| Параметры | |
| ———- | |
| параметры_модели: объект | |
| объект сетевых параметров | |
| raw_label: str | |
| file_type: str | |
| Префикс файла | : str |
| скипроунов: int | |
| Возвращает | |
| ——- | |
| Нет | |
| ” | |
| по умолчанию get_raw_gids (параметры_модели): | |
| ” | |
| Читает текстовый файл, содержащий gid популяций нейронов, созданных в пределах | .|
| симуляция NEST.Эти gid не являются непрерывными, как в модели | .|
| Между ними создается | устройств. |
| Параметры | |
| ———- | |
| параметры_модели: объект | |
| объект сетевых параметров | |
| Возвращает | |
| ——- | |
| gids: список | |
| список идентификаторов и значений нейронов (время всплеска, напряжение и т. Д.)) | |
| ” | |
| gidfile = open (os.path.join (model_params.raw_nest_output_path, | |
| параметры_модели.GID_filename), ‘r’) | |
| gids = [] | |
| для l в гидфайле: | |
| а = л.сплит () | |
| gids.append ([int (a [0]), int (a [1])]) | |
| возврат | |
| # некоторая предварительная обработка | |
| raw_gids = get_raw_gids (model_params) | |
| pop_sizes = [raw_gids [i] [1] – raw_gids [i] [0] + 1 | |
| для i в нп.arange (model_params.Npops)] | |
| raw_first_gids = [raw_gids [i] [0] для i в np.arange (model_params.Npops)] | |
| convert_first_gids = [int (1 + np.sum (pop_sizes [: i])) | |
| для i в np.arange (model_params.Npops)] | |
| для pop_idx в np.аранж (model_params.Npops): | |
| , если pop_idx% SIZE == RANK: | |
| файлов = glob (os.path.join (model_params.raw_nest_output_path, | |
| raw_label + ‘{} *. {}’. Format (pop_idx, | |
| file_type))) | |
| gdf = [] # init | |
| для f в файлах: | |
| new_gdf = помощники.read_gdf (f, skiprows) | |
| для строки в new_gdf: | |
| строка [0] = строка [0] – raw_first_gids [pop_idx] + \ | |
| convert_first_gids [pop_idx] | |
| gdf.append (строка) | |
| принт ( | |
| ‘письмо: {}’.формат ( | |
| os.path.join ( | |
| model_params.spike_output_path, | |
| префикс файла + | |
| ‘_ {}. {}’. Format ( | |
| model_params.X [pop_idx], | |
| file_type)))) | |
| помощников.write_gdf ( | |
| гдф, | |
| os.path.join ( | |
| model_params.spike_output_path, | |
| префикс файла + | |
| ‘_ {}. {}’. Format ( | |
| параметры_модели.X [pop_idx], | |
| file_type))) | |
| Барьер связи () | |
| возврат | |
| по умолчанию dict_of_numpyarray_to_dict_of_list (d): | |
| ” | |
| Преобразовать словарь, содержащий массивы numpy, в словарь, содержащий списки | |
| Параметры | |
| ———- | |
| d: dict | |
| Имя и значение параметра SLI в виде пар словарных ключей и значений | |
| Возвращает | |
| ——- | |
| d: dict | |
| модифицированный словарь | |
| ” | |
| для ключа, значение в d.пунктов (): | |
| if isinstance (значение, dict): # if value == dict | |
| # recurse | |
| d [ключ] = dict_of_numpyarray_to_dict_of_list (значение) | |
| elif isinstance (значение, np.ndarray): # или isinstance (значение, список): | |
| d [ключ] = значение.Толист () | |
| возврат d | |
| по умолчанию send_nest_params_to_sli (p): | |
| ” | |
| Считать параметры и отправить их в SLI | |
| Параметры | |
| ———- | |
| p: dict | |
| Имя и значение параметра SLI в виде пар словарных ключей и значений | |
| Возвращает | |
| ——- | |
| Нет | |
| ” | |
| для имени в списке (стр.ключи ()): | |
| значение = p [имя] | |
| , если isinstance (значение, np.ndarray): | |
| значение = значение.tolist () | |
| , если isinstance (значение, dict): | |
| значение = dict_of_numpyarray_to_dict_of_list (значение) | |
| if name == ‘neuroon_model’: # особый случай, поскольку neon_model – это | |
| # модель NEST, а не строка | |
| попытка: | |
| гнездо.ll_api.sli_run (‘/’ + имя) | |
| nest.ll_api.sli_push (значение) | |
| nest.ll_api.sli_run (‘eval’) | |
| nest.ll_api.sli_run (‘def’) | |
| , кроме BaseException: | |
| print (‘Не удалось поместить переменную% s в стек SLI’% (name)) | |
| печать (тип (значение)) | |
| иначе: | |
| попытка: | |
| гнездо.ll_api.sli_run (‘/’ + имя) | |
| nest.ll_api.sli_push (значение) | |
| nest.ll_api.sli_run (‘def’) | |
| , кроме BaseException: | |
| print (‘Не удалось поместить переменную% s в стек SLI’% (name)) | |
| печать (тип (значение)) | |
| возврат | |
| def slip_run (параметры = объект (), | |
| fname = ‘микросхема.слица, | |
| многословие = ‘M_INFO’): | |
| ” | |
| Принимает в качестве входных данных класс параметров и имя основного скрипта скольжения, инициируя | |
| моделирование. | |
| Параметры | |
| ———- | |
| параметры: объект | |
| экземпляр класса параметров | |
| fname: str | |
| путь к исполняемым SLE-кодам | |
| многословие: ул, | |
| флаг детализации гнезда | |
| Возвращает | |
| ——- | |
| Нет | |
| ” | |
| # Загрузить параметры из файла params и передать их в гнездо | |
| # Python -> SLI | |
| send_nest_params_to_sli (переменные (параметры)) | |
| # установить подробность SLI | |
| гнездо.ll_api.sli_run (“% s setverbosity”% verbosity) | |
| # Запустить моделирование NEST / SLI | |
| nest.ll_api.sli_run (‘(% s) запустить’% fname) | |
| def tar_raw_nest_output (raw_nest_output_path, | |
| delete_files = Верно, | |
| filepatterns = [‘напряжения *.dat ‘, | |
| ‘шипы * .dat’, | |
| ‘weighted_input_spikes * .dat’ | |
| ‘* .gdf’]): | |
| ” | |
| Создать tar-файл с содержимым в raw_nest_output_path и, необязательно, | |
| удалить файлы, соответствующие заданному шаблону. | |
| Параметры | |
| ———- | |
| raw_nest_output_path: путь | |
| params.raw_nest_output_path | |
| delete_files: bool | |
| , если True, удалить.dat файлы | |
| filepatterns: список str | |
| паттернов удаляемых файлов | |
| ” | |
| , если РАНГ == 0: | |
| # создать tarfile | |
| fname = raw_nest_output_path + ‘.tar’ | |
| с tarfile.open (fname, ‘a’) как t: | |
| t.add (raw_nest_output_path) | |
| # удалить файлы из | |
| для выкройки в filepatterns: | |
| для f в glob (os.path.join (raw_nest_output_path, шаблон)): | |
| попытка: | |
| ос.удалить (f) | |
| кроме OSError: | |
| print (‘Ошибка при удалении {}’. Формат (f)) | |
| # sync | |
| Барьер связи () | |
| возврат | |
| ########################################################################### ############################# | |
| # ГЛАВНАЯ процедура моделирования | |
| ########################################################################### ############################# | |
| # tic toc | |
| тик = время () | |
| при правильном запуске: | |
| # установить место назначения файла | |
| setup_file_dest (параметры, clearDestination = True) | |
| ######## Выполните сетевое моделирование ############################################ | |
| при правильном запуске: | |
| # запустить симуляцию гнезда только с параметром точечной нейронной сети | |
| # class | |
| networkParams = point_neuron_network_params () | |
| slo_run (параметры = networkParams, | |
| fname = ‘микросхема.слица, | |
| многословие = ‘M_INFO’) | |
| # предварительная обработка файлов gdf, содержащих пиковые выходные данные, напряжения, взвешенные значения и | |
| # пространственные входные выбросы и токи: | |
| merge_gdf (networkParams, | |
| raw_label = networkParams.spike_recorder_label, | |
| file_type = ‘dat’, | |
| fileprefix = params.networkSimParams [‘метка’], | |
| скипроунов = 3) | |
| # создать архив tar файла папки | |
| # больше не нужен.Удалить | |
| tar_raw_nest_output (params.raw_nest_output_path, delete_files = True) | |
| # Создание объектного представления результатов моделирования, использующего sqlite3 | |
| networkSim = CachedNetwork (** params.networkSimParams) | |
| toc = время () – tic | |
| print (‘Моделирование NEST и обработка файла gdf выполнены в%.3f секунд ‘% toc) | |
| # Настройка измерительных датчиков LFPykit для LFP и CSD | |
| при правильном запуске: | |
| зондов = [] | |
| probes.append (lfpykit.RecExtElectrode (cell = None, ** params.electrodeParams)) | |
| зондов.добавить ( | |
| lfpykit.LaminarCurrentSourceDensity ( | |
| ячейка = Нет, | |
| ** params.CSDParams)) | |
| probes.append (lfpykit.CurrentDipoleMoment (cell = None)) | |
| ####### Настройка популяций ########################################### ######## | |
| при правильном запуске: | |
| # итерация по каждому типу ячеек, запуск моделирования отдельных ячеек и создание | |
| # объект населения | |
| для i, y в перечислении (парам.у): | |
| # создать население: | |
| население = Население ( | |
| # параметры родительского класса | |
| cellParams = params.yCellParams [y], | |
| rand_rot_axis = params.rand_rot_axis [y], | |
| SimulationParams = params.SimulationParams, | |
| PopulationParams = params.populationParams [y], | |
| у = у, | |
| layerBoundaries = params.layerBoundaries, | |
| зонды = зонды, | |
| savelist = params.savelist, | |
| savefolder = params.папка сохранения, | |
| dt_output = params.dt_output, | |
| POPULATIONSEED = SIMULATIONSEED + i, | |
| # дочь класса kwargs | |
| X = параметры X, | |
| networkSim = networkSim, | |
| k_yXL = парам.k_yXL [y], | |
| synParams = params.synParams [y], | |
| synDelayLoc = params.synDelayLoc [y], | |
| synDelayScale = params.synDelayScale [y], | |
| J_yX = params.J_yX [y], | |
| tau_yX = params.tau_yX [y], | |
| recordSingleContribFrac = params.recordSingleContribFrac, | |
| ) | |
| # запустить моделирование популяции и собрать данные | |
| pop.run () | |
| pop.collect_data () | |
| # объект больше не нужен | |
| дель поп | |
| ####### Постобработка вывода моделирования ############################### | |
| # сбросить начальное число, но с этого момента вывод должен быть детерминированным | |
| нп.random.seed (SIMULATIONSEED) | |
| при правильном запуске: | |
| # выполнить некоторую постобработку собранных данных, т.е. суперпозицию | |
| # населения LFP, CSD и т. Д. | |
| postproc = Постпроцесс (y = params.y, | |
| dt_output = параметры.dt_output, | |
| зонды = зонды, | |
| savefolder = params.savefolder, | |
| mapping_Yy = params.mapping_Yy, | |
| savelist = params.savelist | |
| ) | |
| # пройти процедуру | |
| постпроцесс.запустить () | |
| # создать tar-архив с выводом | |
| postproc.create_tar_archive () | |
| # tic toc | |
| print (‘Время выполнения:% .3f секунд’% (time () – tic)) | |
| ########################################################################### ######################### | |
| # Создать набор графиков из результатов моделирования | |
| ########################################################################### ######################### | |
| ########## настройки matplotlib ################################### ###### | |
| PLT.закрыть (‘все’) | |
| , если РАНГ == 0: | |
| # создать сетевой растровый график | |
| x, y = networkSim.get_xy ((500, 1000), дробь = 1) | |
| fig, ax = plt.subplots (1, figsize = (5, 8)) | |
| рис.subplots_adjust (слева = 0,2) | |
| networkSim.plot_raster (ax, (500, 1000), x, y, markersize = 1, marker = ‘o’, | ).|
| альфа = 0,5, легенда = False, pop_names = True) | |
| remove_axis_junk (топор) | |
| ax.set_xlabel (r ‘$ t $ (мс)’, labelpad = 0.1) | |
| топор.set_ylabel (‘популяция’, labelpad = 0.1) | |
| ax.set_title (‘сетевой растр’) | |
| fig.savefig (os.path.join (params.figures_path, ‘network_raster.pdf’), | |
| dpi = 300) | |
| plt.close (рис) | |
| # расположение ячеек графика | |
| fig, ax = plt.подзаголовки (1, 1, figsize = (5, 8)) | |
| fig.subplots_adjust (слева = 0,2) | |
| plot_population (ax, params.’ вместо y в params.y], | |
| цветов = [‘b’, если ‘b’ в y, иначе ‘r’ вместо y в params.y], | |
| слоев = [‘L1’, ‘L2 / 3’, ‘L4’, ‘L5’, ‘L6’], | |
| изометрический угол = np.pi / 24, аспект = ‘равно’) | |
| ax.set_title (‘слои’) | |
| рис.savefig (os.path.join (params.figures_path, ‘Layers.pdf’), dpi = 300) | |
| plt.close (рис) | |
| # расположение ячеек графика | |
| fig, ax = plt.subplots (1, 1, figsize = (5, 8)) | |
| fig.subplots_adjust (слева = 0,2) | |
| plot_population (топор, парам.’for y в params.y], | |
| цветов = [‘b’, если ‘b’ в y, иначе ‘r’ вместо y в params.y], | |
| слоев = [‘L1’, ‘L2 / 3’, ‘L4’, ‘L5’, ‘L6’], | |
| изометрический угол = np.pi / 24, аспект = ‘равно’) | |
| plot_soma_locations (топор, X = params.y, | |
| populations_path = params.’for y в params.y], | |
| цветов = [‘b’, если ‘b’ в y, иначе ‘r’ вместо y в params.y], | |
| изометрический угол = np.pi / 24,) | |
| ax.set_title (‘позиции сомы’) | |
| fig.savefig (os.path.join (params.figures_path, ‘soma_locations.pdf’), | |
| dpi = 150) | |
| PLT.закрыть (рис) | |
| # морфология участков в соответствующих местах | |
| fig, ax = plt.subplots (1, 1, figsize = (5, 8)) | |
| fig.subplots_adjust (слева = 0,2) | |
| plot_population (ax, params.populationParams, params.electrodeParams, | |
| парам.’for y в params.y], | |
| цветов = [‘b’, если ‘b’ в y, иначе ‘r’ вместо y в params.y], | |
| слоев = [‘L1’, ‘L2 / 3’, ‘L4’, ‘L5’, ‘L6’], | |
| изометрический угол = np.pi / 24, аспект = ‘равно’) | |
| plot_morphologies (топор, | |
| X = парам.’for y в params.y], | |
| цветов = [‘b’, если ‘b’ в y, иначе ‘r’ вместо y в params.y], | |
| изометрический угол = np.pi / 24, | |
| populations_path = params.populations_path, | |
| cellParams = params.yCellParams, | |
| дробь = 0.02) | |
| ax.set_title (‘Генераторы LFP’) | |
| fig.savefig (os.path.join (params.figures_path, ‘populations.pdf’), dpi = 300) | |
| plt.close (рис) | |
| # морфология участков в соответствующих местах | |
| fig, ax = plt.подзаголовки (1, 1, figsize = (5, 8)) | |
| fig.subplots_adjust (слева = 0,2) | |
| plot_population (ax, params.populationParams, params.electrodeParams, | |
| params.layerBoundaries, | |
| X = params.’ вместо y в params.y], | |
| цветов = [‘b’, если ‘b’ в y, иначе ‘r’ вместо y в params.y], | |
| слоев = [‘L1’, ‘L2 / 3’, ‘L4’, ‘L5’, ‘L6’], | |
| изометрический угол = np.pi / 24, аспект = ‘равно’) | |
| plot_individual_morphologies ( | |
| топор, | |
| X = парам.’for y в params.y], | |
| цветов = [ | |
| ‘b’, если ‘b’ в y, иначе ‘r’ вместо y в params.y], | |
| изометрический угол = np.pi / 24, | |
| cellParams = params.yCellParams, | |
| PopulationParams = params.populationParams) | |
| топор.set_title (‘морфологии’) | |
| fig.savefig (os.path.join (params.figures_path, ‘cell_models.pdf’), dpi = 300) | |
| plt.close (рис) | |
| # построить составные трассы LFP и CSD | |
| fig = plt.figure (figsize = (13, 8)) | |
| рис.subplots_adjust (слева = 0,075, справа = 0,95, внизу = 0,075, вверху = 0,95, | |
| hspace = 0,2, wspace = 0,2) | |
| gs = gridspec.GridSpec (2, 2) | |
| ax0 = fig.add_subplot (gs [:, 0]) | |
| ax1 = fig.add_subplot (gs [0, 1]) | |
| ax2 = рис.add_subplot (gs [1, 1]) | |
| ax0.set_title (‘сетевой растр’) | |
| ax1.set_title (‘CSD’) | |
| ax2.set_title (‘LFP’) | |
| Т = (500, 700) | |
| x, y = сетьSim.get_xy (T, дробь = 1) | |
| networkSim.plot_raster (ax0, T, x, y, markersize = 1, marker = ‘o’, | ).|
| альфа = 0,5, легенда = False, pop_names = True) | |
| remove_axis_junk (ax0) | |
| ax0.set_xlabel (r ‘$ t $ (мс)’, labelpad = 0.1) | |
| ax0.set_ylabel (‘популяция’, labelpad = 0.1) | |
| plot_signal_sum (ax1, z = params.electrodeParams [‘z’], | |
| fname = os.path.join (params.savefolder, | |
| ‘LaminarCurrentSourceDensity_sum. {- 3} $’, T = T) | |
| ax1.set_xticklabels ([]) | |
| ax1.set_xlabel (”) | |
| plot_signal_sum (ax2, z = params.electrodeParams [‘z’], | |
| fname = os.path.join (params.savefolder, | |
| ‘RecExtElectrode_sum.h5’), | |
| ед. = ‘МВ’, T = T) | |
| ax2.set_xlabel (‘$ t $ (мс)’) | |
| fig.savefig (os.path.join (params.figures_path, ‘complex_signals.pdf’), | |
| dpi = 300) | |
| plt.close (рис) | |
| # построить статистику для текущих дипольных моментов каждой популяции, | |
| # временные следы, | |
| # и прогнозы ЭЭГ на коже черепа с использованием модели 4-сферического объемного проводника | |
| из LFPy импорт FourSphereVolumeConductor | |
| Т = [500, 1000] | |
| P_Y_var = np.zeros ((len (params.Y) + 1, 3)) # дисперсия дипольного момента | |
| для i, Y в перечислении (params.Y): | |
| f = h5py.File ( | |
| os.path.join ( | |
| params.savefolder, | |
| ‘популяции’, | |
| ‘{} _population_CurrentDipoleMoment.h5’.format (Y)), | |
| ‘r’) | |
| srate = f [‘srate’] [()] | |
| P_Y_var [i,:] = f [‘data’] [:, int (T [0] * 1000 / srate):]. Var (axis = -1) | |
| f_sum = h5py.File (os.path.join (params.savefolder, | )|
| ‘CurrentDipoleMoment_sum.h5 ‘),’ r ‘) | |
| P_Y_var [-1,:] = f_sum [‘данные’] [:, int (T [0] * 1000 / srate):]. Var (axis = -1) | |
| tvec = np.arange (f_sum [‘data’]. Shape [-1]) * 1000. / srate | |
| fig = plt.figure (figsize = (5, 8)) | |
| рис.subplots_adjust (слева = 0,2, справа = 0,95, внизу = 0,075, вверху = 0,95, | |
| hspace = 0,4, wspace = 0,2) | |
| ax = fig.add_subplot (3, 2, 1) | |
| ax.plot (P_Y_var, ‘-o’) | |
| ax.legend ([‘$ P_x $’, ‘$ P_y $’, ‘$ P_z $’], fontsize = 8, frameon = False) | |
| топор.2) $ ‘, labelpad = 0) | |
| ax.set_title (‘отклонение сигнала’) | |
| # сделать некоторые прогнозы ЭЭГ | |
радиусов = [79000., 80000., 85000.,.] | |
| сигма = [0,3, 1,5, 0,015, 0,3] | |
г = нп.массив ([[0., 0.,.]]) | |
| rz = np.array ([0., 0., 78000.]) | |
| # рисовать сферические оболочки | |
| ax = fig.add_subplot (3, 2, 2, aspect = ‘equal’) | |
| фи = np.linspace (np.pi / 4, np.pi * 3/4, 61) | |
| для R в радиусах: | |
| х = R * нп.cos (phi) | |
| y = R * np.sin (phi) | |
| осевой график (x, y, lw = 0,5) | |
| ax.plot (0, rz [-1], ‘k.’, Clip_on = False) | |
| ax.plot (0, r [0, -1], ‘k *’, clip_on = False) | |
| ax.axis (‘off’) | |
| топор.легенда ([‘мозг’, ‘CSF’, ‘череп’, ‘скальп’, r ‘$ \ mathbf {P} $’, ‘EEG’], | |
| fontsize = 8, frameon = False) | |
| ax.set_title (‘модель головы с четырьмя сферами’) | |
| сфера_модель = FourSphereVolumeConductor (r, радиусы, сигмы) | |
| # текущий дипольный момент | |
| p = f_sum [‘данные’] [:, int (T [0] * 1000 / скорость): int (T [1] * 1000 / скорость)] | |
| # вычислить потенциал | |
| потенциал = сфера_модель.get_dipole_potential (p, rz) | |
| # график дипольного момента | |
| ax = fig.add_subplot (3, 1, 2) | |
| ax.plot (tvec [(tvec> = T [0]) & (tvec | |
| ax.set_ylabel (r ‘$ \ mathbf {P} (t) $ (nA $ \ mu $ m)’, labelpad = 0) | |
| топор.легенда ([‘$ P_x $’, ‘$ P_y $’, ‘$ P_z $’], fontsize = 8, frameon = True) | |
| ax.set_title (‘текущая сумма дипольного момента’) | |
| # нанесите поверхностный потенциал прямо поверх | |
| ax = fig.add_subplot (3, 1, 3, sharex = ax) | |
| ax.plot (tvec [(tvec> = T [0]) & (tvec | |
| потенциал.T * 1000) # преобразование единиц мВ-> мкВ | |
| ax.set_ylabel (r’EEG ($ \ mu $ V) ‘, labelpad = 0) | |
| ax.set_xlabel (r ‘$ t $ (мс)’, labelpad = 0) | |
| ax.set_title (‘потенциал скальпа’) | |
| fig.savefig ( | |
| ос.path.join ( | )|
| params.figures_path, | |
| ‘current_dipole_moments.pdf’), | |
| dpi = 300) | |
| plt.close (рис) | |
| # добавить цифры в выходной архив .tar | |
| с tarfile.открыть (params.savefolder + ‘.tar’, ‘a:’) как f: | |
| для pdf в glob (os.path.join (params.figures_path, ‘* .pdf’)): | |
| arcname = os.path.join (os.path.split ( | |
| params.savefolder) [- 1], ‘цифры’, os.path.split (pdf) [- 1]) | |
| f.add (name = pdf, arcname = arcname) |
Больше новостей
