
Схема реле на муравей — Bitbucket
———————————————————
>>> СКАЧАТЬ ФАЙЛ <<<
———————————————————
Проверено, вирусов нет!
———————————————————
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
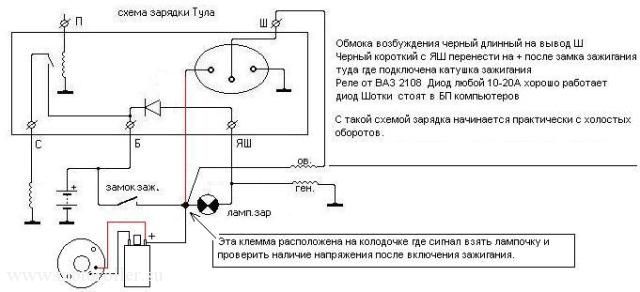
ссылка на схему . Переделка Реле 121 мотороллер Тулица (Часть 2). Innovation. ремонт и проверка динамостартера мотороллер муравей – Duration: 0:37. николай н 8,145 views. →Схема зарядки мотоциклов ТМЗ. pavelmelikov. МУРАВЕЙ) часть 1. Как проверить исправен ли реле регулятор мотоцикла?. В этом видео я расскажу вам о том, как собрать своими руками Регулятор напряжения в сборе с реле стартера, чтобы. →Ссылка на схему зарядки ТМЗ: .
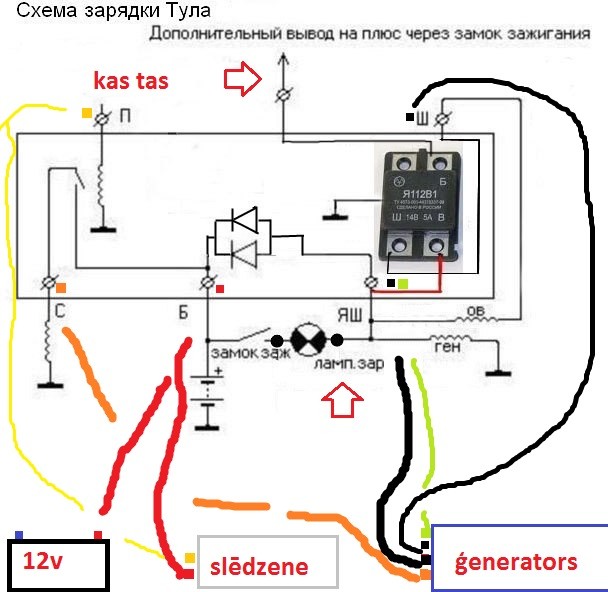 После подключения проводов, и лампочки, как показано на схеме слева ( плюс к плюсу реле, а минус к минусу), и включения зарядного. Имеется генератор 15КВт 400В, нужен реле регулятор. Завтра попробую схему которую предложил Муравей 69 и напишу о. принципиальная электрическая схема мотороллера «Турист» и ее спецификация. Приводим. СПР, Стартерно-пусковое реле, 2 в – 6 в, 1. РОТ, Реле. Переделка реле-регулятора рр121, рр121А на Муравей Тула Тулица Турист. Ремонт двигателей на мотоциклы Муравей Тула ТМЗ Тулица Турист. Муравей . Продам мотороллер Муравей ТМЗ 1982г. выпуска. Мото » Мотоциклы. 6 000 грн. Цылиндер на муравей 1 ремонт. Мотозапчасти и. Систе́ма зажига́ния это совокупность всех приборов и устройств, обеспечивающих. Схемы с электронным зажиганием разделяются на. запуске двигателя добавочное сопротивление шунтируется контактами реле стартера. Если напряжение сразу зашкалило за 15 В, то сгорело реле-регулятор, генератор в. Такая схема работы реле-регулятора принята на всех типах. Магазин.
После подключения проводов, и лампочки, как показано на схеме слева ( плюс к плюсу реле, а минус к минусу), и включения зарядного. Имеется генератор 15КВт 400В, нужен реле регулятор. Завтра попробую схему которую предложил Муравей 69 и напишу о. принципиальная электрическая схема мотороллера «Турист» и ее спецификация. Приводим. СПР, Стартерно-пусковое реле, 2 в – 6 в, 1. РОТ, Реле. Переделка реле-регулятора рр121, рр121А на Муравей Тула Тулица Турист. Ремонт двигателей на мотоциклы Муравей Тула ТМЗ Тулица Турист. Муравей . Продам мотороллер Муравей ТМЗ 1982г. выпуска. Мото » Мотоциклы. 6 000 грн. Цылиндер на муравей 1 ремонт. Мотозапчасти и. Систе́ма зажига́ния это совокупность всех приборов и устройств, обеспечивающих. Схемы с электронным зажиганием разделяются на. запуске двигателя добавочное сопротивление шунтируется контактами реле стартера. Если напряжение сразу зашкалило за 15 В, то сгорело реле-регулятор, генератор в. Такая схема работы реле-регулятора принята на всех типах. Магазин.
Реле регулятор РР-121 мотороллера Муравей (2903.3702) старого образца
Купить Реле регулятор РР-121 мотороллера Муравей (2903.3702) старого образца – (фото, цена, описание, отзывы) Вы можете с доставкой в следущие города Антополь, Барановичи, Барань, Бегомль, Белицк, Белоозерск, Белыничи, Береза, Березино, Березовка, Бешенковичи, Бобр, Бобруйск, Богушевск, Болбасово, Большая Берестовица, Борисов, Боровуха, Браслав, Брест, Буда-Кошелево, Быхов, Василевичи, Верхнедвинск, Ветка, Ветрино, Вилейка, Витебск, Волковыск, Воложин, Вороново, Воропаево, Высокое, Ганцевичи, Глубокое, Глуск, Глуша, Гомель, Горки, Городея, Городище, Городок, Гродно, Давид-Городок, Дзержинск, Дисна, Добруш, Докшицы, Дрибин, Дрогичин, Дубровно, Дятлово, Езерище, Ельск, Жабинка, Желудок, Житковичи, Жлобин, Жодино, Заречье, Заславль, Зеленый Бор, Зельва, Иваново, Ивацевичи, Ивенец, Ивье, Калинковичи, Каменец, Кировск, Клецк, Климовичи, Кличев, Кобрин, Козловщина, Копаткевичи, Копыль, Кореличи, Корма, Коссово, Костюковичи, Коханово, Красная Слобода, Краснополье, Красносельский, Кривичи, Кричев, Круглое, Крупки, Лельчицы, Лепель, Лида, Лиозно, Логишин, Логойск, Лунинец, Любань, Любча, Ляховичи, Малорита, Марьина Горка (Пуховичи), Мачулищи, Микашевичи, Миоры, Мир, Могилев, Мозырь, Молодечно, Мосты, Мстиславль, Наровля, Негорелое, Несвиж, Новогрудок, Новоельня, Новолукомль, Новополоцк, Оболь, Озаричи, Октябрьский, Ореховск, Орша, Осиповичи, Острино, Островец, Ошмяны, Паричи, Петриков, Пинск, Плещеницы, Подсвилье, Полоцк, Порозово, Поставы, Правдинский, Пружаны, Радошковичи, Радунь, Речица, Рогачев, Россь, Руба, Руденск, Ружаны, Светлогорск,Свирь, Свислочь, Сенно, Скидель, Славгород, Слоним, Слуцк, Смиловичи, Смолевичи, Сморгонь, Солигорск, Сопоцкин, Старобин, Старые Дороги, Столбцы, Столин, Стрешин, Сураж, Телеханы, Тереховка, Толочин, Туров, Уваровичи, Узда, Улла, Уречье, Ушачи, Фаниполь, Хойники, Чаусы, Чашники, Червень, Чериков, Чечерск, Шарковщина, Шерешево, Шклов, Шумилино, Щучин, Юратишки и другие.
Модернизация электрооборудования грузового мотороллера Муравей-2
Ремонт династартера мотороллера «муравей»
Эх, сколько-же было в свое время, послано проклятий и не лестных слов «счастливыми» владельцами «Муравьев» в адрес конструкторов, которые нечего лучшего не придумали, кроме как вместо обычного и хорошо зарекомендовавшего себя с лучшей стороны генератора переменного тока, установить весьма несовершенный, сложный, а главное очень ненадежный и капризный «девайс» под названием династартер.
Вот и приходится по сей день владельцам «Муравьев» время от времени брать в руки инструменты и отдуваться за чужие «косяки». Впрочем ладно, сегодня разговор пойдет не об этом…
Типичная ситуация: на «Муравье» при работающем двигателе горит красная лампочка, что нам говорит об отсутствии зарядки, то есть генератор попросту не дает ток. В таких случаях я сперва осматриваю провода идущие к династартеру и реле-регулятору, проверяю работу реле-регулятора и для полного счастья подаю принудительно ток на шунтирующею обмотку, если после всего проделанного зарядка не появилась-значит неисправность скрыта в самом династартере.
Основные неисправности династартера заключаются в следующем:
- Износ или зависание щеток
- Загрязнение коллектора ротора
- Нарушение целостности проводов и обмоток, межвитковое замыкание, замыкание на корпус-все эти неисправности встречается крайне редко в отличии от первых двух
Вот такой у нас сегодня будет экземплярчик.
Для начала нам необходимо снять кожух охлаждения, о том как его снять я подробно останавливаться не буду, так как при его снятии проблем у вас возникнуть не должно.
После снятия кожуха, нам нужно открутить кулачок прерывателя за которым находиться гайка крепящая ротор.
Теперь нужно открутить гайку на цапфе. Берем ключ, в моем случае это головка на 17 и придерживая левой рукой ротор за крыльчатку вентилятора откручиваем гайку (резьба правая).
Ротор династартера снять можно будет только с помощью специального съемника, все ваши попытки снять ротор при помощи монтировок молотков и зубил заранее обречены на провал, так что не советую…
На всякий случай выкладываю размеры съемника.
Затем вкручиваем съемник до упора в ротор, и начинаем закручивать винт. Если таким способом ротор снять не удается, нужно слегка ударять молотком по торцу винта одновременно его закручивая.
Ротор мы благополучно сняли и как вам хорошо видно на фото коллектор его сильно загрязнен (типичная неисправность) что само собой привело к отказу династартера.
После снятия ротора, попутно была выявлена другая неисправность: следы масла в корпусе генератора что свидетельствует о негерметичности сальника или большом радиальном люфте в коренном подшипнике. Сальник в любом случае надо заменить. Для этого, — откручиваем болты на статоре и снимаем его с фланца.
Итак, фланец вместе с сальником снят. Как вам хорошо видно на фото кромки сальника оказались сильно изношены, почему он износился я расскажу дальше.
Как оказалось: причина износа сальника в том, что при установке фланца, герметиком замазали масляный канал (выделен красным) по которому масло поступает к кромкам сальника.
О том как правильно установить фланец с сальником на место я вам расскажу в конце статьи.
А сейчас, мы поговорим о причине которая спровоцировала отказ династартера. Причина оказалась так сказать типичная для генераторов данного типа (постоянного тока) а именно — загрязнение коллектора.
Для того чтобы восстановить его работоспособность нам нужно почистить коллектор ротора. Как это сделать? Все не так сложно как может показаться на первый взгляд, для этого нам понадобиться игла и наждачная бумага зернистостью примерно 400.
Нужно иглой вычистить всю грязь и пыль от щеток между ламелями коллектора, затем зачистить коллектор наждачной бумагой до равномерного блеска. Затем не забудьте тщательно отмыть коллектор от грязи чистым бензином и просушить. Если коллектор сильно изношен его можно проточить на токарном станке (данную операцию можно доверять только хорошему токарю).
После зачистки и отмывки ротора нам необходимо заняться статором, его тоже желательно отмыть чистым бензином и поменять щетки на новые, тут сложного ничего нет главное чтобы щетки ходили в гнездах без заеданий, если щетки туго сидят в гнездах подточите их напильником.
Принципиальная схема
Что касается системы зажигания «Муравья», то ее можно модернизировать самому. Уменьшить ток, проходящий через контакты прерывателя, можно с помощью транзисторного коммутатора ТК-102 (рис.1), который применялся на самых распространенных в прошлом грузовых автомобилях ЗИЛ-130, ГАЗ-53А и др.
Уменьшив таким образом ток в 6-8 раз (до 0,3. ..0,8 А), мы сделаем контакты прерывателя практически вечными. К недостаткам такого решения следует отнести повышенные требования к чистоте контактов, так как масло, грязь и пыль, оказавшиеся между контактами, уже не выгорают, как это было при обычной системе зажигания.
Рис. 1. Схема подключения транзисторного коммутатора ТК-102.
Применение транзисторного коммутатора дает возможность использовать более высоковольтную катушку зажигания Б-114, имеющую большую вторичную обмотку (41500 витков). Поскольку напряжение на свече возрастет с 17 до 25.30 кВ, можно применить свечу с зазором до 1,2 мм, что даст экономию бензина около 30%.
Смотрите также
Комментарии 36
Здравствуйте скажите как зарекомендовал себя двигатель Брайт?Хочу подобный поставить на самоделку но что-то информации по нему в Интернете не нашёл.
Стал тихим (по шуму)и экономичным? А ведь классная идея переоборудовать и двухколесные электроны и туристы
По шуму наверное так же, все зависит от глушителя, у меня он самодельный. Но звук выхлопа приятнее, сразу слышно что мощный двигатель работает, напоминает мощные Японские мотоциклы.
огромное спасибо за ваш опыт…очень пригодиться мне)))
Себе на муравья поставил 125 кубиков 4тактник, пришлось редуктор зеркально собирать, что бы звезда была с другой стороны, переключение передач под родную ножку, выхлоп на него самодельный, теперь можно на муравье тихо подкрастся к человеку и его не слышно, заводится с пол тычка и не паришся со свечками, акум нужен только для запуска с кнопочки, работает, без акума,
как проверить реле регулятор
Как проверить реле регулятор на автомобиле или мотоцикле ? Таким вопросом задаются многие водители, которые обнаружили проблемы с зарядкой аккумуляторной батареи. Реле регулятор — это небольшой, но очень важный прибор, от которого зависит исправная работа и долговечность аккумуляторной батареи. В этой статье мы подробно рассмотрим, как точно убедиться, что реле регулятор вышел из строя, и что для этого понадобится.
Реле регулятор — это небольшой, но очень важный прибор, от которого зависит исправная работа и долговечность аккумуляторной батареи. В этой статье мы подробно рассмотрим, как точно убедиться, что реле регулятор вышел из строя, и что для этого понадобится.Любой исправный реле регулятор поддерживает вырабатываемое генератором напряжение, нужное для зарядки аккумулятора и от исправной работы реле регулятора зависит долговечность аккумуляторной батареи. Ведь реле-регулятор является точным стабилизатором напряжения и не позволяет ему повышаться или понижаться выше положенного, и поддерживает нужное напряжение в независимости от оборотов двигателя (и соответственно ротора генератора) и от количества потребителей электроэнергии.
Кроме этого, реле регулятор постоянно контролирует напряжение на полюсных штырях аккумуляторной батареи и при необходимости включает или отключает напряжение, приходящее на обмотку возбуждения генератора переменного тока.
Для начала следует отметить, что современные интегральные реле регуляторы, при выходе их из строя, ремонту не подлежат, так как имеют неразборный корпус (залит компаундом). Да и вряд ли кто-то будет возиться с ремонтом, даже если реле старого типа и оно разборное. Гораздо проще купить новый реле регулятор, так как стоит он недорого. Но вот прежде чем отправляться в магазин, следует убедиться, что старый реле регулятор действительно неисправен.
Да и вряд ли кто-то будет возиться с ремонтом, даже если реле старого типа и оно разборное. Гораздо проще купить новый реле регулятор, так как стоит он недорого. Но вот прежде чем отправляться в магазин, следует убедиться, что старый реле регулятор действительно неисправен.
В этой статье будет рассмотрена проверка нескольких реле-регуляторов разных типов, и хотя эти приборы бывают и другой маркировки (например на иномарках) но принцип диагностики большинства подобных интегральных реле, практически одинаковый. Проверить зарядку батареи и реле регулятор можно на автомобиле, а можно и после демонтажа его с машины. Рассмотрим оба варианта, а при проверке можно будет воспользоваться лампочкой и простейшим китайским тестером (мультиметром).
Как проверить реле регулятор на автомобиле.
Неисправности реле регулятора обычно подтверждаются систематическим недозарядом (или перезарядом — понижение уровня электролита от выкипания воды) аккумуляторной батареи. А если у вас сгорел реле регулятор и пропала зарядка где нибудь в пути, а ехать домой ещё прилично, то чтобы не разрядить батарею, можно пока воспользоваться обычной лампочкой, которая заменит реле регулятор в пути (подробнее об этом читаем вот в этой статье).
Для простейшей проверки нам понадобится тестер, выставленный в режим вольтметра, на замер постоянного тока в пределах от 0 до 20 вольт.
Щупы тестера подключаем к полюсным штырям батареи и прежде чем заводить двигатель, запоминаем какое напряжение батареи показал вольтметр (12 — 12,8 вольт — зависит от состояния аккумулятора). Далее заводим двигатель и смотрим на показания вольтметра — напряжение на полюсных штырях батареи, после пуска двигателя должно повыситься (примерно 13 — 13,8 вольт — зависит от оборотов двигателя).
При повышении оборотов двигателя, напряжение тоже должно повышаться, например на средних оборотах оно примерно 13, 5 — 14 вольт, а на максимальных 14 — 14,5 вольт. Если после пуска двигателя, напряжение не повысилось (осталось таким, какое было до запуска мотора, примерно 12 — 12,8 вольт) то скорей всего реле регулятор неисправен.
Конечно же зарядки батареи может не быть и по другим причинам, например от неисправностей генератора, но чтобы точно убедиться, что реле неисправно, его следует снять с машины для более точной проверки, которая будет описана ниже.
Проверка реле регулятора снятого с машины.
Если кто то из начинающих водителей пока не знает, как снять реле регулятор с автомобиля (и заменить его) то подробно об этом читаем и смотрим вот здесь. Там же описано и про дифектовку и замену щёток генератора (ведь щётки современных генераторов объединены в один узел с интегральным реле регулятором).
Для начала опишу проверку интегрального реле регулятора более современного типа (например Я212А, подобные реле стоят не только на современных отечественных машинах, но и на многих иномарках), который объединён в один узел вместе с щёткодержателем (см. фото слева).
А раз такой реле регулятор — это и щёточный узел, то разумеется он крепится прямо на генераторе сзади, в районе контактных колец якоря генератора. Открутив два винта, удерживающих реле регулятор и сняв клеммы с него, извлекаем его из окна крышки генератора, сдуваем угольную пыль и подключаем к несложной схемке, показанной на рисунке слева.
Для проверки понадобится опять же тестер, выставленный в режим вольтметра и 12-ти вольтовая лампочка с патрончиком (например от поворотника), несколько проводов с клеммами. Так же потребуется блок питания (от 0 до 20 вольт) или зарядное устройство с регулировкой тока.
А так как некоторые зарядные устройства не являются блоками питания (то есть не вырабатывают постоянный ток без нагрузки — без подключенной батареи) то к зарядному устройству придётся подключить нагрузку — то есть подключить аккумулятор (так же как на зарядку) и уже от аккумулятора подключить провода, как на простейшей электро схеме чуть ниже.
После подключения проводов, и лампочки, как показано на схеме слева (плюс к плюсу реле, а минус к минусу), и включения зарядного устройства, лампочка загорится и после этого начинаем повышать напряжение регулятором тока на зарядном устройстве и наблюдаем за показаниями вольтметра и лампочкой (у кого нет встроенного вольтметра в зарядном устройстве, то подключаем щупы тестера, выставленного в режим вольтметра, к полюсным штырям аккумулятора).
Лампа должна гореть до показаний вольтметра вплоть до 14,5 вольт, но как только напряжение превысит 14,5 вольт (при повышении напряжения регулятором зарядного устройства), лампа должна погаснуть. Если напряжение попробовать уменьшить (меньше 14,5 вольт) то лампочка опять должна загореться.
Если не происходит всё то, что описано выше, то значит реле неисправно. Если же лапочка гаснет при достижении напряжения более 14,5 вольт (например при 14,6 — 15 вольт и выше) то такое реле тоже следует заменить, так как с ним будет постоянный перезаряд батареи, что приведёт к постоянному выкипанию воды из электролита (придётся постоянно доливать) и это ощутимо снизит ресурс батареи.
В итоге, если сэкономить немного денег на новое реле, потом придётся тратить в десятки раз больше, на новую батарею.
Аналогично проверяем интегральные реле регуляторы Я112В и им подобные так называемые «шоколадки», которые стоят на более старых отечественных машинах.
Реле подключаем для проверки, как показано на рисунке слева и также следим за показаниями вольтметра и лампочки, которая должна погаснуть при достижении напряжения на полюсных штырях батареи более 14,5 вольт.
Ну и ещё одна схема, которая позволяет проверить реле регулятор самого старого типа (маркировка 591,3702-01) , который устанавливается не на генераторе, а отдельно на кузове вазовской классики (такие же реле регуляторы многие ставят и на отечественные тяжёлые мотоциклы).
Кстати такое выносное реле регулятор удобно не только тем, что до него легко добраться, но ещё и тем, что если даже реле сгорит где то в пути, всегда можно добраться домой не разрядив аккумулятор. Для этого к проводам, которые приходят на контакты реле 67 и 15, подключаем 12-ти вольтовую лампочку, и можно ехать дальше, не опасаясь, что батарея разрядится.
Ну, а если подключить лампочку вместо такого реле регулятора (к тем проводам, которые шли на контакты 15 и 67) и снять клемму с полюсного штыря батареи, и при этом мотор не заглохнет, значит генератор исправен.
И последнее, что хотелось бы написать. Часто виновником проблем с зарядкой батареи являются окисленные контакты (клеммы) реле регулятора (особенно контакты под маркировкой БВ, Б3, В, 61, 15 — маркировка зависит от типа реле) или проводов и их клемм, приходящих к реле, замку зажигания, генератору. Ведь окислы, грязь на клеммах, являются причиной возникновения переходного сопротивления и потери напряжения.
Иногда на слишком запущенных машинах, вместо напряжения 12,5 вольт на окисленных клеммах, вольтметром можно отметить лишь жалкие 10 — 11 вольт. Но стоит привести контакты в порядок (зачистить до блеска), и всё приходит в норму. Следует отметить, что прежде чем проверять реле, как было описано выше, проверьте контакты реле и не только реле (и по необходимости приведите их в порядок), но как было сказано выше, особенно те контакты реле, маркировка которых описана выше.
Напряжение на контактах должно быть точно такое же как на полюсных штырях вашей аккумуляторной батареи (без потерь). Если же напряжение на контактах меньше, то следует обязательно их зачистить и возможно реле регулятор вполне исправен и причина была только лишь в окислах. После зачистки контактов, чтобы проблемы не повторились, их следует опрыснуть специальным спреем для контактов (например таким, как в этой статье), который надолго предотвратит окисление.
Если же напряжение на контактах меньше, то следует обязательно их зачистить и возможно реле регулятор вполне исправен и причина была только лишь в окислах. После зачистки контактов, чтобы проблемы не повторились, их следует опрыснуть специальным спреем для контактов (например таким, как в этой статье), который надолго предотвратит окисление.
Ну вот вроде бы и всё. Надеюсь многие новички теперь знают, как проверить реле регулятор самостоятельно, не обращаясь к услугам авто-электрика, успехов всем.
Муравьи, следующие по маршруту, реагируют на изменение последовательности просмотра
. 2020 15 июля; 223 (Пт 14): jeb218701. DOI: 10.1242 / jeb.218701.Принадлежности Расширять
Принадлежности
- 1 Центр исследований в области познания животных, CNRS, Университет Поля Сабатье, Тулуза, 31062 Cedex 09, France sebastian.
 [email protected].
[email protected]. - 2 Sheffield Robotics, Департамент компьютерных наук, Университет Шеффилда, Вестерн Банк, Шеффилд S10 2TN, Великобритания.
- 3 Школа информатики Эдинбургского университета, Крайтон-стрит, Эдинбург EH8 9AB, Великобритания.
- 4 Центр исследований в области познания животных, CNRS, Университет Поля Сабатье, Тулуза, 31062 Cedex 09, Франция.
Элемент в буфере обмена
Себастьян Шварц и др.
J Exp Biol. .
Показать варианты
Формат АннотацияPubMedPMID
.2020 15 июля; 223 (Пт 14): jeb218701. DOI: 10.1242 / jeb.218701.Принадлежности
- 1 Центр исследований в области познания животных, CNRS, Университет Поля Сабатье, Тулуза, 31062 Cedex 09, Франция sebastian.schwarz@univ-tlse3.фр.
- 2 Sheffield Robotics, Департамент компьютерных наук, Университет Шеффилда, Вестерн Банк, Шеффилд S10 2TN, Великобритания.

- 3 Школа информатики Эдинбургского университета, Крайтон-стрит, Эдинбург EH8 9AB, Великобритания.
- 4 Центр исследований в области познания животных, CNRS, Университет Поля Сабатье, Тулуза, 31062 Cedex 09, Франция.
Элемент в буфере обмена
Полнотекстовые ссылки Опции CiteDisplayПоказать варианты
Формат АннотацияPubMedPMID
Абстрактный
Муравьи могут перемещаться, сравнивая текущее восприятие вида с запомненными видами на знакомом маршруте поиска пищи.Модели, касающиеся следования по маршруту, предполагают, что представления сохраняются и вызываются из памяти независимо от последовательности, в которой они появляются. Следовательно, муравью нужно только оценить мгновенное знакомство с текущим видом, чтобы получить направление движения. Это исследование исследует, влияет ли на самонаведение муравья изменения в последовательности взглядов на знакомом маршруте, используя частоту остановок и сканирований в качестве индикатора неопределенности навигации муравья. Муравьев приучили искать корм между своим гнездом и кормушкой, из которой они выходили через короткий канал, прежде чем отправиться домой.В ходе испытаний муравьев собирали перед входом в гнездо и снова выпускали в канал, который помещали либо в исходное положение, либо на полпути по маршруту. Таким образом, муравьи, покидающие знакомый канал в середине маршрута, увидят знакомые виды в новой последовательности. Результаты показывают, что муравьи, выходящие из канала, значительно больше сканируют, когда оказываются в середине маршрута, по сравнению с муравьями, появляющимися в ожидаемом месте рядом с кормушкой. Такое поведение предполагает, что ранее встречавшиеся представления влияют на распознавание текущих представлений, даже если эти представления хорошо знакомы, открывая компонент последовательности для маршрутизации памяти.
Следовательно, муравью нужно только оценить мгновенное знакомство с текущим видом, чтобы получить направление движения. Это исследование исследует, влияет ли на самонаведение муравья изменения в последовательности взглядов на знакомом маршруте, используя частоту остановок и сканирований в качестве индикатора неопределенности навигации муравья. Муравьев приучили искать корм между своим гнездом и кормушкой, из которой они выходили через короткий канал, прежде чем отправиться домой.В ходе испытаний муравьев собирали перед входом в гнездо и снова выпускали в канал, который помещали либо в исходное положение, либо на полпути по маршруту. Таким образом, муравьи, покидающие знакомый канал в середине маршрута, увидят знакомые виды в новой последовательности. Результаты показывают, что муравьи, выходящие из канала, значительно больше сканируют, когда оказываются в середине маршрута, по сравнению с муравьями, появляющимися в ожидаемом месте рядом с кормушкой. Такое поведение предполагает, что ранее встречавшиеся представления влияют на распознавание текущих представлений, даже если эти представления хорошо знакомы, открывая компонент последовательности для маршрутизации памяти.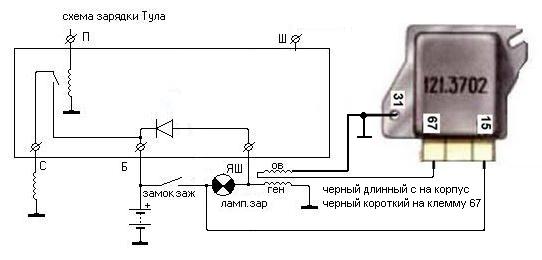 Обсуждается, как информация о последовательностях просмотра может быть реализована в мозгу насекомого, а также возможные альтернативные объяснения нашим результатам.
Обсуждается, как информация о последовательностях просмотра может быть реализована в мозгу насекомого, а также возможные альтернативные объяснения нашим результатам.
Ключевые слова: Катаглифис; Муравьи пустыни; Самонаведение; Обучение насекомых; Память насекомых; Навигация.
© 2020. Издатель ООО «Компания Биологов».
Заявление о конфликте интересов
Конкурирующие интересы Авторы заявляют об отсутствии конкурирующих или финансовых интересов.
Похожие статьи
- Воспоминания пустынного муравья о недавнем визуальном опыте и контроль над маршрутом.
Коллетт М. Коллетт М.Proc Biol Sci. 2014 22 июля; 281 (1787): 20140634. DOI: 10.1098 / rspb.2014.0634. Proc Biol Sci. 2014 г. PMID: 24870046 Бесплатная статья PMC.
- Интеграция путей контролирует следование за гнездом и шлейфом у пустынных муравьев.
Бюльманн К., Ханссон Б.С., Кнаден М. Buehlmann C, et al. Curr Biol. 2012 10 апреля; 22 (7): 645-9. DOI: 10.1016 / j.cub.2012.02.029. Epub 2012 8 марта. Curr Biol.2012 г. PMID: 22405868
- Оценка пройденного муравьями расстояния: эксперименты с пустынными муравьями Cataglyphis fortis.
Зоммер С., Венер Р. Sommer S, et al. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2004 Янв; 190 (1): 1-6. DOI: 10.1007 / s00359-003-0465-4.
 Epub 2003 12 ноября.
J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2004 г.
PMID: 14614570
Epub 2003 12 ноября.
J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2004 г.
PMID: 14614570 - Навигация по насекомым: живут ли муравьи сейчас?
Грэм П., Манган М.Грэм П. и др. J Exp Biol. 2015 Март; 218 (Pt 6): 819-23. DOI: 10.1242 / jeb.065409. J Exp Biol. 2015 г. PMID: 25788724 Рассмотрение.
- Просто следите за своим носом: муравьи ориентируются по обонятельным сигналам.
Штек К. Штек К. Curr Opin Neurobiol. 2012 Апрель; 22 (2): 231-5. DOI: 10.1016 / j.conb.2011.10.011. Epub 2011 1 декабря. Curr Opin Neurobiol. 2012 г. PMID: 22137100 Рассмотрение.
Типы публикаций
- Поддержка исследований, за пределами США.
 Правительство
Правительство
LinkOut – дополнительные ресурсы
Полнотекстовые источники
Материалы исследований
цитировать
КопироватьФормат: AMA APA ГНД NLM
границ | Изучение четких химических меток на сокамерниках у муравьев
Введение
У эусоциальных насекомых способность различать членов из собственной колонии (сокамерников) и членов из чужой колонии (не спаренных) имеет фундаментальное значение для когерентности колонии и, в конечном итоге, способствует приспособленности колонии (Hölldobler and Wilson, 1990).Дискриминация между сокамерниками и не-соклестницами предотвращает использование ресурсов колонии (например, хранилища пищи, выводка) конкурирующими колониями особого и гетероспецифического вида, хищниками и паразитами. У муравьев различие между сокамерниками и не-гнездовыми товарищами основывается на обонянии и на смесях низколетучих химических компонентов в экзоскелете муравьев (кутикулярные углеводороды: CHCs; Lahav et al., 1999). Основные функции КГК – предотвращение высыхания (Lockey, 1988) и защита от инфекций.Во-вторых, CHC использовались как сигналы признания членства в колонии. Состав КГК зависит от вида (Martin S. and Drijfhout F., 2009) и определяется генетически, но профили КГС отдельных особей дополнительно формируются диетой и материалом гнезд (Jutsum et al., 1979; Vander Meer et al., 1989). Частый обмен CHC между сокамерниками посредством трофалаксиса и аллогруминга (Lenoir et al., 2001) приводит к однородной, специфичной для колонии химической сигнатуре (Crozier and Dix, 1979). Эта сигнатура обычно называется запахом колонии и состоит из всех профилей CHC, присущих особям одной колонии.Запахи соседних, созидательных колоний, как правило, различаются только количественным соотношением ХГК.
У муравьев различие между сокамерниками и не-гнездовыми товарищами основывается на обонянии и на смесях низколетучих химических компонентов в экзоскелете муравьев (кутикулярные углеводороды: CHCs; Lahav et al., 1999). Основные функции КГК – предотвращение высыхания (Lockey, 1988) и защита от инфекций.Во-вторых, CHC использовались как сигналы признания членства в колонии. Состав КГК зависит от вида (Martin S. and Drijfhout F., 2009) и определяется генетически, но профили КГС отдельных особей дополнительно формируются диетой и материалом гнезд (Jutsum et al., 1979; Vander Meer et al., 1989). Частый обмен CHC между сокамерниками посредством трофалаксиса и аллогруминга (Lenoir et al., 2001) приводит к однородной, специфичной для колонии химической сигнатуре (Crozier and Dix, 1979). Эта сигнатура обычно называется запахом колонии и состоит из всех профилей CHC, присущих особям одной колонии.Запахи соседних, созидательных колоний, как правило, различаются только количественным соотношением ХГК. Поскольку рацион, материал гнезд и состав колонии могут меняться, запах колонии не постоянен, а меняется со временем (Vander Meer et al., 1989). Важно отметить, что профили CHC среди отдельных рабочих колонии не так однородны, как предполагалось ранее. У рабочих есть профили CHC для конкретных задач, потому что выполнение заданий, например, строительство гнезда, уход за выводком или поиск пищи за пределами гнезда, влияет на профили CHC (Wagner et al., 1998; Kaib et al., 2000). Более того, смешивание КГК между отдельными лицами в колонии не является полным, потому что рабочие в рамках целевой группы сталкиваются и взаимодействуют друг с другом чаще, чем между рабочими группами (Sendova-Franks and Franks, 1995; Mersch et al., 2013; Pamminger et al., 2014; Tschinkel, Hanley, 2017). Основываясь на этих систематических различиях в профилях CHC, рабочие могут различать сокамерников, выполняющих разные задачи (Bonavita-Cougourdan et al., 1993; Greene and Gordon, 2003).
Поскольку рацион, материал гнезд и состав колонии могут меняться, запах колонии не постоянен, а меняется со временем (Vander Meer et al., 1989). Важно отметить, что профили CHC среди отдельных рабочих колонии не так однородны, как предполагалось ранее. У рабочих есть профили CHC для конкретных задач, потому что выполнение заданий, например, строительство гнезда, уход за выводком или поиск пищи за пределами гнезда, влияет на профили CHC (Wagner et al., 1998; Kaib et al., 2000). Более того, смешивание КГК между отдельными лицами в колонии не является полным, потому что рабочие в рамках целевой группы сталкиваются и взаимодействуют друг с другом чаще, чем между рабочими группами (Sendova-Franks and Franks, 1995; Mersch et al., 2013; Pamminger et al., 2014; Tschinkel, Hanley, 2017). Основываясь на этих систематических различиях в профилях CHC, рабочие могут различать сокамерников, выполняющих разные задачи (Bonavita-Cougourdan et al., 1993; Greene and Gordon, 2003).
Хотя профили CHC различаются в пределах колонии, агрессия среди сокамерников редка, и их быстро и с высокой надежностью отличить от одноклассников, не являющихся одноклассниками, даже несмотря на то, что однотипные негнездовые товарищи могут иметь профили CHC, которые качественно идентичны и различаются только соотношением компонентов ( Строймейт и др. , 2010). Неизвестно, как рабочие различают сокамерников и других на основе соотношения CHC. Чтобы понять нейронные и поведенческие механизмы распознавания сородичей, нам необходимо изучить все части системы распознавания, от профилей CHC до поведенческих реакций.Это включает в себя анализ того, как информация, содержащаяся в профиле CHC, обнаруживается и обрабатывается нервной системой, как «соседи по гнезду» и «не сожитель» закодированы в мозгу, как это влияет на принятие решений и, в конечном итоге, приводит к наблюдаемым поведенческим результатам во время встречи.
, 2010). Неизвестно, как рабочие различают сокамерников и других на основе соотношения CHC. Чтобы понять нейронные и поведенческие механизмы распознавания сородичей, нам необходимо изучить все части системы распознавания, от профилей CHC до поведенческих реакций.Это включает в себя анализ того, как информация, содержащаяся в профиле CHC, обнаруживается и обрабатывается нервной системой, как «соседи по гнезду» и «не сожитель» закодированы в мозгу, как это влияет на принятие решений и, в конечном итоге, приводит к наблюдаемым поведенческим результатам во время встречи.
Были предложены различные модели и возможные стратегии, позволяющие различать сородичей и не сородичей. Основываясь на модели соответствия фенотипа (Holmes and Sherman, 1983), каждый человек несет как признаки распознавания своего собственного профиля CHC (метка), так и нейронное представление (шаблон) своего собственного запаха колонии (Lacy and Sherman, 1983).Теоретически метка от встретившегося человека сравнивается с «собственным» шаблоном, что обозначается как процесс сопоставления метки с шаблоном. В самой простой форме предполагается, что люди аносмичны по отношению к запаху собственной колонии, и поэтому ярлыки, специфичные для колонии, не воспринимаются, а все ярлыки, вызывающие заметное восприятие, считаются принадлежащими не-гнездовым товарищам. Хотя эта идея широко предлагалась (Ozaki et al., 2005), она не объясняет, как рабочие могут различать сокамерников из разных рабочих групп (Bonavita-Cougourdan et al., 1993; Greene and Gordon, 2003) и последующие исследования показали, что рабочие не аносмичны по отношению к запаху собственной колонии (Brandstaetter and Kleineidam, 2011; Brandstaetter et al., 2011; Sharma et al., 2015).
В самой простой форме предполагается, что люди аносмичны по отношению к запаху собственной колонии, и поэтому ярлыки, специфичные для колонии, не воспринимаются, а все ярлыки, вызывающие заметное восприятие, считаются принадлежащими не-гнездовым товарищам. Хотя эта идея широко предлагалась (Ozaki et al., 2005), она не объясняет, как рабочие могут различать сокамерников из разных рабочих групп (Bonavita-Cougourdan et al., 1993; Greene and Gordon, 2003) и последующие исследования показали, что рабочие не аносмичны по отношению к запаху собственной колонии (Brandstaetter and Kleineidam, 2011; Brandstaetter et al., 2011; Sharma et al., 2015).
Наиболее широко распространенная модель распознавания сокамерников рассматривает часто используемые ярлыки товарищей по гнезду в раннем взрослом возрасте как поучительные для формирования шаблона (Carlin and Hölldobler, 1983; Morel and Blum, 1988; Errard, 1994). Этот процесс обучения позволяет людям распознавать сокамерников и различать «мы» и«Другие». Поскольку запах колонии может меняться со временем, людям необходимо соответствующим образом скорректировать свой шаблон. Манипулирование запахом колонии приводит к принятию как текущего запаха колонии, так и запаха предыдущей колонии, и для одного шаблона это потребует расширения диапазона приемлемости (Leonhardt et al., 2007; Guerrieri et al., 2009). ; Bos, d’Ettorre, 2012; Bos, 2014).
Поскольку запах колонии может меняться со временем, людям необходимо соответствующим образом скорректировать свой шаблон. Манипулирование запахом колонии приводит к принятию как текущего запаха колонии, так и запаха предыдущей колонии, и для одного шаблона это потребует расширения диапазона приемлемости (Leonhardt et al., 2007; Guerrieri et al., 2009). ; Bos, d’Ettorre, 2012; Bos, 2014).
Поскольку для распознавания товарищей по гнезду можно использовать различные метки (Fielde, 1903; Errard, 1994), вызывает недоумение, как рабочие могут по-прежнему достичь высокой надежности в распознавании товарищей по гнезду, используя один шаблон.Диапазон специфичности этого нейронного шаблона должен быть достаточно широким, чтобы охватывать множество различных меток, присутствующих в колонии, и в то же время исключать метки не-гнездовых товарищей (Reeve, 1989).
В качестве альтернативы единому унифицирующему шаблону мы предполагаем, что рабочие используют несколько шаблонов, каждый со своим собственным диапазоном специфичности, для распознавания соседей по гнезду. В этом случае химически разные этикетки распознаются с помощью разных шаблонов, которые, возможно, снабжены дополнительной информацией о носителе этикеток.Метки, идентифицированные из различных шаблонов, могут восприниматься отдельно или обобщаться как все принадлежащие «товарищам по гнезду». Такая система распознавания с множеством шаблонов сравнима с ассоциативным обучением, аналогичным тому, как пчелы изучают связь между цветами и цветочными ароматизаторами, когда цветы обеспечивают видоспецифический химический образец, и можно изучить множество различных типов профилей цветочных запахов (Bitterman et al. , 1983; Гербер и др., 1996). Разница между этими двумя гипотезами распознавания сородичей заключается в том, что в системе распознавания сородичей с одним шаблоном общие черты профилей CHC запаха колонии используются для распознавания сородичей, тогда как в системе распознавания сородичей с множеством шаблонов химически различные профили CHC внутри колонии распознаются независимо друг от друга и классифицируются как принадлежащие к товарищам по гнезду.
В этом случае химически разные этикетки распознаются с помощью разных шаблонов, которые, возможно, снабжены дополнительной информацией о носителе этикеток.Метки, идентифицированные из различных шаблонов, могут восприниматься отдельно или обобщаться как все принадлежащие «товарищам по гнезду». Такая система распознавания с множеством шаблонов сравнима с ассоциативным обучением, аналогичным тому, как пчелы изучают связь между цветами и цветочными ароматизаторами, когда цветы обеспечивают видоспецифический химический образец, и можно изучить множество различных типов профилей цветочных запахов (Bitterman et al. , 1983; Гербер и др., 1996). Разница между этими двумя гипотезами распознавания сородичей заключается в том, что в системе распознавания сородичей с одним шаблоном общие черты профилей CHC запаха колонии используются для распознавания сородичей, тогда как в системе распознавания сородичей с множеством шаблонов химически различные профили CHC внутри колонии распознаются независимо друг от друга и классифицируются как принадлежащие к товарищам по гнезду.
Чтобы исследовать эти два возможных механизма распознавания товарищей по гнезду, мы систематически манипулировали профилями CHC рабочих, происходящих из одной и той же колонии, не добавляя ни одного, одного или двух углеводородов, и проверяли, принимают ли они друг друга как товарищей по гнезду. Основываясь на системе распознавания соплеменников с одним шаблоном, мы прогнозируем, что рабочие, которые включили два дополнительных углеводорода в свой профиль CHC, также примут людей, у которых добавлен только один углеводород (рис. 1). Напротив, если рабочие с двумя дополнительными углеводородами не принимают людей, у которых добавлен только один из углеводородов, поддерживается идея системы распознавания соплеменников с несколькими шаблонами.
Рисунок 1 . Схематические диапазоны приемлемости в виде овалов (пунктирные линии) для одного шаблона и в виде кружков (сплошные линии) для системы распознавания соплеменников с несколькими шаблонами (0: без дополнительных компонентов, A: один дополнительный, AB: два дополнительных компонента, добавленных к кутикуле. углеводород, профиль CHC).
углеводород, профиль CHC).
Материалы и методы
Для всех экспериментов мы использовали взрослых рабочих вида Camponotus floridanus (Buckley, 1866) из зрелой колонии, собранной А.Эндлер в июле 2003 г. Сбор маток-основателей для лабораторных колоний соответствовал законам Соединенных Штатов Америки, действующим на момент сбора. В колониях этого вида спаривается одна матка (моногинная), которая спаривается только с одним самцом (моноандрический; Gadau et al., 1996), что приводит к высокой генетической однородности внутри колонии по сравнению с полигинными / полиандрическими видами. В лаборатории колония королевы содержалась в искусственном гипсовом гнезде при постоянной температуре 25 ° C и влажности около 40-50% с периодом фотосъемки 12:12 ч L / D.Колонию кормили водой с медом и мертвой саранчой два раза в неделю и обеспечивали водой ad libitum . Проведенные эксперименты и протоколы соответствуют «Руководству по уходу и использованию лабораторных животных Национальных институтов здравоохранения, опубликованному National Academic Press (1996)». Протокол был одобрен «Regierungspräsidium Freiburg» в соответствии с § 8a (TierSchG) и действующим законодательством Федеративной Республики Германии («Tierschutzgesetz»).
Протокол был одобрен «Regierungspräsidium Freiburg» в соответствии с § 8a (TierSchG) и действующим законодательством Федеративной Республики Германии («Tierschutzgesetz»).
Манипулирование профилями колоний субколоний
Из зрелой колонии (исходной колонии) мы создали четыре различных субколонии, которые содержались в отдельных гипсовых гнездах.Каждую субколонию обрабатывали по-разному, добавляя либо один, либо два синтетических углеводорода к запахам колоний субколоний. Для манипуляции мы использовали следующие углеводороды, которые не присутствовали в исходном профиле CHC C. floridanus : линейный алкантриаконтан (C30, Sigma-Aldrich) и далее называемый углеводородом A, был добавлен к двум суб- колонии (субколония A и субколония AB). Разветвленный алкан 5-метилгептакозан (5-MeC27), синтезированный для этого исследования и далее обозначаемый как углеводород B, был добавлен вместе с A в субколонию AB.Разветвленный алкан 11-метилгептакозан (11-MeC27), синтезированный для этого исследования, далее обозначаемый как углеводород C, добавляли в субколонию C.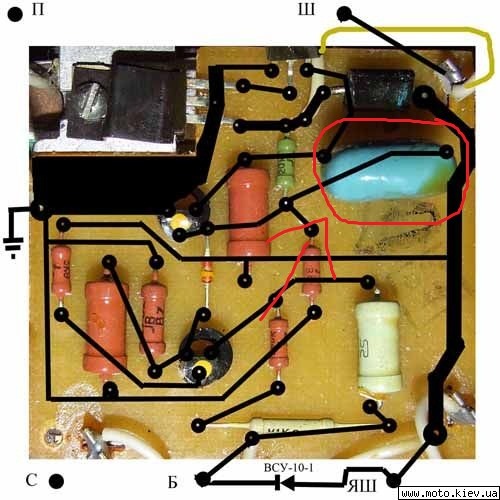 Одну субколонию ложно обрабатывали как контроль (субколония 0).
Одну субколонию ложно обрабатывали как контроль (субколония 0).
Мы использовали двухэтапный подход для управления профилями CHC. На первом этапе мы вызвали немедленное изменение профилей CHC всех рабочих перед их переносом в соответствующие субколонии. На втором этапе мы противодействовали уменьшению попадания углеводородов из кутикулы рабочих, предоставляя гранулы с углеводородным покрытием на входе в гнездо и в пище.
Для первого этапа манипуляции мы покрыли внутреннюю стенку колбы Эрленмейера (25 мл) соответствующим углеводородом (ами), добавив 3 мг углеводорода A (субколония A) или 3 мг углеводорода A и 3 мг углеводород B (субколония AB) или 6 мг углеводорода C (субколония C) и 6 мл пентана в колбу. Мы добавили относительно большое количество углеводорода C в субколонию C, чтобы вызвать сильное изменение в профилях CHC одной субколонии, которая с высокой вероятностью должна распознаваться как незнакомая и принадлежащая не-гнездовым товарищам.
Колбу осторожно встряхивали и ненадолго нагревали до ~ 60 ° C на горячей плите для испарения растворителя, а затем охлаждали до комнатной температуры. Мы случайным образом выбрали пять рабочих из исходной колонии, охладили их на льду и перенесли в покрытую колбу. Рабочих осторожно покручивали в колбе в течение 10–15 с, а затем переносили в новое гипсовое гнездо. Это повторялось до тех пор, пока не прошли лечение 100–150 рабочих. Рабочие субколонии 0 получали такую же обработку, за исключением того, что к пентану не добавляли углеводород (фиктивная обработка).
Мы случайным образом выбрали пять рабочих из исходной колонии, охладили их на льду и перенесли в покрытую колбу. Рабочих осторожно покручивали в колбе в течение 10–15 с, а затем переносили в новое гипсовое гнездо. Это повторялось до тех пор, пока не прошли лечение 100–150 рабочих. Рабочие субколонии 0 получали такую же обработку, за исключением того, что к пентану не добавляли углеводород (фиктивная обработка).
Для второго этапа манипуляции мы покрыли 600 мг гранул (суперабсорбирующие полимеры, натриевая соль полиакриловой кислоты, Stockosorb ® 500) 6 мл пентана и 3 мг углеводорода A (субколония A) или 3 мг. углеводорода A и 3 мг углеводорода B (субколония B) или 6 мг углеводорода C (субколония C) в химическом стакане. Стакан встряхивали и затем помещали на плиту с температурой 60 ° C для испарения пентана. Затем покрытые гранулы переносили в стеклянную пробирку и небольшие стеклянные пузырьки.Мы подключили стеклянную трубку (диаметр: 17 мм; длина: 100 мм) к гнезду (примерно 100 мм × 80 мм), которое служило длинной входной зоной. Рабочие, входящие в гнездо и выходящие из него, пассивно поглощали углеводороды от контакта с гранулами. Мы добавили воду с медом или воду в стеклянные флаконы и предоставили их субколониям, чтобы облегчить поглощение углеводородов во время кормления. После манипуляций в каждой субколонии 0, A и AB было примерно 150 взрослых рабочих, а в колонии C – примерно 100 рабочих.Мы предоставили выводок (яйца, личинки и куколки) каждой из субколоний, чтобы сохранить их естественную социальную структуру.
Рабочие, входящие в гнездо и выходящие из него, пассивно поглощали углеводороды от контакта с гранулами. Мы добавили воду с медом или воду в стеклянные флаконы и предоставили их субколониям, чтобы облегчить поглощение углеводородов во время кормления. После манипуляций в каждой субколонии 0, A и AB было примерно 150 взрослых рабочих, а в колонии C – примерно 100 рабочих.Мы предоставили выводок (яйца, личинки и куколки) каждой из субколоний, чтобы сохранить их естественную социальную структуру.
Все рабочие в субколониях, которые были протестированы позже в поведенческих экспериментах, были знакомы с предыдущим исходным профилем CHC, а в субколониях, подвергшихся манипуляции, рабочие также познакомились с новым профилем CHC. Таким образом, мы создали рабочих с разным опытом, знакомых с одним или двумя разными профилями CHC и незнакомых с тремя или двумя профилями CHC, для поддельных и обработанных субколоний, соответственно.
Поведенческий анализ
Через 1 день после манипуляций мы проводили встречи один на один в маленьких чашках Петри между двумя рабочими, либо из одной, либо из разных субколоний. Поскольку социальная изоляция рабочих снижает склонность к агрессии по отношению к не-гнездовым товарищам (Kleineidam et al., 2017), мы сначала переместили одного из рабочих (стимулирующего работника) в чашку Петри, а через 1 час мы начали эксперимент, добавив второй рабочий (ответственный) к блюду.Мы оценили поведение координатора во время первого взаимодействия со стимулом-работником как «агрессивное», когда координатор демонстрировал одно из действий: широко раскрытые нижние челюсти, щелканье, поднятие туловища или сгибание живота, все из которых являются типичными агрессивными. поведения для этого вида (Brandstaetter et al., 2008). Координаторы, которые не проявляли ни одного из этих видов поведения, были классифицированы как «неагрессивные».
Поскольку социальная изоляция рабочих снижает склонность к агрессии по отношению к не-гнездовым товарищам (Kleineidam et al., 2017), мы сначала переместили одного из рабочих (стимулирующего работника) в чашку Петри, а через 1 час мы начали эксперимент, добавив второй рабочий (ответственный) к блюду.Мы оценили поведение координатора во время первого взаимодействия со стимулом-работником как «агрессивное», когда координатор демонстрировал одно из действий: широко раскрытые нижние челюсти, щелканье, поднятие туловища или сгибание живота, все из которых являются типичными агрессивными. поведения для этого вида (Brandstaetter et al., 2008). Координаторы, которые не проявляли ни одного из этих видов поведения, были классифицированы как «неагрессивные».
Мы сделали вывод о распознавании и дискриминации сокамерников на основе поведенческих реакций координаторов.В случаях, когда координаторы не отвечали агрессией, мы классифицировали это как признание и возможное принятие в качестве одноклубников. В случаях, когда рабочие из субколонии демонстрируют низкую вероятность агрессии по отношению к стимулирующим работникам из разных субколоний, либо их профили CHC не дискриминируются, либо они дискриминируются, но наблюдатель не считывает их поведение. Таким образом, распознавание сокамерника рабочими, как классифицируется наблюдателем, либо является обобщением с предположением, что рабочие могут различать разнообразие профилей CHC, либо категоризация основана на недискриминационных профилях CHC.
В случаях, когда рабочие из субколонии демонстрируют низкую вероятность агрессии по отношению к стимулирующим работникам из разных субколоний, либо их профили CHC не дискриминируются, либо они дискриминируются, но наблюдатель не считывает их поведение. Таким образом, распознавание сокамерника рабочими, как классифицируется наблюдателем, либо является обобщением с предположением, что рабочие могут различать разнообразие профилей CHC, либо категоризация основана на недискриминационных профилях CHC.
Оценка поведенческих записей проводилась вслепую, при этом наблюдатель не знал тип стимула рабочего. Сразу после первого взаимодействия фокусные и стимулирующие работники были разделены либо для изучения профилей КГС (описанных ниже), либо координатор был переведен обратно в свою субколонию. В течение четырех последовательных дней после создания субколоний и управления запахами их колоний было проведено в общей сложности 360 встреч один на один за шесть сеансов с (почти) сбалансированными типами встреч. Для каждого из 12 возможных типов встреч проводилось 25–31 повторение; изменения в числах явились результатом слепого эксперимента. Поскольку некоторые из основных рабочих были переведены обратно в свои субколонии после эксперимента, возможно, что некоторые из основных рабочих были протестированы более одного раза на одной и той же субколонии. На основе моделирования (5000 раз) с количеством координаторов, которые не использовались для анализа их профилей CHC, мы подсчитали, что не более 7.5% всех протестированных фокусных работников были выбраны более одного раза для встречи с стимулом-работником из той же субколонии, что и раньше.
Для каждого из 12 возможных типов встреч проводилось 25–31 повторение; изменения в числах явились результатом слепого эксперимента. Поскольку некоторые из основных рабочих были переведены обратно в свои субколонии после эксперимента, возможно, что некоторые из основных рабочих были протестированы более одного раза на одной и той же субколонии. На основе моделирования (5000 раз) с количеством координаторов, которые не использовались для анализа их профилей CHC, мы подсчитали, что не более 7.5% всех протестированных фокусных работников были выбраны более одного раза для встречи с стимулом-работником из той же субколонии, что и раньше.
Анализ поведенческих данных
Во-первых, мы проверили, влияет ли время, прошедшее с момента основания субколонии, на вероятность агрессии у фокальных рабочих. Мы запустили обобщенную линейную модель (GLM) с переменной бинарной реакции агрессии («агрессия» / «отсутствие агрессии») и добавленным временем (экспериментальные сеансы 1–6) в качестве единственной объясняющей переменной. Поскольку оценка наклона не была значимой ( p > 0,05), мы не включили время в окончательную модель (модель 2).
Поскольку оценка наклона не была значимой ( p > 0,05), мы не включили время в окончательную модель (модель 2).
Во-вторых, мы проверили различия в вероятности агрессии у координационных работников по отношению к стимулирующим работникам из разных субколоний, используя GLM с бинарной переменной ответа агрессии («агрессия» / «отсутствие агрессии»), а в качестве объясняющих переменных – происхождение стимулирующего работника, происхождение ответственного работника и взаимодействие обеих переменных (модель 2).
Чтобы сделать выводы о различиях в поведении фокусных работников из разных субколоний, мы использовали байесовскую схему. Мы вычислили 20 000 значений, которые являются случайными выборками из апостериорного распределения оценок модели 2. Мы сравнили вероятность того, что работник будет агрессивен по отношению к работникам стимулирования из своей собственной колонии и из разных субколоний. Мы вычислили пропорции смоделированных значений из апостериорного распределения, которые были больше (или меньше) для фокусных рабочих, сталкивающихся с рабочими-стимулами из своей собственной субколонии, по сравнению с фокусными рабочими, сталкивающимися с рабочими-стимулами из другой субколонии (мера достоверности). Мы использовали квантили 2,5% и 97,5% в качестве нижней и верхней границы 95% вероятного интервала. Статистический анализ проводился с использованием R (v3.4.3; R Core Team, 2017), включая пакет arm с функцией sim для извлечения случайных выборок из апостериорного распределения параметров модели (Gelman and Hill, 2007). Уровень значимости для байесовской статистики был установлен на уровне достоверности> 99%.
Мы использовали квантили 2,5% и 97,5% в качестве нижней и верхней границы 95% вероятного интервала. Статистический анализ проводился с использованием R (v3.4.3; R Core Team, 2017), включая пакет arm с функцией sim для извлечения случайных выборок из апостериорного распределения параметров модели (Gelman and Hill, 2007). Уровень значимости для байесовской статистики был установлен на уровне достоверности> 99%.
Экстракции CHC
Чтобы убедиться, что наши манипуляции с профилями CHC были успешными, мы собрали рабочих для извлечения CHC после тестов на агрессию (всегда пары фокусных и стимулирующих рабочих) и обездвижили рабочих на льду.Каждого рабочего помещали в стеклянный флакон объемом 1 мл и полностью покрывали пентаном в течение 1 ч. Флакон осторожно встряхивали, а затем пентан переносили в чистый флакон со стеклянной вставкой на 200 мкл. Пентану давали возможность испариться при комнатной температуре, и запечатанные флаконы затем хранили в морозильной камере при -20 ° C до анализа.
Анализ профилей CHC
Флаконы размораживали до комнатной температуры, добавляли 1-3 капли пентана и встряхивали флаконы в течение 1 мин на вортексе.Затем мы сконцентрировали образец в постоянном потоке воздуха примерно до 20 мкл. Для масс-спектрометрического анализа с использованием парной газовой хроматографии (ГХ-МС) мы вручную вводили 1 мкл образцов в ГХ (Trace GC Ultra, соединенный с масс-спектрометром DSO II, Thermo Scientific). CHC анализировали на колонке из плавленого кварца (Optima-5-MS -0,25 мкм, 30 м × 0,25 мм, Macherey-Nagel GmbH and Co. KG) с гелием в качестве газа-носителя (1,2 мл / мин). Хроматограммы записывали с помощью программного обеспечения Xcalibur 1.4 SR1 (Thermo Scientific).Печь ГХ была запрограммирована следующим образом: (1) 70 ° C в течение 1 мин; (2) увеличение при 30 ° C / мин до 200 ° C; (3) повышение со скоростью 3 ° C / мин до 290 ° C; и (4) выдерживают при 290 ° C в течение 5 минут.
Мы проанализировали химическое сходство рабочих из разных субколоний с помощью анализа главных компонентов (PCA) на основе нормализованных площадей пиков, включая 10 наиболее заметных пиков из анализа GC-MS плюс сигнал во временах удерживания, когда три компонента (A, B, C) были обнаружены при их наличии.
Идентификация CHC
Мы провели стандартный раствор линейных углеводородов в диапазоне от 21 до 40 атомов углерода, используя тот же метод, что и для профилей CHC.Мы рассчитали индекс Коваца для каждого пика профиля. Соединения CHC были идентифицированы на основе их индексов Коваца и их масс-спектров, которые сравнивали с записями известных компонентов в библиотеках Wiley и NIST.
Синтез метил-разветвленных углеводородов
5-метилгептакозан
Бутиллитий (2,2 М в гексане) по каплям добавляли к суспензии бромида докозилтрифенилфосфония (3,26 г, 5 ммоль) в 100 мл сухого тетрагидрофурана (ТГФ) в высушенной в печи колбе в атмосфере аргона до тех пор, пока смесь не приобрела розовый оттенок, с последующим добавлением добавление еще 2.5 мл раствора бутиллития (5,5 ммоль). Полученный прозрачный вишнево-красный раствор перемешивали 30 минут с последующим добавлением 2-гексанона (0,50 г, 5 ммоль) в 5 мл ТГФ с помощью шприцевого насоса в течение 30 минут. Полученную смесь перемешивали в течение ночи, затем гасили добавлением насыщенного водного NH 4 Cl. Затем смесь разбавляли водой и экстрагировали гексаном. Гексановый слой промывали водой и рассолом, сушили над безводным Na 2 SO 4 , концентрировали и очищали вакуумной флэш-хроматографией на силикагеле, элюируя гексаном.Полученный полукристаллический продукт помещали в 25 мл гексана в колбу на 50 мл, добавляли катализатор 5% Pd на угле (250 мг), и колбу промывали водородом, герметично закрывали и перемешивали под водородом в течение 3 часов, когда анализ методом ГХ. показал, что весь исходный материал израсходован. Смесь фильтровали через пробку из целита, хорошо промывая гексаном, и после концентрирования полученное твердое вещество перекристаллизовывали из 30 мл ацетона в течение ночи при 4 ° C. После фильтрации получали продукт в виде белых кристаллов (0.60 г). Масс-спектр EI (70 эВ; m / z, содержание): 394 (M +, след), 379 (7), 365 (7), 351 (3), 337 (54), 336 (31), 309 (7) , 308 (10), 295 (1), 281 (2), 267 (2), 253 (3), 239 (3), 225 (3), 211 (3), 197 (4), 183 (5) , 169 (5), 155 (7), 141 (8), 127 (10), 113 (13), 99 (19), 85 (100), 84 (73), 71 (56), 57 (81) , 43 (71).
Полученную смесь перемешивали в течение ночи, затем гасили добавлением насыщенного водного NH 4 Cl. Затем смесь разбавляли водой и экстрагировали гексаном. Гексановый слой промывали водой и рассолом, сушили над безводным Na 2 SO 4 , концентрировали и очищали вакуумной флэш-хроматографией на силикагеле, элюируя гексаном.Полученный полукристаллический продукт помещали в 25 мл гексана в колбу на 50 мл, добавляли катализатор 5% Pd на угле (250 мг), и колбу промывали водородом, герметично закрывали и перемешивали под водородом в течение 3 часов, когда анализ методом ГХ. показал, что весь исходный материал израсходован. Смесь фильтровали через пробку из целита, хорошо промывая гексаном, и после концентрирования полученное твердое вещество перекристаллизовывали из 30 мл ацетона в течение ночи при 4 ° C. После фильтрации получали продукт в виде белых кристаллов (0.60 г). Масс-спектр EI (70 эВ; m / z, содержание): 394 (M +, след), 379 (7), 365 (7), 351 (3), 337 (54), 336 (31), 309 (7) , 308 (10), 295 (1), 281 (2), 267 (2), 253 (3), 239 (3), 225 (3), 211 (3), 197 (4), 183 (5) , 169 (5), 155 (7), 141 (8), 127 (10), 113 (13), 99 (19), 85 (100), 84 (73), 71 (56), 57 (81) , 43 (71).
11-метилгептакозан
11-Метилгептакозан получали аналогичным образом, начиная с гексадецилтрифенилфосфонийбромида и 2-додеканона, за исключением того, что конечный продукт перекристаллизовывали сначала из изооктана, а затем снова из ацетона при 4 ° C, давая белые кристаллы.Масс-спектр EI (70 эВ; m / z, содержание): 394 (M +, след), 379 (5), 365 (3), 351 (2), 337 (2), 323 (2), 309 (1) , 295 (1), 281 (1), 267 (1), 253 (22), 252 (26), 239 (1), 225 (3), 224 (7), 211 (2), 197 (3) , 183 (4), 169 (21), 168 (45), 155 (5), 141 (7), 140 (7), 127 (11), 113 (18), 99 (27), 85 (65) , 71 (80), 57 (100), 43 (53).
Результаты
Дискриминация различных профилей CHC
Мы тестировали рабочих из разных субколоний в ходе встреч один на один и использовали агрессивные реакции как меру определения рабочими категории рабочих встреч как «товарищей по гнезду» или «товарищей по гнезду».«Наши манипуляции позволили нам ответить на вопрос, связана ли агрессия либо с химическим сходством, либо с опытом работника с различными профилями CHC (знакомыми и незнакомыми профилями CHC).
Профиль CHC субколонии, в которую были добавлены как линейные, так и метилированные углеводороды (субколония AB), больше похож на профиль CHC субколонии, где был добавлен только метилированный углеводород (субколония A ), чем из субколонии, подвергшейся ложной обработке (субколония 0).
Почти во всех встречах один на один (353 из 360) между координаторами и сотрудниками по стимулированию мы смогли классифицировать поведение координаторов как агрессивное или неагрессивное. В семи встречах мы не смогли классифицировать поведение координатора, потому что один из двух рабочих сбежал с арены, перелез через стену, таким образом избежав столкновения.
Мы редко встречали агрессию в столкновениях между фокусными и стимулирующими работниками из совпадающих (одних и тех же) субколоний (10.7% –16,7%, рис. 2), что указывает на то, что согласованность внутри субколоний сохраняется.
Рисунок 2 . Агрессия координаторов по отношению к стимулирующим работникам из разных субколоний. Фокусные работники из всех субколоний редко проявляли агрессию по отношению к стимуляторам из своей субколонии (совпадающие цвета) или из поддельно обработанной субколонии 0 (синие квадраты). Напротив, значительное количество координаторов из всех субколоний проявили агрессию по отношению к стимулирующим работникам из субколонии C (зеленые ромбы).Фокусные работники из субколонии 0 значительно чаще проявляли агрессию по отношению к стимулирующим работникам из субколонии AB (красные треугольники), но не так по отношению к стимулирующим работникам из субколонии A (оранжевые кружки). Фокусные работники субколонии A проявляли агрессию значительно чаще по отношению к стимулирующим работникам из субколонии AB (красные треугольники), чем к стимулирующим работникам из их собственной субколонии. Фокусные работники субколонии AB проявляли агрессию значительно чаще по отношению к стимулирующим работникам из субколонии A (оранжевый кружок), чем к стимулирующим работникам из их собственной субколонии.
Фокусные работники из всех субколоний редко проявляли агрессию по отношению к стимуляторам из своей субколонии (совпадающие цвета) или из поддельно обработанной субколонии 0 (синие квадраты). Напротив, значительное количество координаторов из всех субколоний проявили агрессию по отношению к стимулирующим работникам из субколонии C (зеленые ромбы).Фокусные работники из субколонии 0 значительно чаще проявляли агрессию по отношению к стимулирующим работникам из субколонии AB (красные треугольники), но не так по отношению к стимулирующим работникам из субколонии A (оранжевые кружки). Фокусные работники субколонии A проявляли агрессию значительно чаще по отношению к стимулирующим работникам из субколонии AB (красные треугольники), чем к стимулирующим работникам из их собственной субколонии. Фокусные работники субколонии AB проявляли агрессию значительно чаще по отношению к стимулирующим работникам из субколонии A (оранжевый кружок), чем к стимулирующим работникам из их собственной субколонии. Символы представляют собой подобранные значения из биномиальной обобщенной линейной модели (GLM; модель 2), а вертикальные линии представляют 95% вероятных интервалов, основанных на байесовской статистике. Были протестированы от 25 до 31 рабочего из каждой группы. Звездочки указывают уровни достоверности выше 99% для различий между совпадающими (из одной субколонии) и несовпадающими (из разных субколоний) парами.
Символы представляют собой подобранные значения из биномиальной обобщенной линейной модели (GLM; модель 2), а вертикальные линии представляют 95% вероятных интервалов, основанных на байесовской статистике. Были протестированы от 25 до 31 рабочего из каждой группы. Звездочки указывают уровни достоверности выше 99% для различий между совпадающими (из одной субколонии) и несовпадающими (из разных субколоний) парами.
Большинство фокусных работников из управляемой субколонии А также не проявляли агрессии по отношению к стимулирующим рабочим из поддельно обработанной субколонии (субколония 0; рисунок 2 синий; 17.2%), что показывает, что исходный запах колонии все еще сохраняется. Фокусные работники из субколонии AB чаще (уровень достоверности: 93%) реагировали агрессией по отношению к стимулирующим работникам из субколонии 0 (26,7%), чем к стимулирующим работникам из своей субколонии (10,7%), что указывает на то, что примерно 3/4 рабочих субколонии 0 все еще принимаются.
Процент фокусных рабочих из субколонии 0, которые проявили агрессию по отношению к стимулирующим работникам, получавшим A, также был низким (22,6%), и мы обнаружили мало поддержки того, что это отличается от реакции рабочих на стимулирующих рабочих из их собственной субколонии ( уровень достоверности только 71. 6%). Похоже, что присутствие компонента A (линейный алкан C30) в стимуляторах не означает статус не-гнездящихся для фокусных работников из субколонии 0. Напротив, фокальные рабочие из субколонии 0 и субколонии A проявляли агрессию по отношению к стимулирование рабочих из субколонии AB (рост агрессии на 42% и 34% соответственно; уровень достоверности> 99% в обоих случаях). Таким образом, добавление двух компонентов A и B (C30 и 5-MeC27) к профилю CHC рабочих-стимулов вызывает агрессивные ответы у фокусных рабочих из субколоний, в которых ни одного (субколония 0) или только один из двух компонентов ( C30 в субколонии A) использовали для управления запахом колонии.Более того, координационные работники из субколонии AB значительно чаще реагировали агрессией по отношению к стимулирующим работникам из субколонии A (38,7%), чем к стимулирующим работникам из своей субколонии (10,7%). В этом тестовом сценарии отсутствие компонента B в профиле CHC у стимулирующих рабочих вызывает более агрессивное поведение у фокусных рабочих субколонии AB по сравнению с столкновениями с их текущими товарищами по гнезду (уровень достоверности> 99%).
6%). Похоже, что присутствие компонента A (линейный алкан C30) в стимуляторах не означает статус не-гнездящихся для фокусных работников из субколонии 0. Напротив, фокальные рабочие из субколонии 0 и субколонии A проявляли агрессию по отношению к стимулирование рабочих из субколонии AB (рост агрессии на 42% и 34% соответственно; уровень достоверности> 99% в обоих случаях). Таким образом, добавление двух компонентов A и B (C30 и 5-MeC27) к профилю CHC рабочих-стимулов вызывает агрессивные ответы у фокусных рабочих из субколоний, в которых ни одного (субколония 0) или только один из двух компонентов ( C30 в субколонии A) использовали для управления запахом колонии.Более того, координационные работники из субколонии AB значительно чаще реагировали агрессией по отношению к стимулирующим работникам из субколонии A (38,7%), чем к стимулирующим работникам из своей субколонии (10,7%). В этом тестовом сценарии отсутствие компонента B в профиле CHC у стимулирующих рабочих вызывает более агрессивное поведение у фокусных рабочих субколонии AB по сравнению с столкновениями с их текущими товарищами по гнезду (уровень достоверности> 99%). С вероятностью 83,8% рабочие субколонии AB более агрессивны по отношению к рабочим из субколонии A, чем по отношению к рабочим из субколонии 0.Таким образом, как наличие дополнительного компонента, так и отсутствие определенного компонента может изменить профиль CHC соседа по гнезду на профиль другого члена.
С вероятностью 83,8% рабочие субколонии AB более агрессивны по отношению к рабочим из субколонии A, чем по отношению к рабочим из субколонии 0.Таким образом, как наличие дополнительного компонента, так и отсутствие определенного компонента может изменить профиль CHC соседа по гнезду на профиль другого члена.
Кроме того, мы протестировали фокусных рабочих из всех трех субколоний (0, A, AB) против стимулирующих рабочих из другой субколонии, обработанной метилированным углеводородом в высокой концентрации (11-MeC27, субколония C), для количественной оценки агрессии против чрезмерно манипулируемого профиля CHC, который следует легко распознать как незнакомый и принадлежащий не-сокамерникам.Все фокусные работники, независимо от того, из какой субколонии они были выбраны, проявили значительную агрессию по отношению к стимулирующим работникам из субколонии C (уровни достоверности> 99%, рисунок 2 зеленый).
Манипуляции с профилями CHC
После поведенческих экспериментов мы собрали рабочих и проанализировали их профили CHC, чтобы убедиться, что синтетические углеводороды были перенесены в их кутикулу в результате наших манипуляций. Анализ методом ГХ-МС промывок кутикулы у 38 отдельных рабочих показал, что наша химическая обработка запаха колонии была успешной.Экстракты ХГК индивидуумов содержали те же самые известные ХГК, что и поддельно обработанная субколония (субколония 0), плюс один или два дополнительных углеводорода в зависимости от манипуляции (Таблица 1). Дополнительные углеводороды A и B были обнаружены в виде отдельных пиков на хроматограммах при временах удерживания, когда не появлялись никакие компоненты исходного профиля CHC C. floridanus (рис. 3). Основываясь на данных GC-MS, мы не можем исключить, что мы потенциально собрали синтетические углеводороды, которые были поглощены муравьями, но не представлены в составе CHC на кутикуле.
Анализ методом ГХ-МС промывок кутикулы у 38 отдельных рабочих показал, что наша химическая обработка запаха колонии была успешной.Экстракты ХГК индивидуумов содержали те же самые известные ХГК, что и поддельно обработанная субколония (субколония 0), плюс один или два дополнительных углеводорода в зависимости от манипуляции (Таблица 1). Дополнительные углеводороды A и B были обнаружены в виде отдельных пиков на хроматограммах при временах удерживания, когда не появлялись никакие компоненты исходного профиля CHC C. floridanus (рис. 3). Основываясь на данных GC-MS, мы не можем исключить, что мы потенциально собрали синтетические углеводороды, которые были поглощены муравьями, но не представлены в составе CHC на кутикуле.
Таблица 1 . Относительные количества (среднее ± стандартное отклонение) 10 наиболее распространенных кутикулярных углеводородов (CHC) в экстрактах, полученных от отдельных рабочих (нормализованные к общей площади пика).
Рисунок 3 . Манипулирование профилями CHC различных субколоний. Примеры хроматограмм одиночных рабочих из субколоний 0, A, AB и C. Компоненты обозначены буквами и цифрами: A: C30, B: Me-5C27, C: Me-11C27, 1: 3,7-диметил – и 3,9-диметилнонакозан, 2: 4-метилтриаконтан, 3: 4,10-диметил- и 4,8-диметилтриаконтан, 4: 9-метилгентриаконтан и неизвестно, 5: 5,9-ди -метилгентриаконтан, 6: 8-метил- и 12-метилдотриаконтан, 7: 3,7,11-три-метилгентриаконтан, 8 + 9 + 10: неизвестно.Обратите внимание, что в этих примерах два компонента A и B в разном количестве интегрированы в профиль CHC.
Манипулирование профилями CHC различных субколоний. Примеры хроматограмм одиночных рабочих из субколоний 0, A, AB и C. Компоненты обозначены буквами и цифрами: A: C30, B: Me-5C27, C: Me-11C27, 1: 3,7-диметил – и 3,9-диметилнонакозан, 2: 4-метилтриаконтан, 3: 4,10-диметил- и 4,8-диметилтриаконтан, 4: 9-метилгентриаконтан и неизвестно, 5: 5,9-ди -метилгентриаконтан, 6: 8-метил- и 12-метилдотриаконтан, 7: 3,7,11-три-метилгентриаконтан, 8 + 9 + 10: неизвестно.Обратите внимание, что в этих примерах два компонента A и B в разном количестве интегрированы в профиль CHC.
Мы визуализировали сходство профилей CHC между субколониями с помощью PCA. Профили CHC рабочих были распределены вдоль PC1, тогда как рабочие из субколоний A и AB были отделены от рабочих субколонии C вдоль PC2 (Рисунок 4). Химически наиболее однородной группой были рабочие из субколонии 0, и в среднем рабочие из субколонии A были химически более похожими, чем рабочие из субколонии AB, по сравнению с рабочими из субколонии 0.
Рисунок 4 . Анализ главных компонентов (PCA) профилей CHC, полученных от отдельных рабочих. Рабочие систематически различаются в зависимости от лечения и ориентированы на разные оси. PCA показывает, что количество компонентов, интегрированных в профиль CHC в определенных субколониях, варьируется. Эллипсы представляют собой 95% доверительные области. Синий: 11 рабочих субколонии 0, желтый: 12 рабочих субколонии A, красный: восемь рабочих субколонии AB, зеленый: семь рабочих субколонии C.Серые стрелки указывают загрузки тех же 13 углеводородов, что и в таблице 1.
Обсуждение
Мы исследовали, используют ли рабочие общие черты, обнаруженные в профилях CHC, для распознавания сокамерников, или же распознавание товарищей по гнезду основано на обобщении отдельных профилей CHC, которые могут различаться работниками. Мы успешно управляли запахом колоний в различных субколониях, добавляя один или два разных углеводорода, которые не присутствовали в исходных профилях CHC в C. floridanus на уровнях, аналогичных (для компонентов A и B, а не для компонента C) количеству исходных углеводородов. Это позволило нам протестировать эффекты добавления новых компонентов в профиль, а также протестировать эффект отсутствия одного компонента во всем остальном очень похожем профиле CHC. Мы обнаружили, что рабочие могут настраивать распознавание своих сокамерников на новые измененные профили CHC, но при этом принимать работников как сокамерников, несущих предыдущий профиль CHC. Кроме того, мы показываем, что дополнительные компоненты, а также отсутствие компонента в профиле CHC могут быть различены и могут отображать метку как принадлежащую другим членам.Рабочие, имеющие как линейный, так и метил-разветвленный углеводород в своем новом профиле CHC, воспринимают рабочих, у которых есть только линейный углеводород, но не метилированный углеводород, как не являющихся соседями. Таким образом, наши результаты подтверждают гипотезу о том, что рабочие могут распознавать сокамерников с разными профилями CHC в процессе распознавания нескольких шаблонов и классифицировать их как всех, принадлежащих к товарищам по гнезду.
floridanus на уровнях, аналогичных (для компонентов A и B, а не для компонента C) количеству исходных углеводородов. Это позволило нам протестировать эффекты добавления новых компонентов в профиль, а также протестировать эффект отсутствия одного компонента во всем остальном очень похожем профиле CHC. Мы обнаружили, что рабочие могут настраивать распознавание своих сокамерников на новые измененные профили CHC, но при этом принимать работников как сокамерников, несущих предыдущий профиль CHC. Кроме того, мы показываем, что дополнительные компоненты, а также отсутствие компонента в профиле CHC могут быть различены и могут отображать метку как принадлежащую другим членам.Рабочие, имеющие как линейный, так и метил-разветвленный углеводород в своем новом профиле CHC, воспринимают рабочих, у которых есть только линейный углеводород, но не метилированный углеводород, как не являющихся соседями. Таким образом, наши результаты подтверждают гипотезу о том, что рабочие могут распознавать сокамерников с разными профилями CHC в процессе распознавания нескольких шаблонов и классифицировать их как всех, принадлежащих к товарищам по гнезду. Различают ли они разные профили CHC сородичей, еще предстоит проверить.
Различают ли они разные профили CHC сородичей, еще предстоит проверить.
Дискриминация профилей CHC
Для наших манипуляций с профилями CHC мы использовали два метил-разветвленных и один неразветвленный углеводород, зная, что углеводороды, содержащие метильные группы, могут быть более важными в качестве сигналов распознавания по сравнению с неразветвленными алканами (Dani et al., 2001; Guerrieri et al., 2009; Martin S. и Drijfhout F., 2009), но см. (Akino et al., 2004; Greene and Gordon, 2007). Действительно, мы обнаружили, что добавление линейного алкана (C30) к профилю CHC не оказало значительного влияния на распознавание сородичей у рабочих, у которых не было этого компонента в запахе собственной колонии. Мы предполагаем, что компоненты профиля CHC могут быть незначительными или важными сигналами распознавания, в зависимости от того, обладает ли их присутствие прогностической силой для дискриминации, расширяя концепцию ключевых сигналов распознавания в профилях CHC (Dani et al.
Наши наблюдения, что после изменения запаха колонии новый, а также предыдущий профиль CHC принимается для распознавания сородичей, привели к предположению, что лежащий в основе механизм – это расширение диапазона допустимости шаблона для ярлыков сородичей (Meskali et al. ., 1995; Leonhardt et al., 2007; Guerrieri et al., 2009). В одном случае агрессия против рабочих с предыдущим профилем CHC была сравнительно высокой (рабочие из субколонии AB по отношению к рабочим субколонии 0).Мы не знаем причины этого, но мы рассматриваем вариацию средней вероятности агрессии в разных группах, протестированных по сравнению с их предыдущим профилем CHC, в результате небольшого размера выборки.
Представленные здесь результаты не могут быть объяснены в рамках различных меток, распознаваемых с помощью унифицирующего шаблона. Унифицирующий шаблон будет основан на общих чертах, обнаруженных в большинстве профилей CHC товарищей по гнезду, и их соответствующем нейронном представлении, что привело к концепции «теории включения» (Guerrieri et al. , 2009; Bos et al., 2012, 2013). Однако в одном из наших тестовых сценариев (рабочие из субколонии AB встречают рабочих из субколонии A) критерий включения в отношении химического состава профилей CHC был выполнен. Мы показали, что рабочие по-прежнему могут различать на основе профилей CHC, которые содержат все, кроме одного из компонентов, по сравнению с их собственным (измененным) запахом колонии. Таким образом, что касается меток, то есть состава профилей CHC, дискриминация рабочих не может быть объяснена теорией включения, потому что дополнительные компоненты не были более важны, чем отсутствующие компоненты для распознавания сородичей, как предполагалось ранее (Guerrieri et al. ., 2009).
, 2009; Bos et al., 2012, 2013). Однако в одном из наших тестовых сценариев (рабочие из субколонии AB встречают рабочих из субколонии A) критерий включения в отношении химического состава профилей CHC был выполнен. Мы показали, что рабочие по-прежнему могут различать на основе профилей CHC, которые содержат все, кроме одного из компонентов, по сравнению с их собственным (измененным) запахом колонии. Таким образом, что касается меток, то есть состава профилей CHC, дискриминация рабочих не может быть объяснена теорией включения, потому что дополнительные компоненты не были более важны, чем отсутствующие компоненты для распознавания сородичей, как предполагалось ранее (Guerrieri et al. ., 2009).
Нейронные механизмы для сопоставления метки и шаблона и формирования нового шаблона
Муравьи узнают об запахе своей колонии в раннем взрослом возрасте (Carlin and Hölldobler, 1983; Errard, 1994) и о новых ярлыках позже в жизни (Guerrieri et al., 2009), как и в наших экспериментах. Поскольку они сопоставляют метки с нейронным шаблоном, мы предполагаем, что это происходит в грибовидном теле (МБ) – области мозга, которая важна для обучения и памяти (Heisenberg, 2003; Giurfa, 2007).Компоненты метки обнаруживаются широко настроенными нейронами обонятельных рецепторов (ORN; Sharma et al., 2015), которые проецируются в антеннальную долю (AL), где ORN, экспрессирующие одни и те же обонятельные рецепторы, объединяются в одних и тех же функциональных единицах (Couto et al. ., 2005; Зубе и др., 2008; рис. 5А). Здесь паттерны активности, специфичные для меток, переформатируются и передаются через проекционные нейроны (PN) в высшие мозговые центры, такие как MB (Deisig et al., 2010; Brandstaetter and Kleineidam, 2011). В чашечках MB синапсы PN с клетками Kenyon (KC) преобразуют плотные и перекрывающиеся паттерны активности в разреженные и гораздо менее перекрывающиеся паттерны активности (Perez-Orive et al., 2002; Szyszka et al., 2005). В поисках нейронного субстрата шаблонов мы перенимаем современные знания о процессах оценки и обучения в MB плодовой мушки Drosophila (Aso et al.
Поскольку они сопоставляют метки с нейронным шаблоном, мы предполагаем, что это происходит в грибовидном теле (МБ) – области мозга, которая важна для обучения и памяти (Heisenberg, 2003; Giurfa, 2007).Компоненты метки обнаруживаются широко настроенными нейронами обонятельных рецепторов (ORN; Sharma et al., 2015), которые проецируются в антеннальную долю (AL), где ORN, экспрессирующие одни и те же обонятельные рецепторы, объединяются в одних и тех же функциональных единицах (Couto et al. ., 2005; Зубе и др., 2008; рис. 5А). Здесь паттерны активности, специфичные для меток, переформатируются и передаются через проекционные нейроны (PN) в высшие мозговые центры, такие как MB (Deisig et al., 2010; Brandstaetter and Kleineidam, 2011). В чашечках MB синапсы PN с клетками Kenyon (KC) преобразуют плотные и перекрывающиеся паттерны активности в разреженные и гораздо менее перекрывающиеся паттерны активности (Perez-Orive et al., 2002; Szyszka et al., 2005). В поисках нейронного субстрата шаблонов мы перенимаем современные знания о процессах оценки и обучения в MB плодовой мушки Drosophila (Aso et al. , 2014; Cohn et al., 2015; Hige et al., 2015a). , б) и медоносной пчелы (Strube-Bloss et al., 2011, 2016), чтобы предложить следующий механизм сопоставления метки-шаблона.
, 2014; Cohn et al., 2015; Hige et al., 2015a). , б) и медоносной пчелы (Strube-Bloss et al., 2011, 2016), чтобы предложить следующий механизм сопоставления метки-шаблона.
Рисунок 5 . Предложена упрощенная схемная модель для формирования шаблона и сопоставления метки с шаблоном. (A) Признаки распознавания (RC) от стимулирующего муравья принимаются широко настроенными нейронами обонятельных рецепторов (ORN) на антеннах (AN) оценивающего муравья. Активность, индуцированная запахом, переформатируется по обонятельному пути через антеннальную долю (AL) и проекционные нейроны (PN), что приводит к специфическим для меток паттернам активации клеток Кеньона (KC) в грибовидном теле (MB). В этом примере активность, индуцированная меткой AB в KC, обозначена толстыми красными линиями. Во время изучения нового лейбла (например,g., метка AB), вознаграждение приводит к активации дофаминергических нейронов (DAN) в одном из двух отсеков. Совпадающая активность DAN и KC в этом компартменте приводит к депрессии синапсов между KC и MBON. Основываясь на относительной выходной мощности из разных компартментов МБ, обеспечивается положительная, нейтральная или отрицательная валентность, принимается решение, и муравей-фокус в конечном итоге отвечает поведенческим действием по отношению к муравью-стимулу. В этом примере метка AB активирует определенный набор KC в процессе обучения.Синапсы от KCs, которые также активны, когда представлена метка 0, уже подавлены (от предыдущих обучающих событий), тогда как синапсы от метки AB-специфичных KCs подвергаются депрессии из-за совпадающей активности в DAN (пунктирный кружок). (B) Примеры синаптической передачи между KC и MBON в двух разных компартментах после изучения метки 0 и метки AB как соседа по гнезду. Все три метки (метка 0, A и AB) активируют разные наборы и одинаковое количество KC (цветные толстые линии указывают на активность; серые тонкие линии указывают на отсутствие активности).Из-за более слабых синапсов между KC и MBON в отрицательном компартменте метка 0 и метка AB имеют общую положительную валентность, тогда как метка A имеет нейтральную валентность.
Основываясь на относительной выходной мощности из разных компартментов МБ, обеспечивается положительная, нейтральная или отрицательная валентность, принимается решение, и муравей-фокус в конечном итоге отвечает поведенческим действием по отношению к муравью-стимулу. В этом примере метка AB активирует определенный набор KC в процессе обучения.Синапсы от KCs, которые также активны, когда представлена метка 0, уже подавлены (от предыдущих обучающих событий), тогда как синапсы от метки AB-специфичных KCs подвергаются депрессии из-за совпадающей активности в DAN (пунктирный кружок). (B) Примеры синаптической передачи между KC и MBON в двух разных компартментах после изучения метки 0 и метки AB как соседа по гнезду. Все три метки (метка 0, A и AB) активируют разные наборы и одинаковое количество KC (цветные толстые линии указывают на активность; серые тонкие линии указывают на отсутствие активности).Из-за более слабых синапсов между KC и MBON в отрицательном компартменте метка 0 и метка AB имеют общую положительную валентность, тогда как метка A имеет нейтральную валентность. Функциональная связь DAN и MBON в MB получена из исследований на Drosophila (Aso et al., 2014). Цветовой код KC: черный, неспецифический KC для запахов колоний; синий, метка 0, специфический KC; оранжевый, метка A специфический KC, красный, метка AB специфический KC. LN: локальные интернейроны. (C) На основе предложенной нами модели шаблон реализуется как уменьшенная синаптическая сила между специфическими для метки nestmate KCs и MBON в отрицательном компартменте MB по сравнению с положительным компартментом.
Функциональная связь DAN и MBON в MB получена из исследований на Drosophila (Aso et al., 2014). Цветовой код KC: черный, неспецифический KC для запахов колоний; синий, метка 0, специфический KC; оранжевый, метка A специфический KC, красный, метка AB специфический KC. LN: локальные интернейроны. (C) На основе предложенной нами модели шаблон реализуется как уменьшенная синаптическая сила между специфическими для метки nestmate KCs и MBON в отрицательном компартменте MB по сравнению с положительным компартментом.
Многие KCs (~ 2000 у Drosophila и ~ 170 000 у медоносных пчел; Witthöft, 1967; Aso et al., 2014) синапсы на несколько выходных нейронов (MBON; 34 у Drosophila и ~ 400 у медоносных пчел; Рыбак и Menzel, 1993; Tanaka et al., 2008) в разных компартментах МБ. Один отсек МБ положительный, а другой соответствующий отрицательный, каждый из которых содержит внешний МБОН. Мы постулируем, что метка будет ассоциироваться с положительной или отрицательной валентностью, когда сила активности MBON из разных компартментов становится несбалансированной (рис. 5B).Несбалансированная активность MBON является результатом специфических для компартментов различий в синаптической передаче от KCs к MBON, и мы предполагаем, что синаптическая передача снижается в компартменте, обеспечивающем отрицательную валентность (Hige et al., 2015a). Нижняя синаптическая передача ограничена теми синапсами KC, которые активируются меткой nestmate. Основываясь на этой структуре, нейронный шаблон будет реализован как более слабая синаптическая сила между KC и MBON в отрицательном компартменте по сравнению с положительным компартментом (Рис. 5C).Пластичность синапсов KC может быть вызвана совпадающей активностью дофаминергических нейронов, специфичных для компартментов (DAN), что преимущественно приводит к синаптической депрессии (медоносная пчела: Mauelshagen, 1993; Grünewald, 1999; Drosophila : Hige et al., 2015a), но может дополнительно привести к рекрутированию активности MBON (см. Okada et al., 2007; Strube-Bloss et al., 2011, 2016). Формирование шаблона посредством ассоциативного обучения требует активации пути DAN вознаграждением (например, социальным взаимодействием или едой), а также, возможно, периодической активацией DAN из MBON (Zhao et al.
5B).Несбалансированная активность MBON является результатом специфических для компартментов различий в синаптической передаче от KCs к MBON, и мы предполагаем, что синаптическая передача снижается в компартменте, обеспечивающем отрицательную валентность (Hige et al., 2015a). Нижняя синаптическая передача ограничена теми синапсами KC, которые активируются меткой nestmate. Основываясь на этой структуре, нейронный шаблон будет реализован как более слабая синаптическая сила между KC и MBON в отрицательном компартменте по сравнению с положительным компартментом (Рис. 5C).Пластичность синапсов KC может быть вызвана совпадающей активностью дофаминергических нейронов, специфичных для компартментов (DAN), что преимущественно приводит к синаптической депрессии (медоносная пчела: Mauelshagen, 1993; Grünewald, 1999; Drosophila : Hige et al., 2015a), но может дополнительно привести к рекрутированию активности MBON (см. Okada et al., 2007; Strube-Bloss et al., 2011, 2016). Формирование шаблона посредством ассоциативного обучения требует активации пути DAN вознаграждением (например, социальным взаимодействием или едой), а также, возможно, периодической активацией DAN из MBON (Zhao et al. , 2018). Знакомство с новой меткой, которая в конечном итоге становится действительной для сокамерников, приведет к синаптической депрессии во всех синапсах KC, которые активируются этой новой меткой в компартменте, обеспечивающем отрицательную валентность. Синаптическая депрессия из-за разных меток nestmate может происходить на одном и том же MBON, и мы предполагаем, что высокая специфичность к разным меткам реализуется за счет большого количества KC и их редких паттернов активности для разных меток. В частности, мы предполагаем, что есть KC, которые демонстрируют свойство ответа на один стимул (например,g., метка A), но не к аналогичному стимулу, где добавлен только один дополнительный компонент (например, метка AB; рисунок 5B). Обратите внимание, что у Drosophila такие KCs (отвечающие на один компонент, но не в комбинации со вторым компонентом) еще не описаны (Campbell et al., 2013). Однако исследования Drosophila и медоносных пчел предполагают, что такая синтетическая обработка информации о компонентах происходит в AL (Joerges et al.
, 2018). Знакомство с новой меткой, которая в конечном итоге становится действительной для сокамерников, приведет к синаптической депрессии во всех синапсах KC, которые активируются этой новой меткой в компартменте, обеспечивающем отрицательную валентность. Синаптическая депрессия из-за разных меток nestmate может происходить на одном и том же MBON, и мы предполагаем, что высокая специфичность к разным меткам реализуется за счет большого количества KC и их редких паттернов активности для разных меток. В частности, мы предполагаем, что есть KC, которые демонстрируют свойство ответа на один стимул (например,g., метка A), но не к аналогичному стимулу, где добавлен только один дополнительный компонент (например, метка AB; рисунок 5B). Обратите внимание, что у Drosophila такие KCs (отвечающие на один компонент, но не в комбинации со вторым компонентом) еще не описаны (Campbell et al., 2013). Однако исследования Drosophila и медоносных пчел предполагают, что такая синтетическая обработка информации о компонентах происходит в AL (Joerges et al.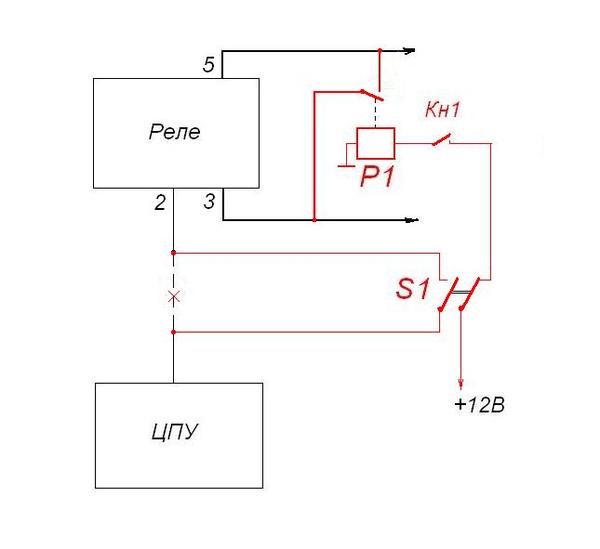 , 1997; Silbering and Galizia, 2007; Silbering et al., 2008; Deisig et al., 2010). Более того, ассоциативное обучение запаху изменяет паттерны активности функциональных единиц AL, что увеличивает разницу между смесями и их компонентами (Faber et al., 1999; Daly et al., 2004; Fernandez et al., 2009; Rath et al. , 2011). Для нашего исследования это подразумевает модификацию паттернов активности AL для метки с помощью одного дополнительного компонента (метка A), тем самым отделяя его от шаблона активности для метки с тем же самым и добавленным другим компонентом (метка AB).Наш результат о том, что этикетка с одним дополнительным компонентом является «странной» только для тех рабочих, которые узнали этикетку с двумя дополнительными компонентами как действительную, также можно объяснить синтетической обработкой.
, 1997; Silbering and Galizia, 2007; Silbering et al., 2008; Deisig et al., 2010). Более того, ассоциативное обучение запаху изменяет паттерны активности функциональных единиц AL, что увеличивает разницу между смесями и их компонентами (Faber et al., 1999; Daly et al., 2004; Fernandez et al., 2009; Rath et al. , 2011). Для нашего исследования это подразумевает модификацию паттернов активности AL для метки с помощью одного дополнительного компонента (метка A), тем самым отделяя его от шаблона активности для метки с тем же самым и добавленным другим компонентом (метка AB).Наш результат о том, что этикетка с одним дополнительным компонентом является «странной» только для тех рабочих, которые узнали этикетку с двумя дополнительными компонентами как действительную, также можно объяснить синтетической обработкой.
Эта структура для механизма формирования шаблона может также объяснить способность рабочих различать несколько связанных с задачей меток из одной и той же колонии (Bonavita-Cougourdan et al. , 1993; Greene and Gordon, 2003), если предположить, что разные метки, связанные с задачами, будут кодироваться разными MBON.
, 1993; Greene and Gordon, 2003), если предположить, что разные метки, связанные с задачами, будут кодироваться разными MBON.
Значение для организации колонии
Предложенный нами механизм множественного шаблона для распознавания сородичей меняет парадигму того, какая информация о членах колонии собирается и как она используется. Согласно предыдущей гипотезе (система с одним шаблоном) членство сначала распознается, а затем обрабатывается дополнительная информация, такая как сигналы о недавнем выполнении задачи. Вместо этого наши данные предполагают, что признаки распознавания членов колонии могут быть восприняты и классифицированы как принадлежащие разным сущностям, а различные метки могут быть приписаны независимо более чем с одним атрибутом e.g., «товарищ по гнезду и собиратель» или «товарищ по гнезду и няня». Такая разделенная система распознавания соплеменников очень избирательна для разных этикеток, оставаясь при этом достаточно гибкой, чтобы учитывать возникающие различия, например, из-за вовлеченности в задачу, сезонности или диеты.
Поскольку предлагаемые ассоциации в системе с несколькими шаблонами происходят по параллельным путям в MB, для обработки требуется меньше времени по сравнению с последовательной категоризацией с дополнительной информацией, такой как участие задачи в системе с одним шаблоном.Рабочие могут очень быстро распознать сокамерников (Stroeymeyt et al., 2010), и, наоборот, быстрые реакции против других, вероятно, выбраны потому, что отсроченная реакция увеличивает риск получения травмы, и первый, кто отреагирует, вероятно, будет иметь преимущество в любой последующий конфликт. Поскольку индивидуальный опыт формирует шаблоны, мы ожидаем, что формирование шаблона – это индивидуальный процесс, и члены колонии, вероятно, имеют разные шаблоны даже для одной и той же метки. Межиндивидуальные различия в распознавании сородичей обсуждались как один из потенциальных механизмов надежной защитной реакции коллектива (Esponda and Gordon, 2015).
Еще одним следствием индивидуального подкрепления во время формирования и обслуживания шаблона является то, что частые взаимодействия внутри группы задач могут соответствующим образом корректировать шаблоны. Например, работники, участвующие в уходе за выводком, больше взаимодействуют с медсестрами и, таким образом, могут настраивать свой соответствующий шаблон для «соседа по гнезду и медсестры» больше, чем рабочие, которые заняты другой задачей в другом месте. Это может привести к зависящему от опыта улучшению обнаружения дополнительных сигналов в метке, которые характеризуют группу задач.В этом сценарии медсестры больше знают о медсестрах, работники-фуражиры больше о фуражирах, охранники, вероятно, больше о фуражирах и так далее. Действительно, рабочие демонстрируют пространственную привязку внутри гнезда (Mersch et al., 2013; Tschinkel and Hanley, 2017), и, хотя распределение задач является гибким, рабочие, как правило, повторно выполняют одну и ту же задачу в течение более длительных периодов времени (Beshers and Fewell, 2001; Дженсон и Вайденмюллер, 2014). Разделенная система распознавания соплеменников с несколькими узкоспециализированными шаблонами может способствовать распределению задач, когда различные сенсорные ощущения влияют на будущие решения о выполнении задач (Sendova-Franks and Franks, 1995; Pamminger et al., 2014).
Например, работники, участвующие в уходе за выводком, больше взаимодействуют с медсестрами и, таким образом, могут настраивать свой соответствующий шаблон для «соседа по гнезду и медсестры» больше, чем рабочие, которые заняты другой задачей в другом месте. Это может привести к зависящему от опыта улучшению обнаружения дополнительных сигналов в метке, которые характеризуют группу задач.В этом сценарии медсестры больше знают о медсестрах, работники-фуражиры больше о фуражирах, охранники, вероятно, больше о фуражирах и так далее. Действительно, рабочие демонстрируют пространственную привязку внутри гнезда (Mersch et al., 2013; Tschinkel and Hanley, 2017), и, хотя распределение задач является гибким, рабочие, как правило, повторно выполняют одну и ту же задачу в течение более длительных периодов времени (Beshers and Fewell, 2001; Дженсон и Вайденмюллер, 2014). Разделенная система распознавания соплеменников с несколькими узкоспециализированными шаблонами может способствовать распределению задач, когда различные сенсорные ощущения влияют на будущие решения о выполнении задач (Sendova-Franks and Franks, 1995; Pamminger et al., 2014).
Концепция соответствия этикеток и шаблонов
Описание процесса распознавания сородичей в терминах сопоставления метки и шаблона изначально было предназначено для создания концептуальной основы для исследований по распознаванию родственников. Предположение о том, что необходима и реализуется только одна нейронная матрица, вероятно, было стимулировано обнаружением систематических различий в профилях ХГС между колониями и низкой вариабельности внутри колоний. Позже выяснилось, что профили CHC в колонии динамичны во времени, а также существуют различия между членами колонии в соответствии с их функциональными ролями, что ставит под сомнение гипотезу одного шаблона.Однако вопрос о том, могут ли работники использовать несколько шаблонов, никогда ранее систематически не рассматривался. Кроме того, сложность профилей CHC и способность рабочих к быстрой дискриминации привели к исследованиям, в которых основное внимание уделялось возможным механизмам сокращения данных от обнаружения до восприятия, таким как теория включения, а не признанию высокой эффективности и дискриминационных способностей высокоразвитой обонятельной системы. система работников по оценке и категоризации сложных смесей CHC.
Мы полагаем, что использование термина «сопоставление метки и шаблона» все еще оправдано для выделения химического единообразия внутри колоний, учитывая, что все члены колонии могут распознавать запах своей собственной колонии. Однако, когда речь идет о механизмах, лежащих в основе распознавания сокамерников, это упрощение скорее вводит в заблуждение, чем информативно, потому что оно неадекватно описывает разделенную систему распознавания товарищей по гнезду, которая позволяет точно настроить категоризацию химически различных товарищей по гнезду.
Глоссарий
ПрофильCHC – Кутикулярные углеводороды, присутствующие на кутикуле насекомого. Обычно используемые растворители для извлечения этих углеводородов включают неполярные растворители, такие как пентан, гексан, дихлорметан и изооктан.
• Признаки распознавания – компоненты профиля CHC, которые могут быть обнаружены и используются для распознавания сородичей.
• Метка – сумма признаков распознавания в профиле CHC.
• Запах колонии – сумма всех химических компонентов, присутствующих на кутикуле членов колонии.Он включает в себя сигналы распознавания сородичей и другие компоненты, не используемые для распознавания сородичей. Примечание: это часто используемое определение относится к химической среде, а не к восприятию и оценке химических компонентов.
• Шаблон – нейронное представление метки товарища по гнезду.
• Прием – обнаружение химических компонентов (отдушек) с помощью ORN, которые кодируют запахи как изменения во времени / скорости потенциалов действия.
• Паттерн активности – параллельные пути в обонятельной системе, которые передают информацию о стимуле функциональным единицам, таким как клубочки в AL или KC в MB.Активность этих функциональных единиц (ассемблерный код) зависит от стимула и может быть записана в виде паттернов активности с течением времени.
• Репрезентация – паттерн деятельности, которому приписывается ценность (усвоенная или врожденная) и, таким образом, становится значимым. Представление химических раздражителей (одорантов) в мозге позволяет воспринимать запахи.
• Обобщение – химические стимулы, которые можно различить, оцениваются или классифицируются как принадлежащие к общей сущности.
Авторские взносы
СК, MH и SN разработали и разработали эксперименты.MH проводил эксперименты, собирал данные о поведении рабочих и экстракты для анализа ГХ-МС. SN провела формальный анализ поведенческих данных. SN и MH проанализировали данные ГХ-МС. JM синтезировал компоненты 5-MeC27 и 11-MeC27 для исследования. С.Н. подготовил первый вариант рукописи. CK критически пересмотрел его на предмет важного интеллектуального содержания. Окончательный вариант рукописи одобрен всеми авторами.
Финансирование
Эта работа финансировалась Исполнительным агентством ЕВРОПЕЙСКОЙ КОМИССИИ по исследованиям Marie Curie Actions – International Research Staff Exchange Scheme Project No.: 319015 (https://ec.europa.eu/research/mariecurieactions/funded-projects/international-research-staff-exchange-scheme_en для CK и SN). DAAD, PPPAustralien2015-Kleineidam (https://www.daad.de/de/ к CK и SN Projekt 57219808). IMPRS Organismal Biology, Университет Констанца (https://www.orn.mpg.de/2453/Short_portrait, SN Projekt M.R8.A.ORNI0001 J). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим Френци Корнер-Нивергельт за ее помощь в анализе данных и Лохан Валадарес за помощь в идентификации углеводородов и комментарии к рукописи. Мы также благодарим Пола Шишку за плодотворные обсуждения и полезные комментарии к рукописи.
Список литературы
Акино, Т., Ямамура, К., Вакамура, С., и Ямаока, Р. (2004). Прямые поведенческие свидетельства углеводородов в качестве сигналов распознавания гнездового члена у Formica japonica (Hymenoptera: Formicidae). Заявл. Энтомол. Zool. 39, 381–387. DOI: 10.1303 / aez.2004.381
CrossRef Полный текст | Google Scholar
Асо, Ю., Хаттори, Д., Ю., Ю., Джонстон, Р. М., Айер, Н. А., Нго, Т. Т. и др. (2014). Нейронная архитектура грибовидного тела обеспечивает логику ассоциативного обучения. ELife 3: e04577. DOI: 10.7554 / eLife.04577
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Биттерман, М. Э., Менцель, Р., Фиц, А., и Шефер, С.(1983). Классическое кондиционирование удлинения хоботка у медоносных пчел ( Apis mellifera ). J. Comp. Psychol. 97, 107–119. DOI: 10.1037 / 0735-7036.97.2.107
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Bonavita-Cougourdan, A., Clement, J. L., and Lange, C. (1993). Функциональная дискриминация подкасты (собиратели и выводки) у муравья Camponotus vagus Scop .: полиморфизм кутикулярных углеводородных структур. J. Chem. Ecol. 19, 1461–1477. DOI: 10.1007 / bf00984890
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бос, Н., Дрейер, С., Йоргенсен, К. Г., Нильсен, Дж., Герьери, Ф. Дж., И д’Этторр, П. (2012). Сходство обучения и восприятия кутикулярных углеводородов у муравьев. J. Insect Physiol. 58, 138–146. DOI: 10.1016 / j.jinsphys.2011.10.010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брандштеттер, А.С., Эндлер, А.и Kleineidam, C.J. (2008). Распознавание сокамерников у муравьев возможно без тактильного взаимодействия. Naturwissenschaften 95, 601–608. DOI: 10.1007 / s00114-008-0360-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Brandstaetter, A. S., and Kleineidam, C. J. (2011). Распределенное представление социальных запахов указывает на параллельную обработку в антенной доле муравьев. J. Neurophysiol. 106, 2437–2449. DOI: 10.1152 / jn.01106.2010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брандштеттер, А.С., Рёсслер В. и Кляйнейдам К. Дж. (2011). Друзья и враги с точки зрения мозга муравья – нейронные корреляты запахов колонии социального насекомого. PLoS One 6: e21383. DOI: 10.1371 / journal.pone.0021383
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бакли, С. Б. (1866). Описание новых видов североамериканских Formicidae. Proc. Энтомол. Soc. Phila. 6, 152–172.
Кэмпбелл, Р.А.А., Онеггер, К.С., Цинь, Х., Ли В., Демир Э. и Тернер Г. К. (2013). Получение популяционного кода для идентификации запаха в теле гриба Drosophila . J. Neurosci. 33, 10568–10581. DOI: 10.1523 / JNEUROSCI.0682-12.2013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кон, Р., Моранте, И., Рута, В. (2015). Скоординированная и компартментальная нейромодуляция формирует сенсорную обработку у Drosophila . Cell 163, 1742–1755. DOI: 10.1016 / j.ячейка.2015.11.019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коуту А., Алениус М. и Диксон Б. Дж. (2005). Молекулярная, анатомическая и функциональная организация обонятельной системы Drosophila . Curr. Биол. 15, 1535–1547. DOI: 10.1016 / j.cub.2005.07.034
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Крозье Р. Х. и Дикс М. В. (1979). Анализ 2 генетических моделей врожденных компонентов запаха колонии у социальных перепончатокрылых. Behav. Ecol. Sociobiol. 4, 217–224. DOI: 10.1007 / bf00297645
CrossRef Полный текст | Google Scholar
Дейли К. К., Кристенсен Т. А., Лей Х., Смит Б. Х. и Хильдебранд Дж. Г. (2004). Обучение модулирует представления ансамбля запахов в первичных обонятельных сетях. Proc. Natl. Акад. Sci. U S A 101, 10476–10481. DOI: 10.1073 / pnas.0401
1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дэни, Ф. Р., Джонс, Г. Р., Дестри, С., Спенсер, С. Х. и Туриллацци, С. (2001). Расшифровка опознавательной подписи в химическом профиле кутикулы бумажных ос. Anim. Behav. 62, 165–171. DOI: 10.1006 / anbe.2001.1714
CrossRef Полный текст | Google Scholar
Deisig, N., Giurfa, M., and Sandoz, J.C. (2010). Обработка антенных лепестков увеличивает разделимость представлений запаховой смеси у медоносной пчелы. J. Neurophysiol. 103, 2185–2194. DOI: 10.1152 / jn.00342.2009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эррард, К. (1994). Долговременная память, участвующая в распознавании сородичей у муравьев. Anim. Behav. 48, 263–271. DOI: 10.1006 / anbe.1994.1240
CrossRef Полный текст | Google Scholar
Фернандес П. К., Локателли Ф. Ф., Персона-Реннелл Н., Делео Г. и Смит Б. Х. (2009). Ассоциативное кондиционирование настраивает временную динамику ранней обонятельной обработки. J. Neurosci. 29, 10191–10202. DOI: 10.1523 / JNEUROSCI.1874-09.2009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Gadau, J., Heinze, J., Hölldobler, B., and Schmid, M. (1996). Популяция и структура колонии муравья-плотника Camponotus floridanus . Мол. Ecol. 5, 785–792. DOI: 10.1111 / j.1365-294x.1996.tb00374.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гельман А. и Хилл Дж. (2007). Анализ данных с использованием регрессии и многоуровневых / иерархических моделей. Нью-Йорк, Нью-Йорк: Издательство Кембриджского университета.
Google Scholar
Гербер Б., Геберзан Н., Хелльстерн Ф., Кляйн Дж., Ковалкси О., Вюстенберг Д. и др. (1996). Медоносные пчелы передают обонятельные воспоминания, созданные во время посещения цветов, в парадигму расширения хоботка в лаборатории. Anim. Behav. 52, 1079–1085. DOI: 10.1006 / anbe.1996.0255
CrossRef Полный текст | Google Scholar
Джурфа, М. (2007). Поведенческий и нейронный анализ ассоциативного обучения медоносной пчелы: вкус волшебного колодца. J. Comp. Physiol. Нейроэтол. Sens. Neural Behav. Physiol. 193, 801–824. DOI: 10.1007 / s00359-007-0235-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Грин, М. Дж., И Гордон, Д. М. (2007). Структурная сложность сигналов химического распознавания влияет на восприятие принадлежности к группе у муравьев Linephithema humile и Aphaenogaster cockerelli . J. Exp. Биол. 210, 897–905. DOI: 10.1242 / jeb.02706
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Грюневальд, Б.(1999). Физиологические свойства и модуляция ответа нейронов обратной связи грибовидного тела во время обонятельного обучения у медоносной пчелы, Apis mellifera . J. Comp. Physiol. 185, 565–576. DOI: 10.1007 / s0035417
CrossRef Полный текст | Google Scholar
Guerrieri, F. J., Nehring, V., Jorgensen, C.G., Nielsen, J., Galizia, C.G., and d’Ettorre, P. (2009). Муравьи узнают врагов, а не друзей. Proc. Биол. Sci. 276, 2461–2468. DOI: 10.1098 / рспб.2008.1860
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хиге, Т., Асо, Ю., Моди, М. Н., Рубин, Г. М., и Тернер, Г. К. (2015a). Гетеросинаптическая пластичность лежит в основе аверсивного обонятельного обучения у Drosophila . Neuron 88, 985–998. DOI: 10.1016 / j.neuron.2015.11.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хиге Т., Асо Ю., Рубин Г. М. и Тернер Г. К. (2015b). Индивидуализация обонятельного кодирования в выходных нейронах грибовидного тела, обусловленная пластичностью. Nature 526, 258–262. DOI: 10.1038 / nature15396
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хельдоблер Б. и Уилсон Э. О. (1990). Муравьи. Кембридж, Массачусетс: Belknap Press of Harvard University Press.
Холмс, В., и Шерман, П. (1983). Распознавание родства у животных. амер. Sci. 71, 46–55.
Google Scholar
Джинсон, Р., Вайденмюллер, А. (2014). Межиндивидуальная изменчивость социальных насекомых – непосредственные причины и конечные последствия. Biol. Преподобный Камб. Филос. Soc. 89, 671–687. DOI: 10.1111 / brv.12074
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Joerges, J., Kuttner, A., Galizia, C.G., and Menzel, R. (1997). Представления запахов и запаховых смесей визуализируются в мозгу медоносной пчелы. Nature 387, 285–288. DOI: 10.1038 / 387285a0
CrossRef Полный текст | Google Scholar
Юцум А. Р., Сондерс Т. С. и Черретт Дж. М. (1979). Внутривидовая агрессия у муравьев-листорезов Acromyrmex octospinosus . Anim. Behav. 27, 839–844. DOI: 10.1016 / 0003-3472 (79) -6
CrossRef Полный текст | Google Scholar
Кайб М., Эйзерманн Б., Шутерс Э., Биллен Дж., Франке С. и Франк У. (2000). Изменение постфарингеального и кутикулярного углеводородных составов у муравья Myrmicaria eumenoides . J. Comp. Physiol. А 186, 939–948. DOI: 10.1007 / s0035
146
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лахав, С., Сорокер В., Хефец А. и Вандер Меер Р. К. (1999). Прямые поведенческие свидетельства углеводородов как дискриминаторов распознавания муравьев. Naturwissenschaften 86, 246–249. DOI: 10.1007 / s001140050609
CrossRef Полный текст | Google Scholar
Ленуар А., Хефец А., Саймон Т. и Сорокер В. (2001). Сравнительная динамика формирования запаха гештальта у двух видов муравьев Camponotus Fellah и Aphaenogaster senilis (Hymenoptera: Formicidae). Physiol. Энтомол. 26, 275–283. DOI: 10.1046 / j.0307-6962.2001.00244.x
CrossRef Полный текст | Google Scholar
Леонхардт С.Д., Брандштеттер А.С. и Клейнейдам К.Дж. (2007). Процесс реформирования нейронной матрицы для сигналов распознавания товарищей по гнезду у муравья-плотника Camponotus floridanus . J. Comp. Physiol. Нейроэтол. Sens. Neural Behav. Physiol. 193, 993–1000. DOI: 10.1007 / s00359-007-0252-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Локки, К.Х. (1988). Липиды кутикулы насекомых – происхождение, состав и функции. Комп. Biochem. Phys. B 89, 595–645. DOI: 10.1016 / 0305-0491 (88) -7
CrossRef Полный текст | Google Scholar
Мартин, С. Дж., И Драйфхаут, Ф. П. (2009). Реплики товарищей по гнезду и задачи по-разному влияют и кодируются в профилях углеводородов кутикулы муравьев. J. Chem. Ecol. 35, 368–374. DOI: 10.1007 / s10886-009-9612-x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Mauelshagen, J.(1993). Нейронные корреляты парадигм обонятельного обучения в идентифицированном нейроне в мозге медоносной пчелы. J. Neurophysiol. 69, 609–625. DOI: 10.1152 / jn.1993.69.2.609
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мерш Д. П., Креспи А. и Келлер Л. (2013). Отслеживание людей показывает, что пространственная верность является ключевым регулятором социальной организации муравьев. Наука 340, 1090–1093. DOI: 10.1126 / science.1234316
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мескали, М., Провост, Э., Бонавита-Кугурдан, А., и Клемент, Дж. Л. (1995). Поведенческие эффекты экспериментального изменения химической сигнатуры муравья Camponotus vagus (scop). Насекомое. Soc. 42, 347–358. DOI: 10.1007 / bf01242163
CrossRef Полный текст | Google Scholar
Морель Л. и Блюм М. (1988). Распознавание сокамерника у Camponotus floridanus детенышей рабочих муравьев – распознаются ли сестры или соседи по гнезду? Anim. Behav. 36, 718–725.DOI: 10,1016 / s0003-3472 (88) 80154-4
CrossRef Полный текст | Google Scholar
Окада, Р., Рыбак, Дж., Манц, Г., и Мензель, Р. (2007). Связанная с обучением пластичность в PE1 и других внешних нейронах грибовидного тела в мозге медоносной пчелы. J. Neurosci. 27, 11736–11747. DOI: 10.1523 / JNEUROSCI.2216-07.2007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Одзаки, М., Вада-Кацумата, А., Фудзикава, К., Ивасаки, М., Йокохари, Ф., Сатодзи, Ю., и другие. (2005). Распознавание с помощью хемосенсорной сенсиллы муравейника и других, не соплеменников. Наука 309, 311–314. DOI: 10.1126 / science.1105244
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Паммингер Т., Фойцик С., Кауфманн К. К., Шютцлер Н. и Менцель Ф. (2014). Личность рабочего и ее связь с пространственно структурированным разделением труда. PLoS One 9: e79616. DOI: 10.1371 / journal.pone.0079616
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перес-Орив, Дж., Мазор, О., Тернер, Г. К., Кассенаер, С., Уилсон, Р. И., и Лоран, Г. (2002). Колебания и уменьшение представлений запаха в грибовидном теле. Наука 297, 359–365. DOI: 10.1126 / science.1070502
PubMed Аннотация | CrossRef Полный текст | Google Scholar
R Основная команда. (2017). R: язык и среда для статистических вычислений. Р. Фонд статистических вычислений . Вена, Австрия. Доступно в Интернете по адресу: https://www.R-project.org/
Рат, Л., Джованни Галиция, К., Шишка, П. (2011). Множественные следы памяти после ассоциативного обучения в антеннальной доле медоносной пчелы. Eur. J. Neurosci. 34, 352–360. DOI: 10.1111 / j.1460-9568.2011.07753.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рыбак Дж. И Мензель Р. (1993). Анатомия грибовидных тел в мозге медоносной пчелы: нейронные связи α-доли. J. Comp. Neurol. 334, 444–465. DOI: 10.1002 / cne.
0309
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сендова-Франкс, А., и Фрэнкс, Н. (1995). Пространственные отношения в гнездах муравья Leptothorax unifasciatus (лат.) И их значение для разделения труда. Anim. Behav. 50, 121–136. DOI: 10.1006 / anbe.1995.0226
CrossRef Полный текст | Google Scholar
Шарма К. Р., Энцманн Б. Л., Шмидт Ю., Мур Д., Джонс Г. Р., Паркер Дж. И др. (2015). Кутикулярные углеводородные феромоны для социального поведения и их кодирование в антенне муравья. Cell Rep. 12, 1261–1271. DOI: 10.1016 / j.celrep.2015.07.031
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Зильберинг, А. Ф., и Галиция, К. Г. (2007). Обработка смесей запахов в антеннальной доле Drosophila обнаруживает как глобальное ингибирование, так и специфичные для клубочков взаимодействия. J. Neurosci. 27, 11966–11977. DOI: 10.1523 / JNEUROSCI.3099-07.2007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Зильберинг, А.Ф., Окада, Р., Ито, К., и Галиция, К. Г. (2008). Обработка обонятельной информации в антенной доле Drosophila : все идет? J. Neurosci. 28, 13075–13087. DOI: 10.1523 / JNEUROSCI.2973-08.2008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Строймейт, Н., Герьери, Ф. Дж., Ван Цведен, Дж. С., и д’Этторр, П. (2010). Быстрое принятие решений с распознаванием специфических сторон восприятия у муравьев. PLoS One 5: e12377. DOI: 10.1371 / journal.pone.0012377
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Strube-Bloss, M. F., Nawrot, M. P., and Menzel, R. (2011). Выходные нейроны грибовидного тела кодируют ассоциации “запах-вознаграждение”. J. Neurosci. 31, 3129–3140. DOI: 10.1523 / JNEUROSCI.2583-10.2011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Strube-Bloss, M. F., Nawrot, M. P., and Menzel, R. (2016). Нейронные корреляты побочной памяти запаха в выходных нейронах грибовидного тела. Proc. Биол. Sci. 283: 2016 1270. DOI: 10.1098 / rspb.2016.1270
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шишка П., Дитцен М., Галкин А., Галиция К. Г. и Мензель Р. (2005). Редкость и временное обострение обонятельных представлений грибовидных тел медоносных пчел. J. Neurophysiol. 94, 3303–3313. DOI: 10.1152 / jn.00397.2005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чинкель, В.Р., Хэнли Н. (2017). Вертикальная организация разделения труда в гнездах флоридского муравья-комбайна, Pogonomyrmex badius . PLoS One 12: e0188630. DOI: 10.1371 / journal.pone.0188630
PubMed Аннотация | CrossRef Полный текст | Google Scholar
ван Цведен, Дж., Понтьери, Л., и Педерсен, Дж. С. (2014). Статистический подход для определения возможных сигналов для распознавания сородичей. Фронт. Ecol. Evol. 2:73. DOI: 10.3389 / fevo.2014.00073
CrossRef Полный текст | Google Scholar
Вандер Меер, Р. К., Саливанчик, Д., и Лавин, Б. (1989). Временные изменения в структуре углеводородов кутикулы колонии Solenopsis invicta последствия для распознавания сородичей. J. Chem. Ecol. 15, 2115–2125. DOI: 10.1007 / BF01207442
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вагнер Д., Браун, М. Дж. Ф., Браун, П., Куэвас, В., Моисей, Л. Е., Чао, Д. Л. и др.(1998). Рабочие различия в углеводородном составе кутикулы муравьев-комбайнов, Pogonomyrmex barbatus . J. Chem. Ecol. 24, 2021–2037. DOI: 10.1023 / A: 1020781508889
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Witthöft, W. (1967). Absolute anzahl und verteilung der zellen im hirn der honigbiene. Z. Morphol Tiere 61, 160–184. DOI: 10.1007 / bf00298776
CrossRef Полный текст | Google Scholar
Чжао, X., Ленек, Д., Даг, У., Диксон, Б. Дж., И Келеман, К. (2018). Постоянная активность в повторяющемся контуре лежит в основе памяти об ухаживании у Drosophila . eLife 7: e31425. DOI: 10.7554 / eLife.31425
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Зубе, К., Кляйнейдам, К. Дж., Киршнер, С., Ниф, Дж., И Рёсслер, В. (2008). Организация обонятельного пути и обработки запаха в усиковой доле муравья Camponotus floridanus . J. Comp. Neurol. 506, 425–441. DOI: 10.1002 / cne.21548
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мониторинг колебаний цитозольного h3O2, возникающих в результате изменения градиентов плазматической мембраны или митохондриальной активности
Экспрессия производных HyPer и roGFP в
S. pombeМногие генетически закодированные репортеры были разработаны и проанализированы в различных модельных системах, что затрудняет интерпретацию и сравнение результатов.Мы решили создать наши собственные датчики на основе пероксидаз Gpx1 и Tpx1 делящихся дрожжей с двумя основными целями: во-первых, однозначно сравнить чувствительность предыдущих и новых датчиков в одной и той же модельной системе; и во-вторых, чтобы коррелировать определенные внутриклеточные концентрации пероксида, ранее установленные у делящихся дрожжей 11 , с окислением различных цитозольных репортеров.
Сначала мы экспрессировали HyPer 14 , roGFP2 13 и Grx1-roGFP2 21 в цитозоле S.pombe под контролем конститутивного промотора sty1 22 с использованием эписомальных плазмид. Высокая концентрация репортеров позволила нам контролировать окисление зонда непосредственно в экспоненциально растущих культурах, без центрифугирования или переноса среды. Мы применили внеклеточные концентрации пероксидов в диапазоне от 1 до 25 мкМ (предварительная токсичность), от 50 до 100 мкМ (передача сигнала в сторону каскада антиоксидантов Pap1) и от 0,2 до 1 мМ (токсичность, останавливает рост клеток дикого типа), что соответствует внутриклеточные концентрации 3–80 нМ, 0.2–0,3 мкМ и 0,7–3 мкМ соответственно 11 . Как показано на рис. 1a – c и дополнительном рис. 1, три зонда HyPer, roGFP2 и Grx1-roGFP2 окисляются в ответ на обработку H 2 O 2 ; минимальные концентрации внеклеточных пероксидов, запускающих окисление зонда, составляют 50, 25 и 25 мкМ соответственно. HyPer показывает максимальное окисление с внеклеточными 0,2 мМ (рис. 1а).
Рис.1Окисление производных HyPer и roGFP внеклеточным H 2 O 2 .Штамм дикого типа HM123 трансформировали плазмидами p605, p406, p379, p429, p407, p407.C169S и p676 для экспрессии HyPer ( a ), roGFP2 ( b ), Grx1-roGFP2 ( c ). , roGFP2-Gpx1 ( d ), roGFP2-Tpx1 ( e ), roGFP2-Tpx1.C169S ( f ) и roGFP2-Tsa2ΔC R ( g ) соответственно. Указанные внеклеточные (EXT.) Концентрации H 2 O 2 , классифицированные как «токсичные» (способные временно останавливать рост), «сигнальные» (способные запускать каскад антиоксидантного Pap1) или «дококсические» ( неспособный активировать Pap1), были непосредственно добавлены к культурам MM при OD 600 1 в 96-луночных планшетах для визуализации, и рост продолжался при 30 ° C со встряхиванием.Также указаны соответствующие внутриклеточные концентрации пероксида после применения мембранного градиента 300: 1 (INT.). Степень окисления зонда (количество окисленного зонда на 1) указывается на оси Y (OxD roGFP2), за исключением HyPer ( a ), для которого показано соотношение 488/405 нм. Для каждой панели показаны данные из трех биологических повторов с полосами ошибок (SD), отображаемыми на дополнительном рисунке 1
Затем мы слили S. pombe Gpx1 и Tpx1 с roGFP2 и определили, что H 2 O 2 порогов, способных вызвать окисление зонда, составляли 25 и 10 мкМ соответственно (рис.1d, e), едва улучшая значения, полученные с ранее опубликованными репортерами roGFP.
roGFP2-Tpx1.C169S очень чувствителен к добавлению H
2 O 2Как указано во введении, мы недавно установили, что Tpx1 не только чрезвычайно чувствителен к окислению пероксидами, но также очень эффективно перерабатывается Система Trx 11 . Таким образом, мы слили Tpx1.C169S с roGFP2. Этот мутантный белок, в котором отсутствует разрешающий Cys, способен взаимодействовать с H 2 O 2 , но пероксидный Cys48-SOH не может реагировать с разделяющимся Cys с образованием дисульфида.Следовательно, эта мутация устраняет конкуренцию Trx и может усиливать перенос окисления от Cys48-SOH к roGFP2. Как показано на рис. 1f, roGFP2-Tpx1.C169S способен воспринимать внеклеточные концентрации до 1–5 мкМ H 2 O 2 ; эта чувствительность аналогична чувствительности ранее охарактеризованного репортера roGFP2-Tsa2ΔC R , полученного из пероксиредоксина Tsa2 S. cerevisiae , экспрессируемого в делящихся дрожжах (рис. 1g). Если мы рассмотрим градиент межклеточных пероксидов от 300 к 1 11 , внутриклеточная концентрация всего лишь 3–16 нМ H 2 O 2 может отслеживаться с помощью обоих зондов на основе Prx. .Напротив, HyPer, roGFP или Grx1-roGFP могут обнаруживать только концентрации H 2 O 2 , достаточно высокие, чтобы активировать антиоксидантные каскады или даже остановить рост клеточных культур. Как ранее описано in vitro для roGFP2-Tsa2ΔCR, обе химеры на основе Prx также очень чувствительны in vivo к другим окислителям, таким как трет-бутилгидропероксид, менадион (образует супероксид in vivo, который дисмутируется до H 2 O 2 ) и гипохлорита натрия (дополнительный рис.2a), но практически не реагируют или не реагируют на высокие концентрации донора оксида азота, окисленного GSH или дегидроаскорбиновой кислоты (дополнительный рис. 2b). Белковая последовательность roGFP2-Tpx1.C169S представлена на дополнительном рисунке 2c. Этот репортер будет использоваться в дальнейшем для мониторинга его поведения, часто параллельно с совершенно другим сенсором HyPer, в ответ на физиологические, дотоксические волны внутриклеточных пероксидов.
Влияние экспрессии AQP8 на окисление roGFP2-Tpx1.C169S
Мы продемонстрировали, что диффузия H 2 O 2 через мембраны делящихся дрожжей ограничена, и что поглощение в основном Tpx1 увеличивает градиенты до 300: 1 11 .Мы проверили, может ли экспрессия человеческого AQP8 в S. pombe уменьшить эти градиенты, отслеживая окисление цитозольного roGFP2-Tpx1.C169S. Аквапорины (AQP) облегчают диффузию незаряженных и частично полярных молекул, таких как вода или глицерин. Недавние исследования показали, что некоторые AQP млекопитающих, такие как AQP8 23 , также опосредуют транспорт H 2 O 2 через клеточные мембраны человека. Как показано на рис. 2а и дополнительном рис. 3а, экспрессия AQP8 в делящихся дрожжах способствует проникновению H 2 O 2 по крайней мере в пять раз, а окисление зонда может быть обнаружено с помощью всего лишь 0.2–1 мкМ внеклеточные пероксиды. Одновременно экспрессия AQP8 в клетках дикого типа (рис. 2b) или в штамме Δpap1 , лишенном антиоксидантного фактора транскрипции 8 (дополнительный рис. 3b), увеличивает чувствительность к пероксидам на твердых пластинах, не влияя на толерантность к другие стрессоры, такие как хлорид калия. Важно отметить, что экспрессия AQP4, который не передает H 2 O 2 через мембраны, не влияет на толерантность делящихся дрожжей к пероксидам, а окисление зонда не отличается от окисления клеток дикого типа (дополнительный рис.3а, б).
Рис. 2Окисление roGFP2-Tpx1.C169S H 2 O 2 усугубляется экспрессией AQP8 человека. штамм дикого типа PN513 был котрансформирован плазмидой p407.C169S, кодирующей roGFP2-Tpx1.C169S, и пустой плазмидой (слева) или плазмидой p675 для запуска конститутивной экспрессии AQP8. H 2 O 2 обработок были выполнены, и окисление репортера оценили, как описано на рис. 1. Показаны данные из трех биологических повторов с полосами ошибок (S.D.) отображается на дополнительном рис. 3. b Экспрессия AQP8 в делящихся дрожжах снижает устойчивость дикого типа к пероксидам. Серийные разведения MM культур штаммов 972 (WT), AV18 ( Δsty1 ) и 364, несущих pREP.42x (WT + пустая плазмида) или p675 (WT + pAQP8), наносили на чашки с MM-агаром, и указанные концентрации H 2 O 2 или хлорид калия (KCl) и выращивание в течение 3-4 дней при 30 ° C
roGFP2-Tpx1.C169S поглощает H
2 O 2 и влияет на физиологию клетокУ нас есть ранее было показано, что клетки, лишенные Tpx1 или экспрессирующие каталитически неактивный Tpx1.Мутант C48S демонстрирует тяжелые фенотипы для роста в присутствии кислорода 11,20 . Тем не менее, Tpx1.C169S может поддерживать аэробный рост, вероятно, с GSH, поставляющим тиоловую группу для замены Cys169 20 . Чтобы проверить, влияет ли экспрессия зонда roGFP2-Tpx1.C169S на гомеостаз клетки H 2 O 2 , мы трансформировали штамм Δtpx1 эписомальной и интегративной плазмидами, содержащими химерный ген roGFP2-tpx1.C169S . Как показано на дополнительном рис.4, высокие уровни roGFP2-Tpx1.C169S способны в значительной степени подавлять аэробные дефекты роста клеток, лишенных Tpx1. Напротив, более низкие уровни, достигнутые интегративной плазмидой, только частично восстанавливают аэробный рост.
roGFP2-Tpx1.C169S OxD
0 , индикатор H 2 O 2 устойчивые уровниКлетки без Tpx1 демонстрируют дефекты аэробного роста из-за отсутствия основного поглотителя пероксидов во время аэробного роста 20 .Мы стремились изучить, сможет ли новый зонд обнаруживать повышенные стационарные уровни пероксидов на фоне этого штамма. Мы сравнили базальные и внеклеточные коэффициенты окисления, инициируемые H 2 O 2 , обоих семейств зондов, HyPer и roGFP2-Tpx1.C169S. Как показано на рис. 3а и дополнительном рис. 5, уровни базального окисления HyPer, экспрессируемого в клетках, лишенных Tpx1, были идентичны уровням, наблюдаемым в клетках дикого типа. Интересно, что HyPer, экспрессируемый в штамме Δtpx1 , теперь способен воспринимать всего лишь 10-25 мкМ внеклеточной H 2 O 2 .Это отражает тот факт, что поглощение с помощью Tpx1 в настоящее время находится под угрозой, и только 40: 1 градиент внеклеточных перекисей во внутриклеточные применяется на этом фоне штамма только из-за проницаемости. В клетках дикого типа и проницаемость, и очистка H 2 O 2 создают ступенчатый градиент 300 к 1 11 .
Рис. 3Базальные уровни окисления, OxD 0 , roGFP2-Tpx1.C169S повышены в клетках, лишенных Tpx1. Штаммы HM123 (WT) и SG5 ( Δtpx1 ) трансформировали плазмидами p605, кодирующими HyPer ( a ) и p407.C169S, кодирующий roGFP2-Tpx1.C169S ( b ) и H 2 O 2 было проведено лечение, и окисление репортеров оценивалось, как описано на рис. 1. Для каждой панели показаны данные из трех биологических повторов. , с полосами ошибок (SD), отображаемыми на дополнительном рисунке 5
roGFP2-Tpx1.C169S, выраженный в Δtpx1 теперь может определять концентрации пероксидов ниже 1 мкМ, как показано на рисунке 3b (правая панель). Но, что наиболее важно, базальный уровень окисления зонда OxD 0 также значительно изменяется, при этом только 50–60% зонда окисляются на фоне дикого типа (рис.3b, левая панель) и перемещение до 70–80% при деформации Δtpx1 (рис. 3b, правая панель). Важно отметить, что усиление OxD 0 этого зонда в Δtpx1 связано с повышенным содержанием H 2 O 2 , поскольку Grx1-roGFP2, предназначенный для определения окислительно-восстановительных изменений в GSH, не демонстрирует этого увеличения (дополнительный рис. 5в). Мы пришли к выводу, что базальные уровни окисления, OxD 0 , roGFP2-Tpx1.C169S могут использоваться для мониторинга стационарных уровней H 2 O 2 при различных фоновых напряжениях.
Влияние генетических мутаций на окислительно-восстановительное состояние roGFP2-Tpx1.C169S
Мы продемонстрировали, что устойчивые уровни H 2 O 2 , повышенные в Δtpx1 , могут изменить базальное окисление roGFP2-Tpx1.C169S зонд OxD 0 (рис. 3б). Однако установившееся окисление зонда может также зависеть от способности клетки восстанавливать тиол. Мы удалили отдельные гены, кодирующие компоненты систем восстановления тиолов Trx и Grx. Как сообщалось ранее для других производных roGFP 18 , система Grx является основным восстановителем дисульфидной связи roGFP: базовые стационарные уровни окисленного roGFP2-Tpx1.C169S значительно усиливается в клетках, лишенных Pgr1 (GSH редуктаза) или Grx1, основного цитозольного глутаредоксина (рис. 4a и дополнительный рис. 6a).
Рис. 4Влияние генетических мутаций на окисление roGFP2-Tpx1.C169S. Штаммы SB36 ( Δgrx1 ) и AD88 ( Δpgr1 ) ( a ) и штаммы HM123 (WT), SG61 ( Δtrx1 ) и IC76 ( Δtrx1 Δtrx3 ) были трансформированы с помощью p407.C169S и H 2 O 2 обработок и окисление репортера оценивали, как описано на рис.1. c Схема, изображающая роль клеточных доноров электронов в окислении и восстановлении зонда roGFP2-Tpx1.C169S. Блокирование H 2 O 2 детоксикация либо из-за отсутствия рециклинга Tpx1, либо из-за отсутствия активности Ctt1 приводит к накоплению базального или H 2 O 2 -индуцированного окисленного roGFP2.C169S. Переработка roGFP2.C169S зависит от Grx1 за счет GSH и Pgr1. d Штаммы HM123 (WT) и EP160 ( Δctt1 ) трансформировали плазмидой p407.C169S и H 2 O 2 обработок было выполнено, и окисление репортера оценили, как описано на рис. 1. Для каждой панели показаны данные из трех биологических повторов с полосами ошибок (SD), отображенными на дополнительном рис.
Что касается генетических мутаций в цитозольных Trxs, Trx1 и Trx3, ожидается, что они не окажут серьезного влияния на roGFP или на окислительно-восстановительное состояние Tpx1.C169S, поскольку разрешающий Cys пероксидазы отсутствует (см. Введение) 20 .Тем не менее, эти Trx необходимы для рециркуляции Tpx1 24 , и поэтому внеклеточно добавленный H 2 O 2 может не эффективно очищаться в клетках, лишенных обоих Trx. Как показано на фиг. 4b, репортер roGFP2-Tpx1.C169S не подвергается значительному влиянию в базовых условиях у штамма Δtrx1 . Δtrx1 Δtrx3 ячеек показывают OxD 0 0,8, подтверждая, что отсутствие обоих Trx имеет большее влияние на рециклинг Tpx1 24 .Из этих экспериментов мы заключаем, что на базовый уровень окисления roGFP2-Tpx1.C169S могут влиять изменения системы GSH / Grx1 на уровне восстановления зонда, и что она напрямую реагирует на возмущения H 2 O 2 в случае мутантов Trx (рис. 4в).
Затем мы проверили, было ли окисление зонда при внеклеточном H 2 O 2 обратимым. Мы применяли различные концентрации H 2 O 2 к штаммам дикого типа и Δctt1 , лишенным каталазы, и контролировали окисление зонда в течение 7 ч (рис.4г). На фоне дикого типа 2–2,5 часа достаточно для полного восстановления зонда после внесения внеклеточных концентраций пероксидов ниже 100 мкМ (рис. 4d, верхняя панель). Эти кинетики очень похожи в клетках, лишенных Ctt1 (Fig. 4d, нижняя панель). Окисленному зонду в клетках дикого типа требуется более длительное время (3 и 6 часов соответственно), чтобы вернуться к базовым значениям OxD после 0,2 и 1 мМ H 2 O 2 стресса, что согласуется с эффектом остановки этих токсичных концентраций. рост культуры за 1–5 ч 11,22 .Важно отметить, что культуры клеток штамма Δctt1 не могут возобновить рост после 0,2 и 1 мМ внеклеточного стресса, а зонд roGFP2-Tpx1.C169S остается частично (0,2 мМ) или полностью окисленным (1 мМ H 2 O 2 ) больше чем через 7 ч после приложения нагрузки (рис. 4г, нижняя панель). В заключение, зонд roGFP2-Tpx1.C169S является обратимым индикатором внутриклеточных концентраций пероксидов.
roGFP2-Tpx1.C169S может обнаруживать митохондриальную продукцию H
2 O 2Важным источником продукции H 2 O 2 является митохондриальная электронная транспортная цепь (ETC).Мы проверили, способен ли цитозольный roGFP2-Tpx1.C169S воспринимать H 2 O 2 , продуцируемый в митохондриях и диффундирующий из них. Мы обработали клетки антимицином A (ANT), ингибитором комплекса III, который усиливает выработку случайных активных форм кислорода (ROS) в митохондриях из-за накопления восстановленных компонентов ETC выше по течению 25 . Сначала мы отслеживали цитозольное окисление HyPer при добавлении ANT как в штамме дикого типа, так и в клетках, лишенных основного поглотителя H 2 O 2 , Tpx1.Мы представили окисление зонда после обработки ANT черными линиями, и мы также использовали обработки H 2 O 2 в качестве контроля (рис. 5a, b и дополнительный рис. 7). Как показано на рис. 5а, цитозольный HyPer неспособен ощущать продукцию митохондриальных АФК после обработки ANT.
Рис. 5Цитозольный roGFP2-Tpx1.C169S может обнаруживать митохондриальную продукцию H 2 O 2 . Штаммы HM123 (WT) и SG5 ( Δtpx1 ) трансформировали плазмидами p605 ( a ) и p407.C169S ( b ). Указанные концентрации H 2 O 2 и антимицина A (ANT) добавляли непосредственно к культурам MM, и окисление репортеров оценивали, как описано на фиг. 1; Окисление зонда после обработки ANT представлено сплошными (5 мкМ ANT) или пунктирными (1 мкМ ANT) черными линиями. a Показаны данные из двух биологических повторов, с полосами ошибок (S.D.), отображенными на дополнительном рисунке 7a. b Показаны данные двух (WT) или трех ( Δtpx1 ) биологических повторов с планками ошибок (S.D.) отображается на дополнительном рис. 7b. c Штамм Δcoq4 был трансформирован плазмидой p407.C169S и H 2 O 2 Было проведено обработок и окисление репортера оценивалось, как описано на рис. 1. Показаны данные из трех биологических повторов с полосами ошибок. (SD) отображается на дополнительном рисунке. 7c
Затем мы обрабатывали клетки дикого типа, экспрессирующие репортер roGFP2-Tpx1.C169S, с помощью ANT. Небольшое, но значительное увеличение базального окисления зонда наблюдалось через 30–90 мин после воздействия ANT (рис.5б, верхняя панель, черные линии). Стоит отметить, что на скорость роста клеток делящихся дрожжей, выращенных в ММ, не оказывает значительного влияния ANT-зависимое ингибирование ETC, что указывает на то, что дыхание не является движущей силой пролиферации S. pombe при высоких концентрациях глюкозы 26 . Окисление зонда roGFP2-Tpx1.C169S после обработки ANT гораздо более заметно в клетках Δtpx1 , что позволяет предположить, что H 2 O 2 , продуцируемый в митохондриях и диспергированный в цитозоле, быстро поглощается Tpx1 в клетках дикого типа. клетки (рис.5б, нижняя панель).
В соответствии с химическим ингибированием ETC с помощью ANT, генетическая блокировка также вызывает утечку H 2 O 2 из мембраны. Таким образом, штаммы, несущие делеции отдельных генов компонентов ETC, обнаруживают дефекты роста на твердых пластинах MM, которые можно облегчить добавлением антиоксидантов или недостатком кислорода. Мы предположили, что повышенные стационарные уровни H 2 O 2 из-за утечки электронов ставят под угрозу рост этих клеток 27 .Как теперь показано с помощью зонда roGFP2-Tpx1.C169S, стационарные уровни H 2 O 2 в клетках Δcoq4 , в которых отсутствует белок, необходимый для биосинтеза убихинона 27 , вызывают 70-80% окисление зонд. Это подтверждает массивную (и постоянную) утечку H 2 O 2 из митохондрий в цитозоль на фоне этого штамма, вероятно, на уровне комплекса I 28 (рис. 5c).
Влияние истощения глюкозы на roGFP2-Tpx1.C169S окисление
Баланс между дыханием и ферментацией во многом зависит от доступности питательных веществ. В большинстве типов клеток, включая опухоли и дрожжи, рост в богатой питательными веществами среде вызывает ферментацию и подавляет дыхание (эффект Крэбтри 29,30 ). Условия, богатые глюкозой, в разной степени подавляют респираторный метаболизм всех типов клеток. Поскольку производство АФК связано с метаболизмом митохондрий, мы решили определить влияние низкого уровня глюкозы на roGFP2-Tpx1.Базовое окисление C169S, OxD 0 . Таким образом, мы сравнили OxD 0 в стандартном MM (MM-Glu), который содержит 2% глюкозы в качестве единственного источника углерода, с таковым в MM-Gly (1,85% глицерина и 0,15% глюкозы). К нашему удивлению, OxD 0 имеет очень низкий уровень MM-Gly (рис. 6a и дополнительный рис. 8), что позволяет предположить, что уровни внутриклеточного H 2 O 2 ниже в клетках, выращенных в условиях, склонных к респираторным заболеваниям. .
Рис. 6Влияние сред с низким содержанием глюкозы на roGFP2-Tpx1.C169S окисление. Штамм HM123 , трансформированный p407.C169S, выращивали в MM-Glu или MM-Gly (респираторно-предрасположенные условия), проводили обработки H 2 O 2 и оценивали окисление репортера, как описано на фиг. 1. Показаны данные из трех биологических повторов, с полосами ошибок (SD), отображенными на дополнительном рис. 8a. b Схема, изображающая эффект депривации глюкозы в активации и инактивации киназ Sty1 и Pka1 соответственно.Активация пути Sty1 запускает транскрипцию Atf1-зависимых антиоксидантных генов, таких как ctt1 или tpx1 . Инактивация киназы Pka1 приводит к подавлению репрессии глюконеогенных генов и усилению регуляции митохондриального потребления кислорода. Фосфатаза Pyp1 является негативным регулятором киназы Sty1. c Штаммы HM123 (WT), EP16 ( Δsty1 ), EP193 ( Δatf1 ) и ED1150 ( Δcgs1) трансформировали плазмидой p407.Уровни C169S и базального OxD 0 , определенные, как описано на фиг. 1. Показаны данные из трех биологических повторов с планками ошибок (S.D.) и наложенными отдельными точками данных (черные точки). d Штаммы MC22 ( Δpka1 ), MC24 ( Δpka1 Δsty1 ) и AZ64 ( Δpyp1 ) были трансформированы плазмидой p407.C169S и H 2 O 2 обработок репортера были выполнены оценивается, как описано на рис. 1. Показаны данные из трех биологических повторов с планками погрешностей (S.D.) показан на дополнительном рисунке. 8b
Все типы клеток напрямую реагируют на доступность глюкозы, активируя сигнальные каскады. В S. pombe программы экспрессии генов, зависимых от Pka1 и Sty1, запускаются при истощении запасов глюкозы и являются драйверами этой глюкозозависимой катаболитной репрессии (рис. 6b). Киназа Pka1 специфически инактивируется в условиях низкого уровня глюкозы, а нижестоящий фактор транскрипции Rst2 активирует, среди прочего, глюконеогенный ген fbp1 и гены, связанные с дыхательными путями.Удаление pka1 вызывает дерепрессию fbp1 31 и высокую частоту дыхания, как определено с потреблением кислорода 32 , даже в присутствии повышенных уровней глюкозы. Киназа Sty1 и фактор транскрипции Atf1 активируются не только лишением глюкозы, но и многими другими сигналами окружающей среды, и запускают массивную антистрессовую генную программу, которая включает антиоксидантную активность 33 .
Конечные установившиеся уровни АФК зависят от равновесия между их синтезом и улавливанием.Мы проверили, может ли активация каскада антиоксидантов Sty1-Atf1 за счет истощения глюкозы вызвать снижение базальных уровней H 2 O 2 , что объясняет низкий OxD 0 roGFP2-Tpx1.C169S в клетках, выращенных в среде, склонной к респираторным заболеваниям. . Мы определили OxD 0 штаммов, лишенных компонентов глюкозо-чувствительных путей Pka1 и Sty1, выращенных в стандартном MM (MM-Glu) или в MM-Gly. В отличие от клеток дикого типа, штаммы, лишенные Atf1, демонстрируют очень похожие уровни OxD 0 в обеих средах (рис.6c), в то время как клетки, лишенные Cgs1, ингибитора киназы Pka1, ведут себя как клетки дикого типа. Более того, клетки, лишенные Pka1, обнаруживают конститутивную активацию каскада Sty1-Atf1 посредством неизвестного механизма 32 . При этом базальный уровень окисления зонда в MM-Glu значительно ниже в клетках, лишенных Pka1 (рис. 6d, левая панель OxD 0 : 0,26), чем в клетках дикого типа (рис. 6a, левая панель; OxD 0 : 0,55) в зависимости от Sty1 (рис. 6d, центральная панель, OxD 0 в Δpka1 Δsty1 ячейках: 0.6). Эти результаты предполагают, что активация антиоксидантного ответа Sty1-Atf1 вызывает низкие уровни внутриклеточных пероксидов штамма Δpka1 . Действительно, конститутивной активации только пути MAP-киназы Sty1 путем делеции его основной фосфатазы Pyp1 достаточно для запуска низких уровней окисления репортера roGFP2-Tpx1.C169S в клетках, выращенных в MM-Glu (рис. 6d, правая панель). , OxD 0 в Δpyp1 ячейках: 0,3).
Мониторинг митохондриальной активности с помощью roGFP2-Tpx1.C169S
Как объяснено выше, делящиеся дрожжи, выращенные при низких концентрациях глюкозы, смещаются в сторону учащенного дыхания, хотя катаболитная репрессия не так драматична, как у других микробов 27,34 . Недавно сообщалось, что глюкозозависимая репрессия более эффективна в стандартной среде YE, чем в MM, поскольку потребление кислорода и ANT-зависимое подавление роста ниже в YE 27,35 .
Мы провели поиск питательной среды на основе YE, изменяющей митохондриальную активность в различной степени.Мы определили уровни потребления кислорода культур дикого типа, выращенных в стандартной среде, богатой YE, содержащей 3% глюкозы, YE-0,08% глюкозы (эта концентрация глюкозы в MM запускает сдвиг ферментации в дыхание у делящихся дрожжей, как красиво описано. Янагида и его коллеги 26 ) и YE-3% глицерин (недавно было описано, что эта среда вызывает экстремальную реакцию углеродного голодания на уровне транскрипции 36 ). Как показано на фиг. 7a, потребление кислорода при той же плотности клеток значительно ниже в культурах YE-3% глюкозы, будучи максимальным для YE-3% глицерина и промежуточным в YE-0.Культуры клеток с глюкозой 08%. Кроме того, присутствие ANT в значительной степени препятствует росту делящихся дрожжей в YE-0,08% глюкозы и YE-3% глицерина, но не в YE-3% глюкозе (дополнительный рис. 9a).
Рис. 7roGFP2-Tpx1.C169S Окисление ANT пропорционально митохондриальной активности. a Потребление кислорода клеток S. pombe , выращенных в различных средах для выращивания. Штамм 972 дикого типа выращивали на среде YE с 3% глицерина (YE-3% Gly), 0,08% глюкозы (YE-0.08% Glu) или 3% глюкозы (YE-3% Glu), и когда культуры достигли OD 600 0,5, регистрировали потребление кислорода (см. Методы). Показаны данные из трех биологических повторов с наложенными планками ошибок (S.D.) и отдельными точками данных (черные точки). b , c Штаммы HM123 (WT) ( b ) и SG5 ( Δtpx1 ) ( c ), трансформированные плазмидой p407.C169S, выращивали в среде YE, содержащей 3% глицерина (YE-3% Gly – зеленые кружки), 0,08% глюкозы (YE-0.08% Glu – красные ромбы) или 3% глюкозы (YE-3% Glu – желтые треугольники). При OD 600 , равном 1, клетки переносили в фильтрованный MM, и окисление зонда после обработки ANT (5 мкМ) отслеживали в течение указанного времени. Через десять минут после добавления 1 мМ H 2 O 2 использовали для определения 100% окисления зонда в каждом штамме и состоянии, в то время как 0% было исходным OxD 0 для каждого штамма и состояния. Показаны данные из трех биологических повторов с ошибками (S.D.) отображается на дополнительном рис. 9bc. Отображается d окисление roGFP2-Tpx1.C169S после обработки ANT в течение 45 минут по данным панелей ( b ) и ( c ). Показаны данные из трех биологических повторов с полосами ошибок (SD) и наложенными отдельными точками данных (черные точки).
Мы вырастили клетки дикого типа и Δtpx1 клеток, экспрессирующих roGFP2-Tpx1.C169S, в трех средах, и применили ингибитор ETC ANT, чтобы определить, может ли повышенная митохондриальная активность вызывать повышение уровней H 2 O 2 и сопутствующее окисление зонда.Как показано ранее для носителей на основе MM, OxD 0 варьировалось в зависимости от носителя (данные не показаны). Чтобы проверить влияние ANT на окисление зонда в трех средах, мы отслеживали процент окисления для каждого начального уровня OxD 0 . Как показано на рис. 7b – d и дополнительном рис. 9bc, в обоих генетических фонах эффект обработки ANT имеет значительно больший эффект на окисление roGFP2-Tpx1.C169S, когда клетки выращивают в YE-3% глицерине, чем в стандартном YE- 3% глюкозы, с промежуточным окислением в YE-0.08% глюкозы. Мы пришли к выводу, что окисление roGFP2-Tpx1.C169S после добавления ингибиторов ETC может использоваться для считывания митохондриальной активности.
Роль поры перехода митохондриальной проницаемости при сердечных заболеваниях
https://doi.org/10.1016/j.bbabio.2008.12.017Получить права и содержание отличная личность. В нормальных физиологических условиях они синтезируют АТФ для удовлетворения энергетических потребностей бьющегося сердца.Здесь кальций действует как сигнал, чтобы сбалансировать скорость производства АТФ с потребностью в АТФ. Однако, когда сердце перегружено кальцием, особенно когда это сопровождается окислительным стрессом, митохондрии охватывают свою темную сторону и вызывают некротическую гибель клеток миоцитов. Это происходит остро при реперфузионном повреждении и хронически при застойной сердечной недостаточности. Здесь перегрузка кальцием, истощение адениновых нуклеотидов и окислительный стресс объединяют силы, чтобы вызвать открытие неспецифической поры в митохондриальной мембране, известной как поры перехода проницаемости митохондрий (mPTP).Молекулярная природа mPTP остается спорной, но текущие данные указывают на матричный белок, циклофилин-D (CyP-D) и два белка внутренней мембраны, транслоказу адениннуклеотида (ANT) и фосфатный носитель (PiC). Ингибирование открытия mPTP может быть достигнуто с помощью ингибиторов каждого компонента, но лучше всего описано нацеливание CyP-D с циклоспорином A (CsA) и его неиммуносупрессивными аналогами. В моделях на животных ингибирование открытия mPTP либо CsA, либо генетическим устранением CyP-D обеспечивает надежную защиту как от реперфузионного повреждения, так и от застойной сердечной недостаточности.Это подтверждает, что mPTP является многообещающей мишенью для лечения сердечно-сосудистых заболеваний человека. Действительно, первые клинические испытания показали, что лечение CsA улучшает выздоровление после лечения коронарного тромбоза с помощью ангиопластики.Сокращения
ANTАдениннуклеотидная транслоказа
GSK3киназа гликогенсинтазы 3
IPишемическое прекондиционирование
mitoK ATPмитохондриальная АТФ-зависимая калиевая передача каналов
PiCTPмитохондриальный переносчик фосфата
PiCTPмитохондриальный переносчик
пептидил-пролил цис – транс изомераза
ROSреактивные формы кислорода
VDACанионный канал, активированный напряжением
Ключевые слова
переходная пора проницаемости митохондрий
Heart
Ischaemia
Ischaemia
Кальций
Адениннуклеотидтранслоказа
Циклофилин-D
Циклоспорин A
Митохондриальный носитель фосфата
Рекомендуемые статьи Цитирующие статьи (0)
Copyright © 2009 Elsevier B.
