
Как считать ножки на микросхеме — MOREREMONTA
Постараемся ответить на этот вопрос
Нумерация выводов микросхем всегда осуществляется против часовой стрелки начиная с ключа, это касается как отечественных, так и импортных ИМС:
Есть правда низначительный исключения в распиновке, но и то они связаны с конструкцией корпуса, так например у микросхем серии TDA нумерация пинов всегда слева на право:
Части гигантской базы разделенны так, чтобы было удобно записать их на двд диск. Все справочные материалы представлены в формате PDF.
Нумерация выводов микросхем
В данной статье рассказывается как определить нумерацию ножек микросхемы
Когда то я сам столкнулся с таким вопросом когда собирал свою первую схемы с микросхемой на то где первая ножка у нее, где вторая и т.д и как определить где какая ножка.Не всегда такая информация пишется на сайтах и нелегко ее найти.
Нумеруются микросхемы против часовой стрелки начиная с ключа.
Ключ бывает разных видов. На картинке ниже 2 варианта ключа:
1. Полукруглая выемка
бывает еще в виде квадратной или прямоугольной выемки как на нижнем рисунке:
На этом статья наша о определении выводов закончена, у микросхем такие как TDA выводы считаются слева на право от одного и далее
Сегодня мне на почту пришло письмо с просьбой объяснить, как нумеруются ножки у микросхемы.
Нумеруются микросхемы против часовой стрелки начиная с ключа.
Ключ бывает разных видов. На картинке ниже 2 варианта ключа:
1. Полукруглая выемка
бывает еще в виде квадратной или прямоугольной выемки как на нижнем рисунке:
Надеюсь Вам теперь понятно как определять порядок нумерацию ножек микросхемы.
Если будут вопросы, задавайте в комментариях.
6 комментариев к “Нумерация ножек микросхем.
Это все конечно замечательно, но как определить нумерацию ножек микросхемы (A82C250) если ключа нет. Причем в документации на эту микруху ключ показан, а в реальность он отсутствует.
Спасибо, долго искал, наконец-то нашел!
что такое микроконтроллер и как его создать?
Алексей, микроконтролер это мини компьютер, который способен выполнять простые задачи. Микроконтроллер сочетает в себе функции процессора и периферийных устройств.
Создать его сами, Вы наврятли сможете .
Их можно приобрести в магазинах радиоэлектроники. Возьмите парочку ATMega разных и можете взять еще ATTiny 2313.
О микроконтролерах на сайте будет отдельный урок!
Здравствуйте, скажите, у микросхем имеются ножки и они обозначаются короткими символами например Vcc, y — там ещё что то, вот знать бы они у всех стандартные или нет, и что они означают, как вообще ориентироватся, спасибо!
Спасибо. Долго искал про ключ. Догадывался, но хотел убедится))
Как проверить работает ли микросхема или нет
Как проверить работает ли микросхема или нет
Интегральная микросхема — электронная схема произвольной сложности (кристалл), изготовленная на полупроводниковой подложке (пластине или плёнке) и помещённая в неразборный корпус или без такового, в случае вхождения в состав микросборки.
Прежде чем проверять любую микросхему на работоспособность, необходимо знать и понимать ее устройство, хотя бы приблизительно. Это нужно для того, чтобы заранее представлять себе, какие сигналы или напряжения ожидать от исправной микросхемы на ее выводах.
Лучше всего для проверки конкретной микросхемы собрать хотя бы на макетной плате схему для ее тестирования, – это в том случае, если микросхема новая или уже выпаяна.
Вообще, если устройство микросхемы известно, то в некоторых ситуациях ее можно проверить даже не выпаивая с платы, на которой она установлена, просто измерив сигналы на ножках при помощи мультиметра или осциллографа. Тогда наличие или отсутствие сигнала либо искаженная форма импульса сразу покажут, что — к чему.
Внешний осмотр микросхемы
Допустим что микросхема все еще установлена на плате и выпаивать ее сразу нежелательно. Прежде чем подавать питание на плату, внимательно осмотрите микросхему со всех сторон. Быть может есть очевидные физические признаки ее неисправности: трещина на корпусе, обгоревший или отпавший вывод, короткое замыкание между ножками из-за попадания куска провда (и такое бывет), горелые обвесные компоненты и т. д. Если при осмотре никаких поврежднеий не выявлено, можно идти дальше.
Быть может есть очевидные физические признаки ее неисправности: трещина на корпусе, обгоревший или отпавший вывод, короткое замыкание между ножками из-за попадания куска провда (и такое бывет), горелые обвесные компоненты и т. д. Если при осмотре никаких поврежднеий не выявлено, можно идти дальше.
Если к текущему моменту на плату подано питание, то можно аккуратно (с соблюдением техники безопасности!) приступать к дальнейшей проверке микросхемы.
Проверка выводов питания
Первым делом диагностируют цепи питания микросхемы. Это можно проделать при помощи вольтметра (мультиметра). Уточнить выводы питания известной микросхемы очень легко — достаточно заглянуть в документацию (datasheet) на нее. Плюс положительного питания обозначаетя в даташите как VCC+, отрицательное питание VCC-, общий провод имеет обозначение GND.
Итак, минусовой щуп мультиметра устанавливается на общий провод — упирается в минусовой вывод микросхемы, а плюсовой (красный) щуп мультиметра — на соответствующую ножку питания. Если напряжение соответствует норме для микросхемы, значит питание подается как надо, следовательно цепи питания всего устройства исправны.
Если же напряжение питания не в норме, значит необходимо далее проверить саму цепь питания, хотя бы предворительно отпаяв ее от микросхемы. Если цепи питания работают нормально без микросхемы, занчит проблема в микросхеме, и в худшем случае ее действительно придется менять. Если же проблема в цепях питания, значит скорее всего необходимо ремонтировать их (конденсатор, стабилизатор и т. д.).
Проверка источника опорного напряжения
Далее проверяют все известные выводы микросхемы. Например, можно начать с измерения напряжения на выводе встроенного в микросхему источника опорного напряжения Vref, нормальное значение которого указано в документации. На этом выводе должно быть постоянное напряжение определенного значения относительно общего провода. Если оно меньше или сильно больше, занчит внутри микросхемы или в обвесных компонентах что-то не так, и следует продолжить диагностику.
Проверка времязадающих цепей
Если на микросхеме есть какая-нибудь RC-цепь, то на ней, как правило, в рабочем режиме должны наблюдаться пилообразные колебания. На этом этапе опять же полезным будет обратиться к даташиту, чтобы понять где находится данная цепь если она предусмотрена, и на какой ножке должны быть колебания.
Проверка осуществляется осциллографом. Общий его щуп цепляется на минус питания, а измерительный — на соответствующий вывод микросхемы. Если колебания есть и их форма приемлема — все в порядке, можно идти дальше. Если колебаний нет, то скорее всего проблема в микросхеме или в обвесных времязадающих компонентах.
Проверка сигнальных выводов
Наконец, проверяют сигнальные выводы (выходы) микросхемы. Если микросхема управляет каким-то ключом или следующим блоком на схеме, то на соответствующих выходах (или хотя бы на одном выходе, если он единственный) микросхемы должны присутствовать правильные сигналы. Посмотрите в даташите, к каким выводам должны подходить управляемые цепи.
Проверьте осциллографом данные выводы тем же путем, как проверяли RC-цепь. Если сигнал нормальный и значительно не искажен по сравнению с нормальной формой, значит все в порядке. Если сигнал отсутствует или сильно искажен, скорее всего микросхема повреждена, и ее следует заменить, предварительно проверив управляемую цепь, ведь в действительности она может оказаться причиной выхода микросхемы из строя.
Ранее ЭлектроВести писали, что шотландский стартап Gravitricity разработал решение, позволяющее превратить старые, неработающие шахты в гигантские хранилища электроэнергии. Схема новации следующая: когда электростанции начинают вырабатывать больше электроэнергии, чем требуется энергосети, излишки энергии идут на то, чтобы питать лебедки, поднимающие тяжелый груз наверх. Когда накопленную энергию нужно высвободить, груз опускается вниз, попутно вырабатывая электричество.
По материалам: electrik.info.
Формовка и обрубка выводов микросхем
В статье рассказывается об особенностях формовки и обрубки выводов микросхем при их монтаже на печатные платы, даны рекомендации специалистам. Рассмотрены возможности современного оборудования для формовки выводов микросхем на примере предназначенной для этого универсальной установки компании Fancort.
Рассмотрены возможности современного оборудования для формовки выводов микросхем на примере предназначенной для этого универсальной установки компании Fancort.
За последние десять лет количество типов корпусов экспоненциально выросло, возросли и требования к точности и качеству формовки выводов проектируемых и собираемых SMD-сборок с различным набором компонентов. Задача усложняется определенным набором требований: коммерческие требования в корне отличаются от требований военной промышленности, одни производители собирают десятки и сотни тысяч устройств, другие — всего несколько. Одни и те же компоненты могут монтироваться как вручную, так и автоматически, а размер партии может значительно варьироваться.
Еще не так давно инструменты для формовки производились под каждую конфигурацию компонента. Все эти дорогостоящие комплекты оснастки подходили для серийного производства, однако совершенно не годились для мелкосерийного выпуска, например, для производства демонстрационных образцов, инженерного проектирования, космических и исследовательских программ (где количество образцов в партии могло быть меньше двух).
Решением проблемы стало создание универсальных односторонних штампов. Универсальный штамп выглядит как стандартный формовочный, только с формовкой по одной стороне (см. рис. 1). В отличие от специализированных штампов, которые обрубают и формуют выводы за одну операцию, универсальное устройство имеет одно место для обрубки выводов заданной длины, а другое — для формовки выводов. Более того, сменные вставки позволили использовать различные конфигурации формовки компонентов.
Рис. 1. Универсальный формовочный штамп имеет заменяемые вставки и микрометры, обеспечивающие гибкость настройки выводов |
Типы расположения выводов
Производители выпускают корпуса с тремя различными расположениями выводов: сверху, сбоку и снизу, как это показано на рисунке 2. Часто из-за этого возникают проблемы при использовании корпусов сразу с несколькими вариантами расположения выводов. Каждый вариант имеет свои особенности проектирования и свой способ формовки выводов. Следует отметить, что корпуса с верхним расположением выводов в большей степени подвержены эффекту остаточных упругих деформаций, чем корпуса с нижним и боковым расположениями, что связано с большей длиной вывода. Хотя этот фактор и не критичен при выборе корпуса, но в сочетании с толщиной выводов и высотой зазора это дает общее представление о занимаемой компонентом площади на плате и позволяет подобрать соответствующие контактные площадки.
Часто из-за этого возникают проблемы при использовании корпусов сразу с несколькими вариантами расположения выводов. Каждый вариант имеет свои особенности проектирования и свой способ формовки выводов. Следует отметить, что корпуса с верхним расположением выводов в большей степени подвержены эффекту остаточных упругих деформаций, чем корпуса с нижним и боковым расположениями, что связано с большей длиной вывода. Хотя этот фактор и не критичен при выборе корпуса, но в сочетании с толщиной выводов и высотой зазора это дает общее представление о занимаемой компонентом площади на плате и позволяет подобрать соответствующие контактные площадки.
а) | б) | в) |
Рис. 2. Примеры расположения выводов на корпусе: а) выводы сверху; б) выводы сбоку; в) выводы снизу | ||
Форма выводов
Технология монтажа микросхем на печатную плату существенно изменилась в последние годы. В прошлом практически все микросхемы выпускались со штыревыми выводами, которые запаивались в отверстия на печатной плате. Преимуществом этой технологии монтажа является ее простота, т.к. производство таких плат не требует сложного оборудования и специальной подготовки. Недостатком является то, что отверстие занимает на плате много места, и расстояние между соседними выводами микросхемы должно быть существенно больше, чем при использовании технологии поверхностного монтажа (SMT — Surface Mount Technology), когда выводы микросхемы припаиваются к поверхности платы. На рисунке 3 изображены формы выводов, наиболее часто используемые при производстве корпусов.
В прошлом практически все микросхемы выпускались со штыревыми выводами, которые запаивались в отверстия на печатной плате. Преимуществом этой технологии монтажа является ее простота, т.к. производство таких плат не требует сложного оборудования и специальной подготовки. Недостатком является то, что отверстие занимает на плате много места, и расстояние между соседними выводами микросхемы должно быть существенно больше, чем при использовании технологии поверхностного монтажа (SMT — Surface Mount Technology), когда выводы микросхемы припаиваются к поверхности платы. На рисунке 3 изображены формы выводов, наиболее часто используемые при производстве корпусов.
Для SMT-монтажа используются корпуса с двумя основными типами выводов: «крыло чайки» (gullwing) и J-образные. Оба типа выводов имеют свои преимущества. Корпуса с выводами типа «крыло чайки» позволяют производить ручную пайку и обеспечивают более простой контроль паяных соединений. Применение корпусов с выводами типа «J» уменьшает площадь печатной платы. В настоящее время корпуса с выводами «крыло чайки» наиболее востребованы, т.к. они позволяют применять простое производственное оборудование и обеспечивать переход к сверхплотному монтажу.
В настоящее время корпуса с выводами «крыло чайки» наиболее востребованы, т.к. они позволяют применять простое производственное оборудование и обеспечивать переход к сверхплотному монтажу.
Другой разновидностью «крыла чайки» являются выводы с формой «горб верблюда» (сamel hump), которые характеризуются более высокой устойчивостью к тепловому расширению и тепловому удару (см. рис. 4).
Рис. 3. Основные типы формовки выводов 1) «Крыло чайки» — ножки загнуты вниз и наружу; 2) J-образные — ножки загнуты вниз и внутрь под корпус компонента, форма буквы J; 3) L-образные — ножки загнуты вниз и внутрь под корпус компонента, форма буквы L; 4) Плоские выводы — ножки направлены в разные стороны прямо от корпуса; 5) Штыревые, обычно это проникающие выводы, которые обрезаются для поверхностного монтажа |
Рис. 4. Вид выводов с формой «горб верблюда» |
Важные размеры
Пожалуй, главной задачей в процессе формовки выводов является разработка спецификации для каждого компонента. Часто у инженера-разработчика есть только чертежи из рекламных брошюр производителя. Обычно размеры в них указаны в широком диапазоне допусков, и использовать эти данные в качестве параметров рабочей спецификации невозможно. На рисунке 5 показаны основные параметры SMD-компонентов, которые необходимо учитывать.
Часто у инженера-разработчика есть только чертежи из рекламных брошюр производителя. Обычно размеры в них указаны в широком диапазоне допусков, и использовать эти данные в качестве параметров рабочей спецификации невозможно. На рисунке 5 показаны основные параметры SMD-компонентов, которые необходимо учитывать.
Другими, не менее важными размерами, являются шаг выводов (расстояние от центра одного вывода до центра следующего), радиус плеча и пятки и размеры самого корпуса. На начальном этапе большинство параметров можно предварительно рассчитать самостоятельно с помощью простых математических формул и справочника допусков. На рисунке 6 показан образец расчета конфигурации изгиба.
Рис. 5. Схематичное изображение основных параметров выводов |
Рис. 6. Расчет конфигурации выводов |
Основные эффекты, влияющие на форму выводов
В данном разделе рассмотрим основные эффекты, которые могут повлиять на правильность формы выводов. Очень важно правильно задать толщину материала, которая представляет собой общую толщину вывода, включая покрытия, лужение и максимальные допуски, потому что в дальнейшем, при подборе оснастки для формовки, это может стать причиной сглаживания, задирания, упругого распрямления и некопланарности выводов.
Поскольку реальное пространство на печатной плате всегда является наиболее важным параметром, инженеры-разработчики, как правило, проектируют компоненты с минимально возможной занимаемой площадью, что, в свою очередь, влияет непосредственно на длину плеча, которым компонент зажимается перед формовкой. Если область для прижима слишком мала, то результатом может стать изменение ширины от края до края, частичное упругое распрямление, скашивание выводов и даже повреждение герметичности.
Оснастка проектируется для формовки под наибольшую толщину используемых выводов с фиксированным отклонением, поэтому специалисту следует крайне внимательно задавать допуск ширины компонента от края до края. Например, если оператор пытается отформовать компонент с выводами меньшего размера, чем предполагает инструмент, то увеличится степень упругих деформаций, и следовательно, значительно увеличится ширина от края до края. Дополнительный эффект вносит и высота ножки: чем ножка длиннее, тем сильнее видно ее распрямление после снятия нагрузки.
Например, если оператор пытается отформовать компонент с выводами меньшего размера, чем предполагает инструмент, то увеличится степень упругих деформаций, и следовательно, значительно увеличится ширина от края до края. Дополнительный эффект вносит и высота ножки: чем ножка длиннее, тем сильнее видно ее распрямление после снятия нагрузки.
Другой проблемой, возникающей при использовании корпусов различных конструкций, является контроль высоты зазора, т.е. высота компонента над платой. Этот параметр критичен при монтаже корпуса на адгезив, а также для циркуляции воздуха, чистки внутреннего пространства или для визуальной инспекции. Известно, например, что производители металлокерамических корпусов не могут точно контролировать их толщину («С» на рисунке 7), она может варьироваться в диапазоне
3,56…4,20 мм (0,140…0,165”). Любые изменения толщины корпуса влияют на высоту зазора («H» на рисунке 7), поэтому перед формовкой каждый корпус необходимо измерить, и только после этого производится ручная или автоматическая настройка инструмента под заданный зазор.
Рис. 7. Параметры CQFP-корпуса |
Не менее важным параметром является копланарность — отклонение расположения выводов от плоскости печатной платы. Большинство компонентов сегодня монтируются автоматически — требуемое значение копланарности составляет ±0,05 мм (±0,002”). Данный параметр зависит от конструкции корпуса и формовочного инструмента, т.е. если выводы будут расположены не на одном уровне или длина плеча будет слишком короткой, то компонент не будет копланарным, что может привести к браку при последующем монтаже и пайке.
Если процесс лужения выводов выполняется до формовки, то необходимо учесть дополнительные факторы. Толщина вывода увеличивается, а материал поверхности становится значительно мягче, поэтому следует увеличить зазор оснастки и следить, чтобы инструмент не «вспахал» поверхность в процессе загиба выводов. В то же время требуется очень точно проводить лужение, чтобы толщина соответствовала заданным допускам, иначе можно повредить инструмент. Наиболее простое решение — проводить лужение после процессов формовки и обрубки.
Наиболее простое решение — проводить лужение после процессов формовки и обрубки.
Рекомендации при формовке выводов типа «крыло чайки»
1. Допустимое распрямление вывода составляет
0,31…0,41 мм (0,012…0,016”) для ножки высотой 2,54 мм (0,1”). При формовке более длинной ножки эти значения будут выше.
2. Стандартный допуск ширины от края до края считается 0,13 мм (0,005”).
3. Минимальная длина плеча 0,89 мм (0,035”).
4. Минимальная длина лапки от пятки до основания 0,89 мм (0,035”).
5. Для ИК-пайки оплавлением лапка должна быть ровной или слегка приподнятой.
6. Для термокомпрессионной пайки лапка должна быть слегка опущена.
7. Минимальный радиус изгиба составляет 1,5 размера толщины вывода.
Данные размеры даны в соответствии с международным стандартом JEDEC, российским аналогом которого является ГОСТ 29137-91.
Универсальные системы формовки
Рассмотрим возможности существующего оборудования для формовки выводов микросхем на примере универсальной установки формовки и обрубки выводов компании Fancort (см. рис. 8). Компания Fancort является лидером в области формовки компонентов для аэрокосмической промышленности и имеет многолетний опыт проектирования инструментов формовки и обрубки для бесчисленного количества применений. Системы для обрубки и формовки по одной стороне за один раз являются наиболее точными и универсальными для большинства компонентов. Однако двух- и четырехсторонние штампы имеют более высокую производительность. На рисунке 9 представлен эскиз оснастки для двусторонней системы формовки выводов формы «крыла чайки», обеспечивающей максимальную гибкость настройки и высокую производительность благодаря автоматическому контролю зазора.
рис. 8). Компания Fancort является лидером в области формовки компонентов для аэрокосмической промышленности и имеет многолетний опыт проектирования инструментов формовки и обрубки для бесчисленного количества применений. Системы для обрубки и формовки по одной стороне за один раз являются наиболее точными и универсальными для большинства компонентов. Однако двух- и четырехсторонние штампы имеют более высокую производительность. На рисунке 9 представлен эскиз оснастки для двусторонней системы формовки выводов формы «крыла чайки», обеспечивающей максимальную гибкость настройки и высокую производительность благодаря автоматическому контролю зазора.
Рис. 8. Установка двусторонней формовки и обрубки выводов Fancort |
Рис. 9. Иллюстрация оснастки двусторонней системы формовки и обрубки |
Сменные элементы штампа и вставки позволяют настраивать все критичные параметры обрабатываемых корпусов.
Изменение конфигурации формуемых компонентов осуществляется следующим образом:
1. Настройка длины (D): измеряется по цифровому индикатору.
2. Настройка толщины вывода (С) и/или радиуса (H): замена верхних формовщиков.
3. Настройка длины лапки (X): замена нижних ножей, верхних формовщиков и прижимных планок.
4. Настройка длины плеча (B): замена опорных плит и прижимных планок.
5. Настройка радиуса плеча (G): замена опорных плит.
6. Настройка угла лапки: замена нижних ножей и верхних формовщиков.
Заключение
Операции формовки и обрубки выводов микросхем являются очень важными в технологическом процессе изготовления электронных компонентов, поскольку они отличаются высокой трудоемкостью и во многом определяют технические характеристики изделий. На качество формовки влияет множество параметров, но современные методы формовки позволяют в минимальные сроки разработать новую конфигурацию компонента, настроить в соответствии с ней установку и отформовать компоненты. Благодаря этому уменьшается время, необходимое для внедрения новых устройств, и значительно повышается эффективность производства.
Благодаря этому уменьшается время, необходимое для внедрения новых устройств, и значительно повышается эффективность производства.
Выпаиваем микросхемы из плат: распайка деталей паяльником
Выпаивание микросхем с платы – задача нетривиальная, вне зависимости от типа контроллера. Отпаиваешь одну ножку, но пока занимаешься другой, она застывает. Можно отгибать ножки после отпаивания, но снова встает проблема отлома контактов. Возникает вопрос, как выпаять микросхему из платы паяльником? Ответ достаточно прост: использовать знания физики и подручные предметы. Существует ряд вариантов аккуратного снятия микрочипов с платы. Но сначала немного теории.
Микросхемы
Типы микросхем
В настоящее время существует ряд корпусов, но наиболее широко распространены всего два, да и по факту все остальные разновидности являются вариантами двух основных типов:
- DIP – грубо говоря, этот вариант корпуса для внутреннего монтажа, ножки этого контроллера помещаются в отверстия на плате;
- SMD – этот тип микрочипов предназначен для поверхностного монтажа, в этом случае на плате размещаются «пятачки», к которым и припаяны ножки микросхемы.

Каждый вариант обладает своими достоинствами и недостатками. Но в рамках статьи интересны их особенности в плане распайки. Как выпаять микросхему в том или ином корпусе, разберём чуть ниже.
Демонтаж DIP-корпуса
Как уже отмечалось, эта разновидность микросхем отличается монтажом в отверстия на монтажной плате. Это налагает определённые ограничения на процесс её демонтажа. Для того чтобы аккуратно извлечь её ножки из отверстий, нужно удалить из места соединения припой, практически полностью освободив ножки. Нужно отметить, что поочерёдный нагрев и демонтаж отдельного контакта тут не подойдёт, так как, остывая, оставшийся на месте припой будет снова фиксировать микрочип на месте. Поэтому распайка DIP корпуса оптимальна следующими методами:
- Использование подручных средств – для этой цели подойдут иглы от медицинских шприцов или специальные полые трубочки, продающиеся сейчас в магазинах электротехники. Но вариант использования медицинской иглы наиболее дешевый и доступный.
 Для этого нужно подобрать иглу диаметром чуть меньше, чем посадочные гнезда для ножки микрочипа. Затем срезать её заостренную часть надфилем либо просто откусить, после чего напильником сточить сплющенную часть. После этого установив получившуюся полую трубку с ровным срезом на посадочное гнездо, просто нагреть её паяльником, освободив этим ножку чипа;
Для этого нужно подобрать иглу диаметром чуть меньше, чем посадочные гнезда для ножки микрочипа. Затем срезать её заостренную часть надфилем либо просто откусить, после чего напильником сточить сплющенную часть. После этого установив получившуюся полую трубку с ровным срезом на посадочное гнездо, просто нагреть её паяльником, освободив этим ножку чипа; - Второй вариант – это перетягивание припоя с места припайки на медные провода, смоченные флюсом, таким, например, как спиртовая канифоль. Нагреваемый паяльником провод с флюсом постепенно перетягивает на себя припой с места пайки. Этот вариант занимает больше времени, но также достаточно эффективен;
- Использование паяльника с отсосом припоя – в этом случае особых сложностей в демонтаже не предвидится. Главное – контролировать температуру нагрева в зоне контакта, чтобы не повредить плату и саму деталь.
Эти варианты позволят быстро и качественно выпаивать DIP-корпуса с платы.
Важно! Основным требованиям к использованию паяльника в этом случае будет постоянный контроль над давлением и температурой в зоне пайки. Перегрев и излишний нажим может вывести деталь из строя.
Перегрев и излишний нажим может вывести деталь из строя.
Вытягивание припоя
Важно! При использовании иглы медицинского шприца можно упростить задачу по её обрезке, для этого перед обрезкой достаточно прокалить докрасна место среза.
SMD контролёры
Поверхностное крепление корпуса более легко поддаётся демонтажу. В этом случае можно использовать широкое жало паяльника и медный провод с флюсом и отпаивать сразу несколько контактов одновременно. Но есть и более интересные методы распайки:
- Использование металлической полосы или половинки бритвенного лезвия для распределения тепла паяльника на один ряд ножек микросхемы. В этом случае на ряд контактов с одной стороны устанавливается стальная полоска и прогревается жалом до плавки припоя, после чего эта сторона чуть приподнимается над платой. Затем таким же образом плавится припой с другой стороны чипа;
- Использование длинного отрезка медной оплётки с нанесённым на неё флюсом.
 Отрезок укладывается на ножки микросхемы с одной стороны и прогревается паяльником; вытягивая на оплётку припоя, деталь приподнимаем пинцетом. Затем таким же образом убираем припой с другой стороны контроллера;
Отрезок укладывается на ножки микросхемы с одной стороны и прогревается паяльником; вытягивая на оплётку припоя, деталь приподнимаем пинцетом. Затем таким же образом убираем припой с другой стороны контроллера; - Технически интересным вариантом является использование сплавов Розе или Вуда. Капли этого припоя наносятся на контакты и прогреваются, этим снижается температура плавления припоя. Далее припой постепенно прогревается, и микросхема демонтируется;
- Использование фена или паяльной лампы. Для использования этого инструмента на места пайки наносится флюс. После чего поверхность и деталь прогреваются, и пинцетом микросхема снимается с монтажных пятачков.
Нужно отметить, что каждый вариант демонтажа используется в конкретных условиях, главная задача в этом случае – подобрать наиболее оптимальный с точки зрения безопасности вариант и при его использовании не повредить саму деталь или дорожки платы.
Использование фена
Важно! При демонтаже микросхемы важно помнить, что любые детали или узлы на плате имеют свой температурный минимум, его превышение приведёт к выводу микросхемы из строя.
Использование подручных средств и паяльника при монтаже или демонтаже микроконтроллеров вполне оправдано, но требует как минимум наличия навыков работы с паяльником. При их отсутствии стоит предварительно потренироваться на ненужных деталях. Этот процесс позволит приобрести нужный опыт, как отпаять микрочип без повреждений, кроме того выбрать наиболее оптимальный вариант работы с конкретной платой и типом корпуса микросхемы.
Видео
Оцените статью:Микросхема паук круглый (145, 537) (примечание белые ноги) – покупка микросхем паук круглый (145, 537) (примечание белые ноги) в новом или бу состоянии
Фото микросхемы паук круглый (145, 537) (примечание белые ноги)
Стоимость покупки микросхемы паук круглый (145, 537) (примечание белые ноги)
Ед. изм. изм. | Цена нов. | Цена б/у | Корпус | Примечание |
|---|---|---|---|---|
| шт | 14,40 грн. | 14,40 грн. | 14,40 грн. | белые ноги |
Микросхемы серии 537 представляют собой статические ОЗУ, выпускавшиеся по некогда передовой КМОП-технологии. 145-я серия микросхем содержит весьма различные по функциональному назначению радиодетали, например специально разработанные киевским НИИ «Микроприбор» БИС для первых советских калькуляторов, например Б3-02 и Электроника 4-71Б. Для последних впервые был сделан оригинальный корпус на сорок выводов, в простонародье называемый микросхема паук круглый белые ноги.
Конструктивные особенности круглых микросхем паук
Характерными особенностями круглых микросхем паук с белыми ногами можно считать:

Мы производим скупку новых, неликвидных и б/у радиоэлементов по самым выгодным в Украине ценам — звоните, будем рады выслушать каждого!
TDA2030A характеристики, DataSheet, аналоги, цоколевка
Интегральная микросхема TDA2030A, согласно своим техническим характеристикам, предназначена для использования в качестве одноканального усилителя низкой частоты класса AB. Она включает в себя, уже стандартные для таких устройств, системы: защиты от короткого замыкания, перенапряжения, ограничения рассеиваемой мощности и термического отключения.
Создаваемые на её основе УНЧ характеризуются небольшими искажениями и уровнем собственных шумов. Широко применяется в изготовлении электронной начинки для систем типа 2.0 и 2.1 компьютерных акустических колонок, сделанных преимущественно из дерева. Мощность последних у разных производителей варьируется от 9 до 14 Вт.
Цоколевка
Распиновка TDA2030A следующая. Она изготавливается в современном, пластиковом корпусе для микросхем PENTAWAT, с пятью металлическими контактами. Более ранние версии выпускались в упаковке ТО220-5. Если смотреть на лицевую сторону, там где маркировка, то первая и вторая “ножки” – это неинверсный (IN) и инверсный (IN inv) входы соответственно, четверная — выход. Отрицательный полюс источника питания (в однополярных схемах) соединяют с третьим (GND), а положительный с пятым выводом микросхемы (Vcc +).
Третий контакт микросхемы физически соединен с радиатором.
Технические характеристики
В техописании интегральной микросхемы TDA2030A указано, что она способна обеспечить большой выходной ток, при этом иметь достаточно низкую гармонику и перекрестные искажения. Внешние диоды защищают от бросков прямого и обратного перенапряжения. Номинальная выходная мощность на один канал составляет до 18 Вт. Устройство может использоваться как с двуполярным, так и однополярным источником питанием.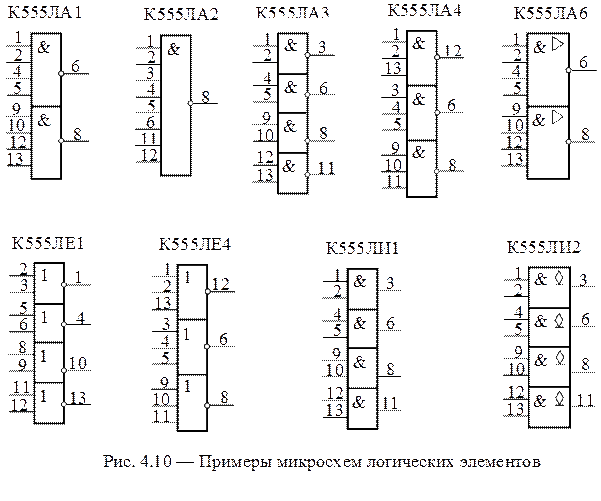 Рассмотрим более подробно его предельно допустимые параметры:
Рассмотрим более подробно его предельно допустимые параметры:
- максимальное напряжение: питания (VS) = ± 22 В; на входе микросхемы (Vi) = ± 22 В; между прямым и инверсным входами (Vdi) = ± 15 В;
- пиковый выходной ток IO = 3,5 А;
- наибольшая мощность рассеивания (при Tк=90 ОС) Ptot = 20 Вт;
- температура хранения и эксплуатации от -40 до +150 ОС.
Максимальное постоянное питающее напряжении TDA2030A может достигать 44 В.
Электрические параметры
Основные электрические характеристики TDA2030A (при VS=±16 В, температуре окружающей среды TA = +25 ОС):
- напряжение питания (VS) от ± 6 до ± 22 В;
- минимальное сопротивлении в нагрузке (RL) — 4 Ом;
- ток покоя (Id) от 50 мА до 80 мА;
- ток смещения на входе Ib (при VS=±22 В) от 0,2 мкА до 2 мкА;
- напряжение смещения на входе Vos (при VS= ±22 В) от ±2 В до ±20 В;
- ток сдвига на входе: от ±20 нА до ±200 нА;
- выходная мощность Po (частота сигнала f от 40 до 15 000 Гц): при RL = 4 Ом — от 15 до 18 Вт; при RL= 8 Ом — от 10 до 12 Вт; при RL= 4 Ом и VS = ± 19 В — от 13 до 16 Вт;
- полоса пропускания BW (при Po = 15 Вт и RL=4) — 100 кГц;
- скорость нарастания SR = 8 В/мкс;
- величина гармонических искажений THD (Po от 0,1 до 14 Вт, f = 40 … 15 000 Гц): при RL= 4 Ом до 0,08%; при RL= 8 Ом до 0,5%;
- отношение сигнал шум: при Po =15 Вт до 106 дБ; Po = 1 Вт до 94 дБ;
- температура отключения при перегреве +145 ОС.
Аналоги
Наиболее подходящими аналогами у TDA2030A являются: LM1875 и TDA2050. Это самая популярная замена у радиолюбителей для ремонта компьютерной акустики. Не стоит путать их с другой микросхемой — TDA2030, которая почти полностью совпадает маркировкой, но не является идентичной и имеет более низкие параметры.
Подобрать похожий операционный усилитель из отечественных образцов не удастся, так как таких просто нет.
Стоит так же отметить модификации рассматриваемой микросхемы с вертикальными (TDA2030AL, TDA2030AV) и горизонтальными выводами (TDA2030AH) для монтажа на плату. Кроме физически измененного расположения контактов, они больше ничем не отличаются от оригинала.
Применение
TDA2030A широко применяются как при создании простейших одномодульных схем, повторителей сигналов, низкочастотных генераторов, так и современных стереоусилителей средней мощности. На видео представлен материал по самостоятельной сборке одного из таких устройств — одноканального УНЧ.
Производители
DataSheet от микросхемы TDA2030A можно посмотреть от ее выпускающих компании: STMicroelectronics, Unisonic Technologies, Contek Microelectronics Co. В нашей стране они широко распространены от STM. Кроме самих микросхем, на прилавках российских магазинов радиотоваров можно встретить готовые модули, с одноименным названием и необходимой обвязкой.
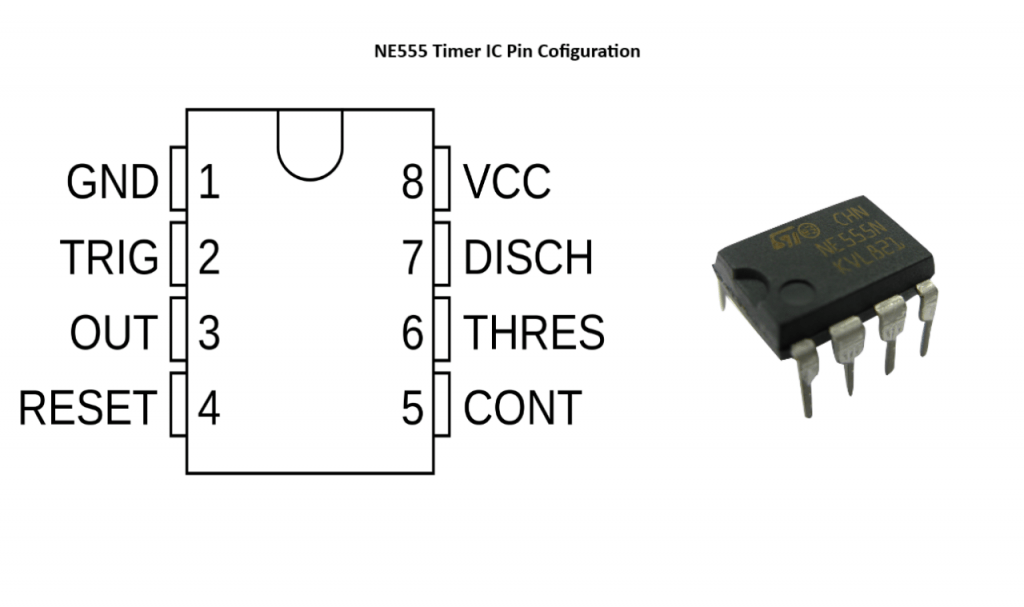
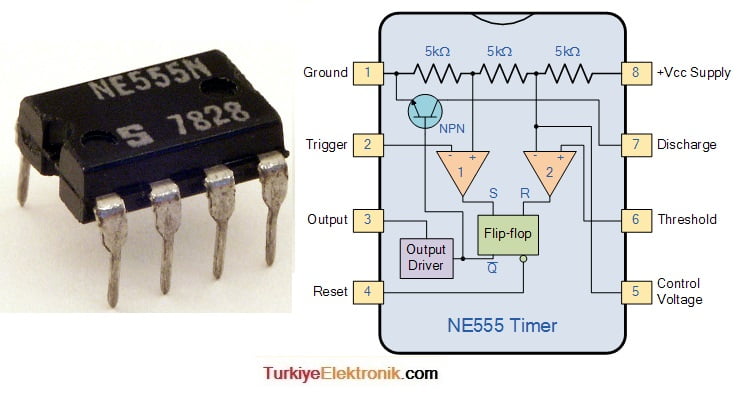
Таймер на включение – выключения в автомобиле NE 555 ?
В автомобиле очень много устройств призванных работать временно, то есть не постоянно а время от времени. Это и различные подогреватели и указатели поворотов (ленивый указатель поворотов) и турботаймеры и устройства включающие камеры заднего хода не сразу, а через какое-то время, то есть с задержкой. Так вот, везде в этих случаях используется таймер, который и задет для исполняющего устройства период его работы или отключения. То есть таймер в машине применяется часто и много где. Мы даже уверены в том, что не все случаи смогли упомянуть и еще несколько вариантов вы можете предложить сами, а может ради них и зашли к нам на страничку. Если это действительно так, то вы здесь как раз и найдете что вам надо, то есть таймер для включения, а равно и отключения исполнительного устройства на машине, в автомобиле.
Так вот, везде в этих случаях используется таймер, который и задет для исполняющего устройства период его работы или отключения. То есть таймер в машине применяется часто и много где. Мы даже уверены в том, что не все случаи смогли упомянуть и еще несколько вариантов вы можете предложить сами, а может ради них и зашли к нам на страничку. Если это действительно так, то вы здесь как раз и найдете что вам надо, то есть таймер для включения, а равно и отключения исполнительного устройства на машине, в автомобиле.
Таймер включения – отключения в автомобиле на микросхеме NE555
Вначале о самой микросхеме, о сердце нашего таймера. Микросхема выпускается а с 70 годов прошлого века и о том, какими компаниями она выпускалась, сколько штук было выпущено уже можно и не вспоминать. Во-первых, это очень значительная информация, а вследствие этого если даже привести статистику, то она будет сильно искажена. Во-вторых, и так понятно, что если микросхема столь востребована, то мы с вами на верном пути, то есть именно эту микросхему целесообразно применять для построения таймера. Здесь кстати стоит отметить, что эта микросхема как раз и задумывалась как таймер, хотя на само деле применяется часто не совсем по назначению, как в одной из наших статьей «Датчик света на микросхеме». Что же, это лишь снова добавляет значимости и плюсиков нашей микросхеме. Теперь о ее подключении и работе схемы.
Здесь кстати стоит отметить, что эта микросхема как раз и задумывалась как таймер, хотя на само деле применяется часто не совсем по назначению, как в одной из наших статьей «Датчик света на микросхеме». Что же, это лишь снова добавляет значимости и плюсиков нашей микросхеме. Теперь о ее подключении и работе схемы.
Схема таймера включения – отключения в автомобиле
Теперь взгляните на классическую схему подключения микросхемы NE555. 1 ножка это земля, 8 это питание «+». Напряжение питания микросхемы 9-12 вольт вполне подойдет. При этом входом микросхемы можно считать ножки 6 и 7, которые соединены между собой, именно на них формируется потенциал от зарядки электролитического конденсатора. В то время, пока конденсатор заряжается, на выходе микросхемы напряжение равно напряжению питания. При этом получается что верхний светодиод не горит, так как для него плюсовое питание осуществляется с двух сторон, а нижний горит из-за разности потенциалов между его ножек. При этом как только электролитический конденсатор заряжается, то потенциал на 3 ножке, на выходе, становится отрицательным, то есть 3 вывод становится землей. В этом случае уже нижний светодиод гаснет, так как для него теперь с двух сторон «минус», а загорается верхний светодиод.
В этом случае уже нижний светодиод гаснет, так как для него теперь с двух сторон «минус», а загорается верхний светодиод.
Вот так работает эта микросхема. Некоторые уже догадались, что заряжается электролитический конденсатор фактически через резистор 1 мОм и 10 кОм, то есть именно от их потенциала, номинала и будет зависеть время зарядки конденсатора, а значит и время срабатывания таймера. В итоге есть два пути изменения время срабатывания таймера. Первый, это изменять номинал резисторов. Второй, изменять емкость конденсатора. Сразу скажем, что изменение емкости конденсатора дает более значимый результат.
А вот весь алгоритм срабатывания таймера реализован в самой микросхеме. Вот собственно и вся схема и принцип ее работы. Осталось лишь сказать, что если вам необходимо управлять большими токами, то здесь как раз и используется сборка на транзисторе (можно взять КТ815Б) и реле 12 вольт, которая так неумело подрисована к рисунку. Само собой реле можно использовать с нормально замкнутым или разомкнутыми контактами, а значит на выходе можно получить включение или отключение. То есть нужным образом коммутировать цепь. Это как раз и будет подтверждать наш заголовок, что микросхема – таймер может обеспечивать как включение, так и отключение каких – либо устройств в автомобиле.
То есть нужным образом коммутировать цепь. Это как раз и будет подтверждать наш заголовок, что микросхема – таймер может обеспечивать как включение, так и отключение каких – либо устройств в автомобиле.
Последнее о чем хотелось сказать, так это о том, что будьте внимательны при монтаже. Подключайте все и вся только проверив все выводы и контакты схемы. Так как микросхема NE 555 сама по себе «нежная», защиты в ней нет, и она просто напросто перегорит. В общем, будьте внимательны и ответственны, тогда у вас все получится!
Видео о работе таймера на микросхеме NE555
Для тех кто не любит читать. ..
.. Видео о работе таймера на микроконтроллере Attiny13
Необходимо сказать об альтернативе сделать таймер на микроконтроллере. В чем-то это весьма лучше! А именно можно легко перенастроить таймер, он не требует конденсаторов и более экономичен. Интегральные схемы– learn.sparkfun.com
Введение
Интегральные схемы (ИС) – краеугольный камень современной электроники. Они сердце и мозг большинства схем. Это вездесущие маленькие черные «микросхемы», которые можно найти практически на каждой печатной плате. Если вы не какой-то сумасшедший мастер аналоговой электроники, у вас, вероятно, будет хотя бы одна микросхема в каждом электронном проекте, который вы создаете, поэтому важно понимать их как внутри, так и снаружи.
Интегральные схемы – это маленькие черные «микросхемы», которые можно найти повсюду во встроенной электронике.
ИС – это набор электронных компонентов – резисторов, транзисторов, конденсаторов и т. Д. – все они помещены в крошечный чип и соединены вместе для достижения общей цели. Они бывают самых разных видов: одноконтурные логические вентили, операционные усилители, таймеры 555, регуляторы напряжения, контроллеры двигателей, микроконтроллеры, микропроцессоры, FPGA … список можно продолжать и продолжать.
Рассмотрено в этом учебном пособии
- Состав IC
- Общие пакеты ИС
- Идентификация ИС
- Часто используемые ИС
Рекомендуемая литература
Интегральные схемы – одна из наиболее фундаментальных концепций электроники.Тем не менее, они основаны на некоторых предыдущих знаниях, поэтому, если вы не знакомы с этими темами, сначала подумайте о прочтении их руководств …
Что такое схема?
Каждый электрический проект начинается со схемы.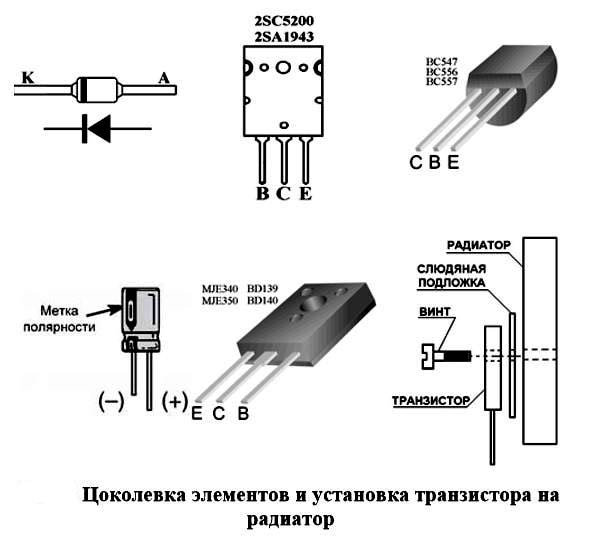 Не знаю, что такое схема? Мы здесь, чтобы помочь.
Не знаю, что такое схема? Мы здесь, чтобы помочь.
Резисторы
Учебник по резисторам. Что такое резистор, как они ведут себя параллельно / последовательно, расшифровка цветовых кодов резисторов и применения резисторов.
Диоды
Праймер диодный! Свойства диодов, типы диодов и их применение.
Полярность
Введение в полярность электронных компонентов. Узнайте, что такое полярность, в каких частях она есть и как ее идентифицировать.
Конденсаторы
Узнайте обо всем, что касается конденсаторов.Как они сделаны. Как они работают. Как они выглядят. Типы конденсаторов. Последовательные / параллельные конденсаторы. Конденсаторные приложения.
Транзисторы
Ускоренный курс биполярных транзисторов. Узнайте, как работают транзисторы и в каких схемах мы их используем.
Узнайте, как работают транзисторы и в каких схемах мы их используем.
Внутри IC
Когда мы думаем об интегральных схемах, на ум приходят маленькие черные микросхемы. Но что внутри этого черного ящика?
Внутренности интегральной схемы, видимые после снятия верхней части.Настоящее «мясо» ИС – это сложное наслоение полупроводниковых пластин, меди и других материалов, которые соединяются между собой, образуя транзисторы, резисторы или другие компоненты в цепи. Вырезанная и сформированная комбинация этих пластин называется штампом .
Обзор кристалла ИС.
Хотя сама ИС крошечная, пластины полупроводника и слои меди, из которых она состоит, невероятно тонкие. Связи между слоями очень сложные.Вот увеличенная часть кубика выше:
Кристалл ИС – это схема в ее наименьшей возможной форме, слишком маленькая для пайки или подключения. Чтобы упростить нам работу по подключению к ИС, мы упаковываем кристалл. Пакет IC превращает тонкий крошечный кристалл в черный чип, с которым мы все знакомы.
Пакет IC превращает тонкий крошечный кристалл в черный чип, с которым мы все знакомы.
Пакеты ИС
Пакет – это то, что инкапсулирует кристалл интегральной схемы и превращает его в устройство, к которому мы можем более легко подключиться. Каждое внешнее соединение на кристалле через крошечный кусочек золотого провода подключается к контактной площадке или контакту на корпусе.Контакты – это серебристые, выдавленные клеммы на ИС, которые используются для подключения к другим частям схемы. Они имеют для нас первостепенное значение, потому что именно они будут подключаться к остальным компонентам и проводам в цепи.
Существует множество различных типов корпусов, каждый из которых имеет уникальные размеры, типы монтажа и / или количество выводов.
Маркировка полярности и нумерация выводов
Все микросхемы поляризованы, и каждый вывод уникален как по расположению, так и по функциям.Это означает, что на упаковке должен быть какой-то способ передать, какой штифт какой. Большинство микросхем будут использовать либо отметку , либо точку , чтобы указать, какой вывод является первым выводом. (Иногда и то, и другое, иногда одно или другое.)
Большинство микросхем будут использовать либо отметку , либо точку , чтобы указать, какой вывод является первым выводом. (Иногда и то, и другое, иногда одно или другое.)
Как только вы узнаете, где находится первый вывод, номера оставшихся выводов последовательно увеличиваются по мере того, как вы перемещаетесь против часовой стрелки по микросхеме.
Тип монтажа
Одной из основных отличительных характеристик типа корпуса является способ его крепления на печатной плате.Все корпуса бывают двух типов: монтаж в сквозное отверстие (PTH) или поверхностный монтаж (SMD или SMT). Пакеты со сквозным отверстием обычно больше, и с ними намного проще работать. Они предназначены для вставки через одну сторону платы и припаивания к другой стороне.
Пакеты для поверхностного монтажа различаются по размеру от маленьких до крохотных. Все они предназначены для размещения на одной стороне печатной платы и припаяны к поверхности. Штыри SMD-корпуса либо выступают со стороны, перпендикулярно чипу, либо иногда располагаются в виде матрицы на дне чипа.ИС в этом форм-факторе не очень удобны для ручной сборки. Обычно для этого требуются специальные инструменты.
Штыри SMD-корпуса либо выступают со стороны, перпендикулярно чипу, либо иногда располагаются в виде матрицы на дне чипа.ИС в этом форм-факторе не очень удобны для ручной сборки. Обычно для этого требуются специальные инструменты.
DIP (двухрядные пакеты)
DIP, сокращение от двухрядного корпуса, является наиболее распространенным корпусом ИС со сквозным отверстием, с которым вы столкнетесь. Эти маленькие микросхемы имеют два параллельных ряда штырей, перпендикулярно выступающих из прямоугольного черного пластикового корпуса.
28-контактный ATmega328 – один из наиболее популярных микроконтроллеров в корпусе DIP (спасибо, Arduino!).Расстояние между контактами DIP IC составляет 0,1 дюйма (2,54 мм), что является стандартным расстоянием и идеально подходит для установки в макетные платы и другие макетные платы. Общие размеры DIP-корпуса зависят от количества контактов, которое может быть от четырех до 64.
Область между каждым рядом контактов идеально разнесена, чтобы позволить микросхемам DIP занимать центральную часть макета. Это обеспечивает каждому контакту свой собственный ряд на плате и гарантирует, что они не замыкаются друг на друга.
Это обеспечивает каждому контакту свой собственный ряд на плате и гарантирует, что они не замыкаются друг на друга.
Помимо использования в макетных платах, микросхемы DIP также могут быть припаяны к печатным платам . Они вставлены в одну сторону платы и припаяны к другой стороне. Иногда, вместо того, чтобы паять микросхему непосредственно на микросхему, рекомендуется установить socket на микросхему. Использование сокетов позволяет снимать и заменять DIP IC, если он «выпустит синий дым».
Обычное гнездо DIP (вверху) и гнездо ZIF с ИС и без нее.Пакеты для поверхностного монтажа (SMD / SMT)
В наши дни существует огромное разнообразие типов корпусов для поверхностного монтажа.Чтобы работать с ИС в корпусе для поверхностного монтажа, вам обычно нужна специальная печатная плата (PCB), изготовленная для них, которая имеет соответствующий узор из меди, на которой они припаяны.
Вот несколько наиболее распространенных типов корпусов SMD, в диапазоне возможностей ручной пайки от «выполнимо» до «выполнимо, но только с помощью специальных инструментов» до «выполнимо только с помощью очень специальных, обычно автоматизированных инструментов».
Small-Outline (СОП)
Малогабаритные ИС (SOIC) – это двоюродный брат DIP для поверхностного монтажа.Это то, что вы получите, если согнете все штыри на DIP наружу и уменьшите его до нужного размера. С твердой рукой и внимательным взглядом эти корпуса являются одними из самых простых для ручной пайки SMD-деталей. В корпусах SOIC каждый штифт обычно отстоит от следующего примерно на 0,05 дюйма (1,27 мм).
SSOP (shrink small-outline package) – это еще меньшая версия пакетов SOIC. Другие похожие пакеты IC включают TSOP (тонкий корпус с мелкими контурами) и TSSOP (корпус с тонкой усадкой и мелкими контурами).
16-канальный мультиплексор (CD74HC4067) в 24-выводном SSOP корпусе.Установлен на плате посередине (четверть добавлена для сравнения размеров).Многие из более простых, ориентированных на одну задачу ИС, таких как MAX232 или мультиплексоры, выпускаются в формах SOIC или SSOP.
Квадратные плоские блоки
Раздвигание выводов микросхемы во всех четырех направлениях дает вам нечто, что может выглядеть как четырехугольный плоский корпус (QFP). ИС QFP могут иметь от восьми контактов на сторону (всего 32) до более семидесяти (всего 300+). Контакты на микросхеме QFP обычно разнесены от 0.От 4 мм до 1 мм. Меньшие варианты стандартного пакета QFP включают тонкий (TQFP), очень тонкий (VQFP) и низкопрофильный (LQFP) пакеты.
ИС QFP могут иметь от восьми контактов на сторону (всего 32) до более семидесяти (всего 300+). Контакты на микросхеме QFP обычно разнесены от 0.От 4 мм до 1 мм. Меньшие варианты стандартного пакета QFP включают тонкий (TQFP), очень тонкий (VQFP) и низкопрофильный (LQFP) пакеты.
Если вы отшлифуете ножки микросхемы QFP, вы получите что-то, что может выглядеть как упаковка с четырьмя плоскими без выводами (QFN) . Соединения на корпусах QFN представляют собой крошечные открытые площадки на нижних угловых краях ИС. Иногда они оборачиваются и обнажены как сбоку, так и снизу, в других упаковках открыта контактная площадка только в нижней части чипа.
Многофункциональный датчик IMU MPU-6050 поставляется в относительно крошечном корпусе QFN с 24 контактами, скрытыми на нижнем крае ИС. Тонкие (TQFN), очень тонкие (VQFN) и микропроводные (MLF) корпуса представляют собой меньшие варианты стандартного корпуса QFN. Существуют даже корпуса с двумя без выводами (DFN) и с двумя тонкими выводами (TDFN), которые имеют контакты только с двух сторон.
Существуют даже корпуса с двумя без выводами (DFN) и с двумя тонкими выводами (TDFN), которые имеют контакты только с двух сторон.
Многие микропроцессоры, датчики и другие современные ИС поставляются в корпусах QFP или QFN. Популярный микроконтроллер ATmega328 предлагается как в корпусе TQFP, так и в форме QFN-типа (MLF), в то время как крошечный акселерометр / гироскоп, такой как MPU-6050, поставляется в миниатюрной форме QFN.
Массивы с шариковой сеткой
Наконец, для действительно продвинутых ИС есть корпуса с шариковой решеткой (BGA). Это удивительно замысловатые маленькие корпусы, в которых маленькие шарики припоя расположены в виде двумерной сетки в нижней части ИС. Иногда шарики припоя прикрепляются непосредственно к матрице!
ПакетыBGA обычно предназначены для продвинутых микропроцессоров, таких как pcDuino или Raspberry Pi.
Если вы умеете паять ИМС в корпусе BGA вручную, считайте себя мастером пайки.Обычно, чтобы поместить эти пакеты на печатную плату, требуется автоматизированная процедура, включающая машины для захвата и размещения и печи оплавления.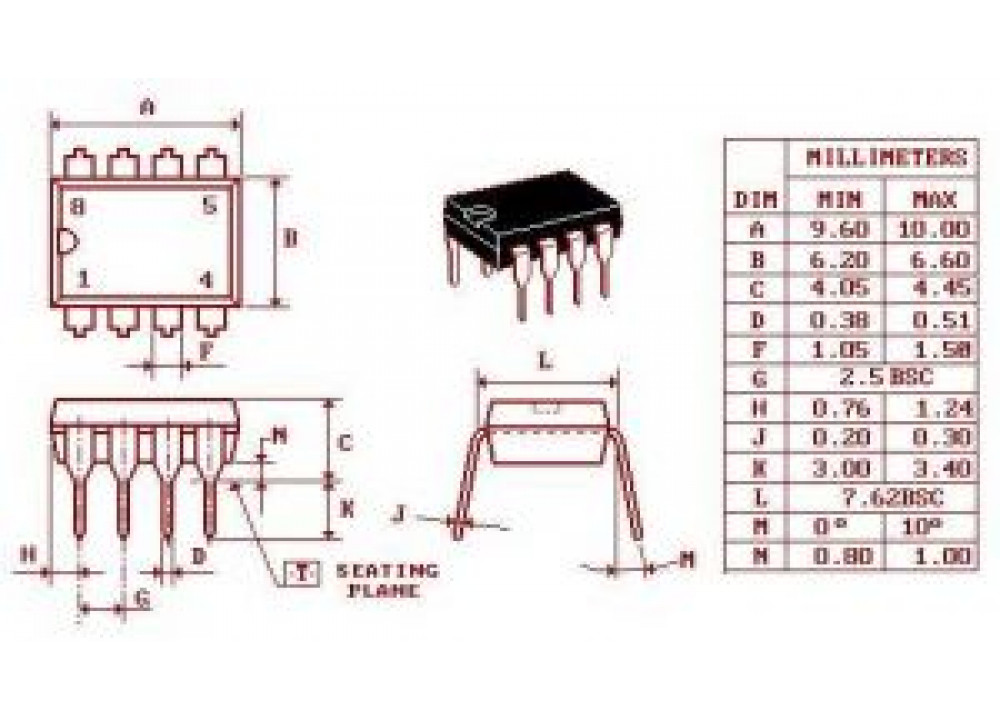
Общие ИС
Интегральные схемы настолько распространены в электронике, что трудно охватить все. Вот несколько наиболее распространенных микросхем, с которыми вы можете столкнуться в образовательной электронике.
Логические вентили, таймеры, регистры сдвига и т. Д.
Логические вентили, являющиеся строительными блоками для гораздо большего числа самих ИС, могут быть объединены в их собственные интегральные схемы.Некоторые ИС логических вентилей могут содержать несколько вентилей в одном корпусе, например этот вентиль И с четырьмя входами:
Логические вентилимогут быть подключены внутри ИС для создания таймеров, счетчиков, защелок, регистров сдвига и других базовых логических схем. Большинство этих простых схем можно найти в пакетах DIP, а также в SOIC и SSOP.
Микроконтроллеры, микропроцессоры, ПЛИС и т. Д.
Микроконтроллеры, микропроцессоры и ПЛИС, содержащие тысячи, миллионы и даже миллиарды транзисторов в крошечной микросхеме, представляют собой интегральные схемы.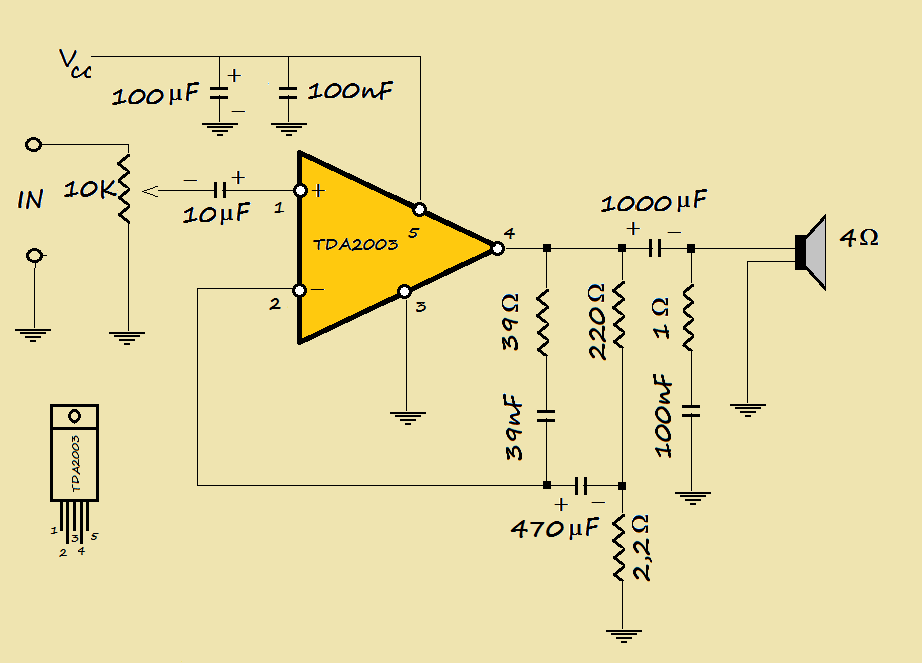 Эти компоненты существуют в широком диапазоне функций, сложности и размеров; от 8-битного микроконтроллера, такого как ATmega328 в Arduino, до сложного 64-битного многоядерного микропроцессора, организующего деятельность на вашем компьютере.
Эти компоненты существуют в широком диапазоне функций, сложности и размеров; от 8-битного микроконтроллера, такого как ATmega328 в Arduino, до сложного 64-битного многоядерного микропроцессора, организующего деятельность на вашем компьютере.
Эти компоненты обычно являются самой большой ИС в цепи. Простые микроконтроллеры можно найти в корпусах от DIP до QFN / QFP, с количеством выводов от восьми до сотни. По мере того, как эти компоненты усложняются, пакет становится одинаково сложным.ПЛИС и сложные микропроцессоры могут иметь до тысячи контактов и доступны только в расширенных пакетах, таких как QFN, LGA или BGA.
Датчики
Современные цифровые датчики, такие как датчики температуры, акселерометры и гироскопы, упакованы в интегральную схему.
Эти микросхемы обычно меньше микроконтроллеров или других микросхем на печатной плате, с числом контактов от трех до двадцати. Микросхемы датчиков DIP становятся редкостью, поскольку современные компоненты обычно встречаются в корпусах QFP, QFN и даже BGA.
Как передвигать ногу, которая когда-то была плавником?
Где-то между 360 и 390 миллионами лет назад группа рыб ожила на суше. Попутно их плавники постепенно трансформировались в опорные конечности с отчетливыми локтевыми и лучезапястными суставами. Плавники превратились в ноги. Пловцы стали ходунками.
Некоторые аспекты этого эволюционного перехода теперь ясны. Мы знаем, как кости этих первопроходцев превратились в те, которые я сейчас использую для печати, благодаря множеству красивых окаменелостей.Мы даже начинаем обнаруживать генетические изменения, которые были связаны с этим преобразованием.
Но загадки остаются. Как, например, хозяева первых ног управляли своими конечностями?
Вытяните руку и согните ее в локте. Происходит несколько вещей: ваши бицепсы сокращаются, но трицепсы также расслабляются. Это происходит из-за модулей в позвоночнике – групп нейронов, которые координируют большое количество движений, которые идут вместе. «Модуль может двигать всю вашу руку, а не только одну пару мышц, или координировать работу мышц руки и туловища», – объясняет Марта Бэгнолл из Северо-Западного университета.
Эти предустановленные программы упрощают управление нашими конечностями. Они похожи на файлы .exe, которые могут запускать сложные компьютерные программы двойным щелчком мыши.
Fish, похоже, не имеет очевидных аналогов для этих модулей. Вместо этого у них есть гораздо более простые нейронные цепи, которые по отдельности управляют левой и правой половинами их тел. Эти цепи срабатывают поочередно, так что мышцы с одной стороны рыбы сокращаются, а мышцы с другой – расслабляются. Это заставляет их тела колебаться из стороны в сторону, толкая их через воду.
Как перейти от этой простой настройки влево-вправо к сложным модулям, которые контролируют ваши собственные конечности? «На самом деле мы не знаем», – говорит Бэгнолл. «Есть отзывы, но они более или менее размахивают руками».
Вместе с коллегой Дэвидом Маклином она нашла важную улику, которая противоречит описанию плавания рыб в учебниках.
Она начала с записи сигналов, которые поступают на двигательные нейроны в мышцах рыбок данио и приказывают им стрелять. Если хрестоматийный взгляд на плавание рыб верен, можно ожидать, что эти сигналы будут приходить одновременно с обеих сторон плывущего животного, поэтому их мышцы могут сокращаться как синхронизированная команда.
Если хрестоматийный взгляд на плавание рыб верен, можно ожидать, что эти сигналы будут приходить одновременно с обеих сторон плывущего животного, поэтому их мышцы могут сокращаться как синхронизированная команда.
Это не то, что происходит. Вместо этого Багнал обнаружил крошечные различия во времени сигналов, достигающих верхней и нижней половины мышц рыбы – всего несколько миллисекунд, но тем не менее различия.
Например, двигательные нейроны в правом верхнем углу рыбы получают синхронизированные сигналы, как и те, которые находятся в ее правом нижнем углу… но эти квадранты синхронизированы друг с другом . Более того, Бэгнолл и Маклин обнаружили, что они получают сигналы от разных наборов нейронов. Есть четыре отдельных цепей, а не две.
При обычном плавании верхний и нижний контуры в основном неразличимы, но они вступают в свои права, когда рыбе нужно катиться. Если рыба данио лежит на правом боку, она посылает более сильные сигналы мотонейронам, контролирующим ее нижний левый и верхний правый квадранты. Возникающие в результате сокращения мышц помогают рыбе выправляться.
Возникающие в результате сокращения мышц помогают рыбе выправляться.
«Мы думаем, что это хороший пример модульной организации спинного мозга рыб», – говорит Багналл. «Они управляют этими четырьмя квадрантами более или менее по отдельности, и это упрощает работу команды, которая производит качение и самовосстановление.
Она предполагает, что эти простые модули могли быть шаблонами для более сложных, управляющих нашими конечностями. Верхние цепи могли развиться для управления мышцами-разгибателями, такими как наши трицепсы, а нижние – для управления сгибателями, такими как наши бицепсы. «На данном этапе это определенно всего лишь гипотеза», – говорит Бэгнолл. «Но поскольку до сих пор не существовало какой-либо правдоподобной гипотезы, мы очень рады этому».
«Это хорошая история, и она красиво изложена», – говорит Оле Кин из Каролинского института, изучающий, как нейроны производят движения.Команде необходимо проработать несколько деталей, например, как отдельные верхний и нижний модули скоординированы в рыбе, но их открытие интересно само по себе, и их гипотеза верна.
Мартин Гулдинг из Института Солка, изучающий схемы позвоночника, более осмотрителен. «Учитывая выдающуюся роль, которую движение конечностей сыграло в переходе позвоночных в наземную среду, этот вопрос является чрезвычайно интересным», – говорит он. «Однако я не уверен, как модульная организация, которую они видят в отношении контроля осевых мышц, легко трансформируется в систему контроля сгибателей и разгибателей.
Например, он отмечает, что мышцы конечностей происходят из другой группы эмбриональных клеток, а не из мышц туловища и позвоночника. Существует поверхностная параллель между верхней и нижней половинами мышц туловища, а также верхней (разгибающей) и нижней (сгибающей) мышцами плавника и конечности, но они могут не совпадать точно.
Багналл соглашается, что для проверки ее гипотезы потребуется гораздо больше работы. Теперь она пытается проверить, полагаются ли нейроны, управляющие верхней и нижней половинами рыбок данио, на разные наборы генов.Если они это сделают, она проверит сгибатели и разгибатели конечностей животных, чтобы увидеть, имеют ли они одинаковые генетические признаки.
Ссылка: Bagnall & McLean. 2013. Модульная организация осевых микросхем в рыбках данио. Наука http://dx.doi.org/10.1126/science.1245629
Как работают интегральные схемы?
Как работают интегральные схемы? – Объясни это РекламаКриса Вудфорда.Последнее изменение: 28 июня 2021 г.
Вы когда-нибудь слышали о компьютере 1940-х годов? называется ENIAC? Он был примерно такой же длины и веса, как три-четыре двухэтажных автобуса. содержал 18 000 гудящих электронных переключателей, известных как вакуумные лампы. Несмотря на свои гигантские размеры, это были тысячи в разы менее мощный, чем современный ноутбук – машина примерно в 100 раз меньше.
Если история вычислительной техники звучит как волшебный трюк – выжать все больше и больше мощности во все меньшее и меньшее пространство – это так! Что сделало это возможно было изобретение интегрированного схема (IC) в 1958 г.Это отличный способ набить сотни, тысячи, миллионы или даже миллиарды электронных компонентов на крошечные чипы кремния нет больше ногтя. Давайте подробнее рассмотрим микросхемы и то, как они работают!
Фото: Интегральная схема снаружи. Он поставляется в удобной форме, называемой двухрядным корпусом (DIP), который состоит из черного пластика или керамический внешний корпус с металлическими штырями по бокам для подключения к электронной плате большего размера (коричневая деталь, которую вы видите на заднем плане).Фактическая схема, которая выполняет эту работу, представляет собой крошечный чип, встроенный в DIP; вы можете увидеть, как он подключен к внешним контактам DIP на следующей фотографии.
Что такое интегральная схема?
Откройте телевизор или радио, и вы увидите, что оно построено вокруг печатная плата (PCB) : немного похожа на электрическую карту улиц с маленький электронный компоненты (например, резисторы и конденсаторы) на месте здания и печатные медные соединения связывая их вместе как миниатюрные металлические улочки.Печатные платы хороши в небольших таких приборов, но если вы попытаетесь использовать ту же технику для построить сложную электронную машину, например компьютер, вы быстро врезался в препятствие. Даже самому простому компьютеру нужно восемь электронных переключает на хранение одного байта (символа) информации. Итак, если вы хотите построить компьютер с достаточным объемом памяти для хранения этого абзаца, вы смотрите примерно 750 символов, умноженных на 8 или около 6000 переключателей – за один абзац! Если вы любите переключатели, как в ENIAC – электронные лампы размером взрослый палец – скоро вы получите колоссально большой, энергоемкая машина, которой нужно собственное мини-электричество завод, чтобы он работал.
Фото: “Электрическая карта улиц”: Интегральная схема (справа) на печатной плате (PCB) с различными обычными электронными компонентами.
Когда в 1947 году три американских физика изобрели транзисторы, несколько улучшилось. Транзисторы были размером с электронные лампы и реле. (электромагнитные переключатели, которые начали заменять электронные лампы в середина 1940-х годов), потребляли гораздо меньше энергии и были гораздо более надежными. Но все еще оставалась проблема соединить все эти транзисторы вместе в сложных схемах.Даже после того, как были изобретены транзисторы, компьютеры по-прежнему представляли собой спутанную массу проводов.
Фото: Интегральные схемы вставляются в печатные платы (ПП), как зеленая, которую вы видите здесь. Обратите внимание на тонкие дорожки, соединяющие «ножки» (клеммы) двух разных ИС. Другие дорожки связывают ИС с обычными электронными компонентами, такими как резисторы и конденсаторы. Вы можете думать о дорожках как о «улицах», прокладывающих пути между «зданиями», где делаются полезные вещи (сами компоненты).Также существует миниатюрная версия печатной платы внутри интегральной схемы: дорожки создаются в микроскопической форме на поверхности кремниевой пластины.
Интегральные схемы все изменили. Основная идея заключалась в том, чтобы взять полная схема со всеми ее многочисленными компонентами и связями между их, и воссоздать все это в микроскопически крошечной форме на поверхности кусок кремния. Это была удивительно умная идея, и она реализована возможные всевозможные «микроэлектронные» гаджеты, которые мы сейчас принимаем за предоставлено, от цифровых часов и карманные калькуляторы на Луну ракеты и ракеты со встроенной спутниковой навигацией.
Фото: Интегральная схема изнутри. Если бы вы могли снять крышку с типичного микрочипа, такого как тот, что на верхнем фото (а это не очень просто – поверьте, я пробовал!), Вы бы нашли внутри именно это. Интегральная схема – это крошечный квадрат в центре. От него выходят соединения к клеммам (металлические штыри или ножки) по краю. Когда вы подключаете что-либо к одной из этих клемм, вы фактически подключаетесь к самой цепи. Вы можете увидеть рисунок электронных компонентов на поверхности самого чипа.Фото любезно предоставлено Исследовательским центром NASA Glenn Research Center (NASA-GRC).
Закон Мура
Интегральные схемы произвели революцию в электронике и вычислительной технике в 1960-х и 1970-х годах. Первый, инженеры помещали десятки компонентов на микросхему в так называемой маломасштабной интеграции (SSI). Вскоре последовала Medium-Scale Integration (MSI) с сотнями компонентов в области такого же размера. Как и ожидалось, примерно в 1970 году крупномасштабная интеграция (БИС) принесла тысячи компонентов, очень крупномасштабная интеграция (СБИС). дали нам десятки тысяч и миллионы сверхбольших масштабов (ULSI) – и все на микросхемах не больше, чем они был раньше.В 1965 году Гордон Мур из компании Intel, ведущего производителя микросхем, заметил, что количество компонентов на микросхеме удваивалась примерно каждые один-два года. Закон Мура , как он известен, продолжает действовать с тех пор. В интервью The New York Times 50 лет спустя, в 2015 году, Мур выразил свое удивление по поводу того, что закон продолжает действовать: «Первоначальное предсказание заключалось в том, чтобы смотреть на 10 лет, что, по моему мнению, было большой натяжкой. Это исходило примерно из 60 элементов. на интегральной схеме до 60 000 – тысячекратная экстраполяция за 10 лет.Я думал, что это было довольно дико. То, что нечто подобное происходит уже 50 лет, поистине удивительно ».
Диаграмма: Закон Мура: количество транзисторов, упакованных в микрочипы, примерно удваивается каждый год или два за последние пять десятилетий – другими словами, оно растет в геометрической прогрессии. Если вы построите график количества транзисторов (ось y) в зависимости от года выпуска (ось x) для некоторых распространенных микрочипов за последние несколько десятилетий (желтые звезды), вы получите экспоненциальную кривую; вместо этого построив логарифм, вы получите прямую линию.Обратите внимание, что вертикальная ось (y) на этой диаграмме является логарифмической. и (из-за графического программного обеспечения OpenOffice, которое я использовал) горизонтальная ось (x) имеет лишь неопределенно линейную форму. Источник: построено с использованием данных Transistor Count, Wikipedia, сверено с данными из других источников.
Рекламные ссылкиКак изготавливаются интегральные схемы?
Как сделать что-то вроде микросхемы памяти или процессора для компьютера? Все начинается с необработанного химического элемента, такого как кремний, который подвергается химической обработке или легированию для придания ему различных электрических свойств…
Легирование полупроводников
Фото: кремниевая пластина. Фото любезно предоставлено Исследовательским центром NASA Glenn Research Center (NASA-GRC).
Если вы читали наши статьи о диодах и транзисторы, ты будешь знаком с идеей полупроводников . Традиционно люди думали, что материалы можно разделить на две аккуратные категории: которые позволяют электричеству течь через их довольно легко (проводники) и те, что нет (изоляторы). Металлы составляют большую часть проводников, а неметаллы, такие как пластик, дерево и стекло изоляторы.На самом деле все гораздо сложнее, особенно когда речь идет об определенных элементы в середине периодической таблицы (в группах 14 и 15), особенно кремний и германий. Обычно изоляторы, эти элементы могут быть заставить вести себя больше как проводники, если мы добавим небольшое количество примеси к ним в процессе, известном как легирование . Если вы добавите сурьму в кремний, вы дадите ему немного больше электронов, чем он. обычно имеет – и способность проводить электричество. Кремний “легированный” таким образом называется n-типа .Добавляем бор вместо сурьмы и вы удаляете часть электронов кремния, оставляя после себя “дырки” которые работают как «отрицательные электроны», неся положительный электрический ток в обратном порядке. Такой кремний называется p-type . Помещение областей кремния n-типа и p-типа рядом друг с другом создает переходы, в которых электроны ведут себя очень интересным образом – и это как мы создаем электронные компоненты на основе полупроводников, такие как диоды, транзисторы и воспоминания.
Внутри завода по производству микросхем
Фото: Интегральные схемы производятся в безупречно чистых условиях; Рабочие должны носить вот такие «костюмы для кроликов», чтобы они не загрязняли чипы, которые они производят.Это завод Intel по производству пластин в Чандлере, Аризона, США. Фото любезно предоставлено архивом Кэрол М. Хайсмит, Библиотека Конгресса, Отдел эстампов и фотографий.
Процесс создания интегральной схемы начинается с большого монокристалл кремния, имеющий форму длинной сплошной трубки, которая «нарезана салями» на тонкие диски (про габариты компакт-диска) называются вафлями . Пластины разделены на множество одинаковых квадратных или прямоугольных областей, каждая из которых из которых будет составлять один кремниевый чип (иногда называемый микрочип).Тогда тысячи, миллионы или миллиарды компонентов создается на каждом чипе путем легирования различных участков поверхности, чтобы превратить их в Кремний n-типа или p-типа. Допинг осуществляется множеством разных процессы. В одном из них, известном как напыление , ионы легирующего материала стреляют в кремниевую пластину, как пули из пистолет. Другой процесс называется осаждение из паровой фазы включает введение легирующего материала в виде газа и его конденсацию, чтобы атомы примеси создают тонкую пленку на поверхности кремния вафля.Молекулярно-лучевая эпитаксия это гораздо более точная форма осаждения.
Конечно, создание интегральных схем, содержащих сотни, миллионы, или миллиарды компонентов на кремниевом чипе размером с ноготь – это все немного сложнее и запутаннее, чем кажется. Представьте себе хаос даже пятнышко грязи может вызвать, когда вы работаете на микроскопический (а иногда даже наноскопический) масштаб. Вот почему полупроводники производятся в безупречных лабораторных условиях, называемых чистых помещений , где воздух тщательно продувается фильтрованный и рабочие должны входить и выходить через шлюзы в любых защитная одежда.
Как сделать микрочип – краткое описание
Хотя создание микросхемы очень сложно и сложно, на самом деле существует всего шесть отдельных шагов (некоторые из них повторяется более одного раза). Процесс значительно упрощен, вот как работает этот процесс:
- Изготовление пластин: мы выращиваем кристаллы чистого кремния в длинные цилиндры и разрезаем их (как салями) на тонкие пластины, каждая из которых в конечном итоге будет разрезана на множество чипов.
- Маскирование: мы нагреваем пластины, чтобы покрыть их диоксидом кремния, и используем ультрафиолетовый свет (синий), чтобы добавить твердый защитный слой, называемый фоторезистом.
- Травление: мы используем химические вещества для удаления части фоторезиста, создавая своего рода шаблонный узор, показывающий, где нам нужны области кремния n-типа и p-типа.
- Легирование: Мы нагреваем протравленные пластины газами, содержащими примеси, для образования областей кремния n-типа и p-типа. Может последовать дополнительная маскировка и травление.
- Тестирование: длинные металлические соединительные провода проходят от испытательной машины с компьютерным управлением к клеммам на каждой микросхеме. Любые чипы, которые не работают, помечаются и отклоняются.
- Упаковка: Все нормально работающие чипы вырезаны из пластины и упакованы в защитные куски пластика, готовые для использования в компьютерах и другом электронном оборудовании.
Кто изобрел интегральную схему?
Вы, наверное, читали в книгах, что ИС были разработаны совместно Джек Килби (1923–2005) и Роберт Нойс (1927–1990), как если бы эти двое мужчин с радостью сотрудничали в их гениальном изобретении! Фактически, Килби и Нойс пришла в голову самостоятельно, более или менее точно так же время, вызвав яростную битву за права на изобретение, которое был совсем не счастлив.
Как два человека могли изобрести одно и то же в одно и то же время? Легко: Идея интегральных схем ждала воплощения. К середине 1950-х гг. мир (и, в частности, военные) открыли потрясающий потенциал электронных компьютеров, и это ослепляюще для таких провидцев, как Килби и Нойс, было очевидно, что лучший способ сборки и подключения транзисторов в больших количества. Килби работал в Texas Instruments, когда наткнулся на идею он назвал принципом монолитности : пытаясь построить все различные части электронной схемы на кремниевом чипе.12 сентября 1958 года он вручную собрал первую в мире грубую интегральную схему. используя чип из германия (полупроводниковый элемент, подобный кремний) и Texas Instruments подали заявку на патент на идея в следующем году.
Между тем, в другой компании под названием Fairchild Semiconductor (образованной небольшая группа сотрудников, которые изначально работали над транзистором пионер Уильям Шокли) не менее блестящий Роберт Нойс экспериментировал с миниатюрой схемы его собственные.В 1959 году он использовал серию фотографических и химические методы, известные как планарный процесс (который только что был разработан коллегой Жаном Орни) создать первую практическую интегральную схему, метод, который Fairchild затем попытался патент.
Работа: Snap! Два великих инженера-электрика, Джек Килби и Роберт Нойс, пришли к этой идее почти в одно и то же время в 1959 году. Хотя Килби первым подал свой патент, патент Нойса был выдан раньше.Вот рисунки из их оригинальных патентных заявок. Вы можете видеть, что у нас, по сути, одна и та же идея с электронными компонентами, сформированными из переходов между слоями полупроводников p-типа (синий) и n-типа (красный). Подключения к областям p-типа и n-типа показаны оранжевым и желтым, а базовые слои (подложки) показаны зеленым. Картины любезно предоставлены Управлением по патентам и товарным знакам США с нашей собственной добавленной окраской, чтобы улучшить ясность и подчеркнуть сходство. Вы можете найти ссылки на сами патенты в приведенных ниже ссылках.
Между работой двух мужчин и Техасом было значительное совпадение. Инструменты и Fairchild боролись в судах большую часть 1960-х годов за то, кто действительно разработал интегральную схему. Наконец, в 1969 г. компании согласились поделиться идеей.
Килби и Нойс в настоящее время по праву считаются соавторами возможно, самая важная и далеко идущая технология, разработанная в 20-м веке. век. Оба мужчины были введены в Национальный зал изобретателей Слава (Килби в 1982 году, Нойс в следующем году) и Килби прорыв был также отмечен присуждением половины доли в Нобелевская премия в Физика в 2000 году (как очень великодушно отметил Килби в своей благодарственной речи, Нойс, несомненно, разделил бы приз, если бы он не умер от сердечного приступа десятью годами ранее).
В то время как Килби помнят как блестящего ученого, наследие Нойса имеет большое значение. добавленное измерение. В 1968 году он стал соучредителем компании Intel Electronics. с Gordon Moore (1929–), который продолжил разработку микропроцессора (однокристальный компьютер) в 1974 г. С IBM, Microsoft, Apple и др. компаниям-новаторам, Intel приписывают помощь в создании доступные персональные компьютеры для дома и на работе. Спасибо Нойсу и Килби и блестящих инженеров, которые впоследствии основывались на своей работе, сейчас используется около двух миллиардов компьютеров. во всем мире многие из них встроены в мобильные телефоны, портативные устройства спутниковой навигации и другие электронные устройства.
Рекламные ссылкиУзнать больше
На сайте
На других сайтах
- Хотите узнать больше о пионерах? Посмотрите на эти страницы о Джек Килби, первоначально опубликовано на веб-сайте Texas Instruments или посетите музей Intel, чтобы узнать о Роберте Нойсе, Гордоне Муре и их коллегах. На обоих сайтах есть великолепная коллекция фотографии ранних интегральных схем.
Статьи
- Startup Graphcore берет верх над Nvidia с последней версией A.I. Чип Джереми Кана, Fortune, 15 июля 2020 г. Представлен транзисторный чип на 59 миллиардов долларов!
- Хорошее, плохое и странное: 3 направления закона Мура Сэмюэля К. Мура. IEEE Spectrum, 26 октября 2018 г. Взгляд на недавний технический прогресс в получении новой жизни из старого закона.
- Intel находит следующий шаг закона Мура при 10 нанометрах, автор Рэйчел Кортленд. IEEE Spectrum, 30 декабря 2016 г. Как новый завод по производству микросхем поможет Intel вдохнуть новую жизнь в закон Мура.
- Закон Мура «Закончилось место, технология ищет преемника» Джона Маркова.The New York Times, 4 мая 2016 г. Почему это имеет значение, если производители микросхем больше не могут следовать закону Мура?
- Закон Мура менее важен для индустрии высоких технологий? пользователя Quentin Hardy. Нью-Йорк Таймс. 25 июля 2014 г. Изменение рабочих привычек и появление облачных вычислений меняют ожидания людей от своих компьютеров, а это означает, что закон Мура уже не так важен, как был.
- Замедляет ли конец закона Мура мировую гонку суперкомпьютеров? пользователя Роберт Макмиллан.Wired, 23 июня 2014 г. Суперкомпьютеры не работают быстрее, чем раньше. Неужели закон Мура, наконец, подходит к концу?
- «25 микрочипов, потрясших мир» Брайана Санто. IEEE Spectrum, 1 мая 2009 г. Если вы думаете, что микросхема – это просто микросхема, подумайте еще раз. В этой статье перечислены два десятка классических микросхем, от схем таймера до флэш-памяти и синтезаторов речи до микропроцессоров, которые радикально изменили историю вычислений.
Книги
История
Технологии
Видео
- От песка к кремнию: Intel показывает вам процесс создания микрочипа, начиная с пустыни (с песка, который дает нам кремний) и заканчивая готовым чипом.Довольно изящное видео, но некоторые комментарии или объяснения не пропали бы даром: это видео действительно имеет смысл только в том случае, если вы уже знаете обо всех процессах, которые вам показывают.
Патенты
Один из лучших способов узнать об изобретениях – это прочитать, как сами изобретатели видели и представляли свои собственные идеи; патенты предлагают отличный способ сделать это. Для тех, кто хочет получить более подробную информацию, вот пара ключевых патентов Килби и Нойса, на которые стоит обратить внимание:
- Патент США 3,115,581: Миниатюрная полупроводниковая интегральная схема от Джека С.Kilby, Texas Instruments, подана 6 мая 1959 г. и опубликована 24 декабря 1963 г. Описывает основную идею создания интегральных схем “с использованием только одного материала для всех элементов схемы и ограниченного числа совместимых этапов процесса для их производства”.
- Патент США 2 981877: Полупроводниковое устройство и структура выводов Роберта Н. Нойса, Fairchild Semiconductor, поданный 30 июля 1959 г. и выданный 25 апреля 1961 г. Хотя Нойс подал заявку на это изобретение через два месяца после Килби, патент Нойса был предоставлен более чем через два годами ранее, что способствовало ожесточенной битве между Texas Instruments и Fairchild за то, кто именно изобрел интегральную схему.
Пожалуйста, НЕ копируйте наши статьи в блоги и другие сайты
статей с этого сайта зарегистрированы в Бюро регистрации авторских прав США. Копирование или иное использование зарегистрированных работ без разрешения, удаление этого или других уведомлений об авторских правах и / или нарушение смежных прав может привести к серьезным гражданским или уголовным санкциям.
Авторские права на текст © Chris Woodford 2009, 2020. Все права защищены. Полное уведомление об авторских правах и условиях использования.
Подписывайтесь на нас
Сохранить или поделиться этой страницей
Нажмите CTRL + D, чтобы добавить эту страницу в закладки на будущее, или расскажите об этом друзьям с помощью:
Цитируйте эту страницу
Вудфорд, Крис.(2009/2020) Интегральные схемы. Получено с https://www.explainthatstuff.com/integratedcircuits.html. [Доступ (укажите дату здесь)]
Больше на нашем сайте …
Раскраска таможенной статистики. Или сколько и каких микросхем ввозят в Россию?
Когда в 2008 году мы разрабатывали микроконтроллеры серии K1986VE9x, возникло много вопросов. Сколько должно быть Flash? сколько оперативной памяти? сколько ног? какие интерфейсы? и т.п.И хотя на тот момент просто создание работающей микросхемы уже имело бы успех, мы смогли сделать так, чтобы эта серия по-прежнему пользовалась спросом и ее продажи растут. Отчасти из-за того, что эти вопросы тогда были заданы, и ответы на них позволили принять взвешенное решение. В 2021 году перед нашей компанией по-прежнему стоят похожие вопросы, и даже шире. Например, что лучше делать в условиях ограниченных ресурсов: маломощный микроконтроллер или миниатюрный Flash или блок питания, а если блок питания, то какой? О том, как мы искали ответы на эти вопросы в 2008 году и как мы их ищем, читайте в этой статье.
Начало пути – 2008 г.
Как мы ответили на эти вопросы в 2008 году? Маркетинговые исследования, высокооплачиваемые консультанты, эксперты, аналитические отчеты, жизненный опыт? Нет, ничего из этого не работает. Точнее, в 2008 году не получилось, все было субъективно, аргументировано только на словах. Потому что не было данных. Никто не мог сказать, сколько и каких микросхем ввозится в страну или сколько вообще потребляется в мире. Многие предприятия даже не имели собственной внутренней базы данных по использованным товарам.Продавцы не раскрывают объем продаж. Ничего не указывало. Поэтому было решено, как всегда, опираться на опыт зарубежных аналогов. Наверное, все те же маркетологи STM, NXP и прочие Renesas лучше «чувствуют» рынок, чем мы. И хотя они тоже ни за что не скажут, сколько и каких микросхем продают, но по популярности (а значит и массе) микросхемы можно судить по разнообразию номенклатуры, представленной на рынке.Те. чем больше различных вариантов микроконтроллера с одним объемом Flash-памяти от нескольких производителей, тем больше будет емкость рынка, а значит, микроконтроллер с таким объемом будет более востребован. Если среди всех 32-битных микроконтроллеров больше всего номиналов размером ~ 100 футов, то именно 100 контактов – это число, которое наиболее востребовано разработчиками оборудования. Для анализа были проанализированы сайты ведущих разработчиков микроконтроллеров – TI, STM, NXP, Renesas, Microchip и ныне несуществующей EnergyMicro.Через формы поиска комплектующих выставляем необходимый параметр, например, объем Flash (16К, 32К, 64К и так далее) и подсчитываем, сколько номиналов нам дал, собрал и проанализировал производитель.
Что произошло с объемом Flash в 2008 году?
STMNXPEnergyMicroTI (+ их как раз тогда купил Luminary) MicrochipRenesasНу в общем для индустрии
Всего по всем производителямЕще раз, это не означает, что микроконтроллеры 256 КБ были самыми продаваемыми (у нас не было данных о продажах в 2008 году), но это означает, что микроконтроллеры 256 КБ предлагаются больше всего в различных номиналах.И, скорее всего, поскольку они самые предлагаемые, то размер рынка для них самый большой. Конечно, у этого метода много недостатков (один и тот же номинал может иметь разные градации в диапазонах температур, и это будет учтено как разные номиналы) Но то есть, то есть. Аналогичное исследование было проведено по количеству контактов в корпусе.
Распределение по количеству выводов упаковки для всех производителейПо количеству оперативной памяти и другим параметрам.
Объем оперативной памяти в микроконтроллерахИ хотя не все полученные результаты были реализованы, на их основе были сформулированы требования к характеристикам для серии. K1986VE9x… А еще я решил, что второй раз не буду собирать данные по рукопашному бою, а у нас есть компьютеры – пусть считают.
Сегодня. Большие данные
Потом было много чего… Вывод на рынок, исправление ошибок, новые проекты, митинги, планы, импортозамещение, встречи в министерствах, пандемия, коронавирус, видеоконференцсвязь и как вишенка на торте – кризис дефицита микросхем на мировом рынке в 2021 г.…
Все это время мы собирали данные.Многие предприятия уже охотнее делятся своей базой импортных ЕЦБ, кто-то фактически реализовал программу импортозамещения, кого-то заставили власти. Появилась статистика ФТС по ввозу электронных компонентов и многие другие данные, которые уже включают реальные объемы потребления микросхем в штуках и в деньгах. Политика Минпромторга РФ в части преференций для отечественной продукции подтолкнула многих гражданских разработчиков взглянуть на отечественные микросхемы, а возникший в начале 2021 года дефицит микросхем заставил серьезно задуматься о переходе на них:
То, о чем мы мечтали в 2008 году, в 2021 году превратилось в кошмар.К нам стали приходить выгрузки Excel из сотен и тысяч наименований зарубежных микросхем с просьбой подсказать аналоги, а для этого нам необходимо:
– найти в интернете документацию на чужую микросхему;
– прочитать и понять его назначение;
– для определения его основных параметров;
– выберите наше решение максимально близко;
– приведем как близкий аналог с указанием основных отличий.
И еще хорошо, будь то микроконтроллер или память. Что такое актуатор? В целом перспективы растут в геометрической прогрессии.Но мы продолжаем это делать, поскольку именно эти знания дают понимание того, что нужно делать и где развиваться. Но как уже было сказано ранее – эту замечательную работу может выполнить компьютер, а не человек. Поэтому мы разработали функционал для автоматического выбора назначения и основных параметров микросхемы по ее названию. И тут же возникла мысль: а что, если «разбирать» не список закупок какого-либо предприятия, а сразу все ввозимые через таможню иностранные микросхемы? И сегодня мы попробуем «раскрасить» таможенную статистику за 2019 год.Сразу скажу, что результат был не очень полным. В подавляющем большинстве импортных микросхем название не указывается. Например, один из крупнейших производителей RFID-меток импортировал около 80 миллионов микросхем NXP по цене около 3 рублей за штуку с названием «Интегральная микросхема». Также в код микросхемы попало множество дискретных компонентов (транзисторы, диоды…). Они дают большую прибавку в штуках, но практически не дают в цене. После всех чисток и еще небольшого ручного труда (самые крупные позиции по стоимости и по штукам, которые не были найдены автоматически и тем не менее были пробиты через Интернет), мы получили базу импортированных микросхем за 2019 год из 18 тысяч наименований с всего 85 микросхем.млн и на общую сумму 45 млн долларов. Всего мы смогли распознать 82% по штуке и 75% по стоимости. Мы получили интересную статистику, которая дала нам пищу для размышлений.
Общее распределение по типам микросхем:
Больше всего было ввезено микросхем, используемых в цепях питания (линейные регуляторы, DC-DC, AC-DC, источники опорного напряжения, предварительные драйверы, переключатели питания, драйверы светодиодов и др.) – около 46 миллионов штук. Еще есть непризнанные микросхемы – 15 миллионов.
На третьем месте простые логические схемы (регистры, счетчики, триггеры, вентиляторы и др.) – более 11 миллионов штук. На четвертом месте по количеству интерфейсных цепей (RS-485, RS-232, USB, Ethernet и так далее) – более 8 миллионов, далее идут простые аналоговые схемы (усилители, компараторы, мультиплексоры) – более 1,7 миллиона. куски. Я ожидал большего от микроконтроллеров и микропроцессоров, но всего их было 1,3 миллиона. Скорее всего, микроконтроллеры разносятся по «серым» каналам.А потом АЦП / ЦАП – 391 тысяча, тактовые генераторы – 352 тысячи, схемы памяти – всего 133 тысячи (снова «серые» каналы) и схемы программируемой логики FPGA / CPLD – 89 тысяч.
Как таможенная стоимость распределяется между одними и теми же классами, можно увидеть на следующем рисунке:
На первом месте по стоимости – Прочие (непризнанные схемы), на втором – схемы питания – более 10 миллионов долларов – но это из-за стоимости, на третьем месте – FPGA / CPLD – более 8 миллионов долларов. , несмотря на то, что их ввозили меньше всего в штуках.
Если посчитать удельную стоимость одной микросхемы, то получим такую картинку
Самыми маргинальными являются FPGA / CPLD, на втором месте схемы АЦП / ЦАП, на третьем месте микроконтроллеры / микропроцессоры и на четвертом месте схемы памяти. “Где вы видели эти цены?” – воскликнут многие читатели. Это не для меня, это для таможни. Но если вы решили заняться разработкой микросхем, то нужно заняться программируемой логикой.
Кто популярнее Xilinx или Altera?
ПобедилаAltera, хотя мне больше нравится Xilinx.Но по стоимости лидирует Xilinx.
Интересно, что и другие производители показали высокую стоимость, но секрет прост – это связано с китайскими радиационно-стойкими ПЛИС, которые, кстати, являются клонами Xilinx.
Если вам интересно, какая логическая емкость является наиболее популярной, будет большим сюрпризом, что более 50% всех программируемых логических массивов являются CPLD, то есть схемами малой емкости до 10K логических блоков (примерно до 100K логических элементов).
Чаще всего используются очень маленькие схемы (EPM240, XC95xx) до 500 логических вентилей – ~ 20 тысяч штук.Следующий пик популярности – линейка логических блоков 2К-4К (около 40К вентилей) – 12 тысяч. Поэтому мы продолжаем упорно «пилить» наш проект MLDR125.
Думаю, для начала достаточно. Напишите в комментариях, какую еще занимательную статистику выкладывать в следующей части статьи. Например, кто производит больше всего АЦП – TI vs Analog? Или какой самый популярный размер энергонезависимой памяти в SOIC8? Кто лидер в России по производству микроконтроллеров – ST, NXP, Renesas или TI? Какие драйверы светодиодов самые популярные?
Количественное исследование способности вспоминания модели вычислительной микросхемы гиппокампа | Brain Informatics
Полная вычислительная модель микросхемы CA1 Cutsuridis et al.[17] изображена на рис. 1. Полная модель состояла из 100 ПК, одной аксо-аксонной клетки (AAC), двух корзинчатых клеток (BCs), одной BSC и одной клетки oriens lacunosum-molculare (OLM). Энторинальный кортикальный (EC) возбуждающий вход возбуждал дистальные дендриты PC, AAC и BC, тогда как возбуждающий коллатеральный вход CA3 Шаффера возбуждал проксимальные дендриты PC, AAC, BC и BSC. Ингибирующий вход медиальной перегородки (МС) подавлял все тормозящие клетки в сети и заставлял их срабатывать в определенных фазах тета-ритма.
В этом исследовании, где проверялась только способность микросхемы вспоминать, когда в ее синапсах сохранялось все большее количество паттернов памяти, без изучения точных деталей процесса обучения (хранения), подсеть полной модели микросхемы была используется. Подсеть состояла из N, ПК ( N = 100 или 300), одного BSC и одной ячейки OLM (см. Рис. 2). Упрощенная морфология, включая сомы, апикальные и базальные дендриты и часть аксона, использовалась для каждого типа клеток.Биофизические свойства каждой клетки были адаптированы из типов клеток, описанных в литературе, которые были всесторонне подтверждены экспериментальными данными в [24,25,26,27]. BC и AAC, хотя и присутствовали в сети, были отключены от нее и были неактивны во время цикла поиска из-за сильного ингибирования MS и, следовательно, не влияли на динамику сети. Вход EC, хотя и присутствует, также не влияет на ячейки сети, потому что он также был отключен. Единственное возбуждение в сети было от CA3, который возбуждал дендриты BSC и PC.Все симуляции были выполнены с использованием NEURON [28], запущенного на ПК с 4 ЦП под Windows 8. Трассы напряжения всех подключенных (ПК, BSC, OLM) и отключенных (AAC, BC) ячеек к сети относительно одного тета-цикла изображены на рис. 3.
Рис. 2(Слева) Вспомните модель микросхемы области CA1 гиппокампа и (справа) модель CA1-PC с одним возбуждающим (CA3) и шестью тормозными (BSC) синаптическими контактами на его дендриты. ЭК: энторинальный корковый вход; CA3: Внесение залога Schaffer; AAC: аксо-аксоническая клетка; BC: корзина ячейка; BSC: бистратифицированная ячейка; OLM: oriens lacunosum-molculare cell; SLM: stratum lacunosum-molculare; SR: stratum radiatum; SP: пирамидальный слой; SO: stratum oriens.Во время отзыва активны только ПК, BSC и ячейка OLM. AAC и BC неактивны из-за сильного ингибирования медиальной перегородки. BSC и PC управляются на своих SR-дендритах сильным возбуждающим сигналом CA3, который представляет собой контекстную информацию. Вход EC отключен от сети, что не оказывает на него никакого воздействия. Красные кружки на дендритах ПК представляют загруженные синапсы, тогда как черные кружки на дендритах ПК представляют ненагруженные синапсы
Рис. 3Кривые напряжения модельных ячеек относительно одного тета-цикла
Чтобы помочь читателям этой работы и повысить читаемость В нашей рукописи мы приводим ниже краткие описания компонентов каждой сети.Заинтересованным читателям следует обратиться к исследованиям [17, 29] для получения более подробной информации о модели микросхемы и ее компонентов, включая размеры соматических, аксонических и дендритных компартментов их клеток, а также распределение пассивной и активной проводимости, синаптических волн и синаптических проводимостей вдоль эти отсеки. Полный математический формализм модели можно найти в дополнительных онлайн-материалах [29].
Пирамидальные ячейки
Каждый ПК имел 15 отсеков.Каждый отсек содержал кальциевый насос и механизмы буферизации, активируемые кальцием медленные постгиперполяризованные (AHP) и средние токи AHP K + , высоковольтный активированный (HVA) ток Ca 2+ L-типа, HVA R- тип Ca 2+ ток, низковольтный активированный (LVA) T-тип Ca 2+ ток, h-ток, быстрый натрий и ток выпрямителя с задержкой K + ток, медленно инактивирующийся M-тип K Ток + и быстро отключающий ток А-типа К + [24, 25].
Каждый ПК получил среднедендритное возбуждение от коллатералей Шаффера (CA3-PC), проксимальное возбуждение от примерно 1% других ПК CA1 в сети (рекуррентные коллатерали) [30], пространственно распределенное (шесть контактов) проксимальное дендритное синаптическое торможение от BSC и дистальное синаптическое ингибирование на каждой дистальной [stratum lacunosum-molculare (SLM)] дендритной ветви от клетки OLM.
Бистратифицированная ячейка
BSC имела 13 отсеков. Каждый отсек содержал ток утечки, натриевый ток, быстродействующий выпрямитель с задержкой K + , ток A-типа K + , токи L- и N-типа Ca 2+ , ток Ca 2+ -зависимый ток K + , а также Ca 2+ – и зависящий от напряжения ток K + [26].Он получал возбуждение от коллатералей CA3 Schaffer в его медиальных дендритных компартментах, возбуждение от активных ПК CA1 в его базальных дендритах и ингибирование от MS в его базальных дендритных компартментах.
Ячейка OLM
Ячейка OLM имела четыре отделения. Каждый отсек имел натриевый (Na + ) ток, ток выпрямителя с задержкой K + , ток K + типа A и ток h [27]. Он получал возбуждение от ПК в его базальных дендритах и ингибирование от МС в соме.
Входные данные модели
Возбуждающий вход, исходящий от аксонов коллатеральных пирамидных клеток CA3-Шаффера, и тормозной вход, исходящий от МС, управляли клетками сети во время отзыва (см. Рис. 4). Вход CA3 был смоделирован как срабатывание M ( M = 5, 10 или 20) из N ( N = 100 или 300) пирамидных ячеек CA3 при средней гамма-частоте 40 Гц (пик тренирует только смоделированные, а не явные ячейки). PC и BSC получали возбуждающий вход CA3 в своих медиальных дендритах.Ингибирование МС моделировалось как ритмическое срабатывание двух популяций из десяти перегородочных клеток (MS 1 и MS 2 ), каждая из которых модулировалась в противоположных фазах тета-цикла (сдвиг по фазе на 180 °) [31] (см. Рис. 4). ). Выход каждой септальной клетки моделировали как всплески потенциалов действия с использованием генератора пресинаптических спайков. Каждая последовательность спайков состояла из всплесков потенциалов действия со средней частотой 8 Гц в течение полутета-цикла (70 мс), за которым следовали пол-тета-цикла молчания. Из-за 8% шума в интервалах между спайками десять последовательностей спайков в каждой популяции перегородки были асинхронными.Во время отзыва ячейки MS 1 заблокировали ячейки MS 2 , что заблокировало ячейки BSC и OLM в сети.
Рис. 4Растровый график , показывающий пики перегородки (верхние 20) и входной сигнал CA3 (нижние 100). b Растровый график, показывающий активность 20 «активных ячеек», кодирующих конкретный образец памяти. c Двадцать спайков «активных ячеек» в скользящем интервале 10 мс. d Качество отзыва в скользящем интервале 10 мс
Синаптические свойства
Учитывались синапсы AMPA, NMDA, GABA-A и GABA-B.ГАМК-А присутствовала в соматическом и дендритном компартментах, тогда как ГАМК-В присутствовала только в медиальных и дистальных дендритах. Синапсы AMPA и NMDA присутствовали только в медиальных дендритах.
Тестирование сети
Целью данной исследовательской работы было проверить способность модели вспомнить, когда в сети уже были сохранены шаблоны, без изучения точных деталей процесса обучения. Для проверки способности модели к воспроизведению была принята методология, описанная в [17].Образец памяти был сохранен путем создания весовой матрицы на основе усеченного правила обучения Хебба. Эта матрица весов использовалась для предварительного определения весов подключения ПК CA3 – CA1. Без потери общности, входные (CA3) и выходные (CA1) шаблоны предполагались одинаковыми, причем каждый шаблон состоял из M ( M = 5, 10 или 20) случайно выбранных ПК (активных ячеек) на выходе. населения N ( N = 100 или 300) шт. N × N ( N × N = 100 × 100 или 300 × 300) -мерная матрица весов была создана путем установки записи матрицы ( i , j ), w ij = 1, если входной ПК i и выходной ПК j активны в одной и той же паре шаблонов; в противном случае веса равны 0.Для создания этой двоичной матрицы весов можно сохранить любое количество пар шаблонов. Матрица была применена к нашей сетевой модели путем подключения входа CA3 к ПК CA1 с высокой проводимостью AMPA (gAMPA = 1,5 нСм), если их вес подключения был равен 1, или с низкой проводимостью (gAMPA = 0,5 нСм), если их подключение было 0. Этот подход подтверждается экспериментальными данными в пользу синапсов с двумя состояниями [32].
Шаблоны памяти
Наборы шаблонов памяти разного размера (1, 5, 10, 20), перекрытия шаблонов (0%, 10%, 20%, 40%) и количества активных ячеек в шаблоне (5, 10 , 20) были созданы.0% перекрытие, например, между пятью шаблонами в наборе означает отсутствие перекрытия между шаблонами 1 и 2, 1 и 3, 1 и 4, 1 и 5, 2 и 3, 2 и 4, 2 и 5, 3 и 4, 3 и 5, и 4 и 5. Точно так же 40% перекрытие между пятью шаблонами в наборе означало, что 0,4 × Μ ячеек совместно использовались между шаблонами 1 и 2, различные ячейки 0,4 × Μ совместно использовались между шаблонами 2 и 3, разные ячейки 0,4 × Μ между шаблонами 3 и 4, разные ячейки 0,4 × Μ между шаблонами 4 и 5 и разные 0. {{N} _ {p}} {C} _ {i}} {{N} _ {p}}, $$
(2)
, где C i – качество отзыва шаблона i и N p – общее количество вызванных шаблонов.Например, когда десять шаблонов ( N p = 10) были первоначально сохранены в сети и шаблон 1 был представлен в сеть во время отзыва, то было вычислено значение качества отзыва для шаблона 1 (C1). Повторяя этот процесс для каждого из других шаблонов [шаблон 2 (C2), шаблон 3 (C3),…, шаблон 10 (C10)], было вычислено значение качества отзыва. Среднее качество запоминания (MC) сети было тогда средним значением этих индивидуальных качеств запоминания.
Выбор модели
В [17] ингибирование BSC дендритов ПК действовало как глобальная неспецифическая пороговая машина, способная устранять ложные действия на сетевом уровне во время отзыва.Ингибирование BSC сохранялось постоянным по мере того, как на дендритные синапсы ПК загружалось больше паттернов. Качество отзыва модели из [17] снижалось по мере того, как сохранялось все больше и больше воспоминаний (см. Рис. 14 в [17]).
Для улучшения способности вспоминания [17] мы искусственно модулировали синаптическую силу избирательных возбуждающих и тормозных путей к BSC и PC-дендритам по мере того, как в сети сохранялось все больше и больше паттернов:
- 1.
Модель 1: усиление возбуждающего синаптического привода CA3 прямого распространения к дендритам BSC (рис.6а) увеличена скорострельность ББК. В результате в дендритах ПК было образовано больше IPSP, создавая очень сильную ингибирующую среду, которая устраняла всю ложную активность.
- 2.
Модель 2: усиление прямого ингибирующего синаптического привода BSC к PC дендритам (рис. 6b) дает меньше IPSPs, но с большей амплитудой.
- 3.
Модель 3: усиление возбуждающего синаптического драйва PC-обратной связи к базальным дендритам BSC (рис. 6c) имело тот же эффект, что и модель 1, но с меньшей эффективностью.
Схематический чертеж a «модель 1», b «модель 2» и c «модель 3». В «модели 1» сильный возбуждающий вход CA3 увеличивает ответную реакцию BSC, которая генерирует на дендритах ПК многочисленные IPSP с малой амплитудой, создавая, таким образом, очень сильную ингибирующую среду, которая отфильтровывает ложную активность нейронов.В «модели 2» сильное подавляющее воздействие BSC на дендриты ПК вызывает постсинаптически меньшее количество IPSP, но с большей амплитудой. В «модели 3» сильный возбуждающий сигнал обратной связи ПК с BSC увеличивает его ответную реакцию, которая генерирует меньше IPSP, чем «модель 1» на дендрите ПК, и, следовательно, менее сильную ингибирующую среду, чем «модель 1»
Введение в нейроны и Нейронные сети | Раздел 1, Вступительная глава | Neuroscience Online: Электронный учебник для неврологии | Кафедра нейробиологии и анатомии
Три фунта желеобразного материала, обнаруженные в наших черепах, – это самая сложная машина на Земле и, возможно, во Вселенной.Его феноменальные свойства были бы невозможны без сотен миллиардов нейронов, из которых он состоит, и, что важно, без связей между этими нейронами. К счастью, многое известно о свойствах отдельных нейронов и простых нейронных сетей, и аспекты сложных нейронных сетей начинают разгадываться. Эта глава начнется с обсуждения нейрона, элементарного узла или элемента мозга, а затем перейдет к обсуждению способов, которыми отдельные нейроны общаются друг с другом.Что делает нервную систему таким фантастическим устройством и отличает мозг от других органов тела, так это не то, что она имеет 100 миллиардов нейронов, а то, что нервные клетки способны взаимодействовать друг с другом таким высокоструктурированным образом, что формируют нейронные сети. . Чтобы понять нейронные сети, необходимо понять способы, которыми один нейрон взаимодействует с другим через синаптические связи, и процесс, называемый синаптической передачей. Синаптическая передача бывает двух основных видов: возбуждение и торможение.Всего несколько взаимосвязанных нейронов (микросхема ) могут выполнять сложные задачи, такие как опосредовать рефлексы, обрабатывать сенсорную информацию, генерировать движения и опосредовать обучение и память. Более сложные сети ( макросхем, ) состоят из множества встроенных микросхем. Макросхемы опосредуют высшие функции мозга, такие как распознавание объектов и познание. Итак, в нервной системе повсеместно встречаются сети нескольких уровней. Сети также распространены в пределах нейронов.Эти наноцепей составляют лежащий в основе биохимический механизм для обеспечения ключевых свойств нейронов, таких как обучение и память, и генезиса ритмичности нейронов. Нейрон
Основные морфологические особенности нейронов
100 миллиардов нейронов в головном мозге имеют ряд общих черт (рис. 1). Нейроны отличаются от большинства других клеток тела тем, что они поляризованы и имеют различные морфологические области, каждая из которых выполняет определенные функции. Дендриты – это область, где один нейрон получает связи от других нейронов. Тело клетки или сома содержит ядро и другие органеллы, необходимые для клеточной функции. Аксон является ключевым компонентом нервных клеток, по которому информация передается от одной части нейрона (например, тела клетки) к концевым областям нейрона. Аксоны могут быть довольно длинными, достигая метра или около того в некоторых сенсорных и двигательных нервных клетках человека.Синапс является конечной областью аксона, и именно здесь один нейрон образует соединение с другим и передает информацию посредством процесса синаптической передачи . Нейрон цвета морской волны на рисунке 1 (щелкните «Нейрон, связанный с постсинаптическим нейроном») называется постсинаптическим нейроном. Терминал коричневого цвета слева, следовательно, называется пресинаптическим нейроном. Один нейрон может получать контакты от множества разных нейронов.На рисунке 1 (щелкните «Нейрон, получающий синаптический ввод») показан пример трех пресинаптических нейронов, контактирующих с одним постсинаптическим нейроном коричневого цвета, но было подсчитано, что один нейрон может получать контакты до 10 000 других клеток. Следовательно, потенциальная сложность сетей огромна. Точно так же любой нейрон может контактировать до 10 000 постсинаптических клеток. (Обратите внимание, что нейрон коричневого цвета, который был пресинаптическим по отношению к нейрону цвета морской волны, является постсинаптическим по отношению к розовым, зеленым и синим нейронам.Таким образом, большинство «пресинаптических» нейронов являются «постсинаптическими» по отношению к некоторым другим нейронам.
Рисунок 1 (щелкните «Синапс») также показывает расширенный вид синапса. Обратите внимание, что пресинаптическая клетка не связана напрямую с постсинаптической клеткой. Они разделены промежутком, известным как синаптическая щель . Следовательно, чтобы общаться с постсинаптической клеткой, пресинаптический нейрон должен выпускать химический мессенджер. Этот мессенджер находится внутри везикул, содержащих нейротрансмиттер, (синие точки обозначают нейромедиатор).Потенциал действия, который проникает в пресинаптический терминал, заставляет эти везикулы сливаться с внутренней поверхностью пресинаптической мембраны и высвобождать их содержимое посредством процесса, называемого экзоцитоз . Освободившийся передатчик диффундирует через промежуток между пре- и постсинаптической клеткой и очень быстро достигает постсинаптической стороны синапса, где связывается со специализированными рецепторами, которые «распознают» передатчик. Связывание с рецепторами приводит к изменению проницаемости ионных каналов в мембране и, в свою очередь, к изменению мембранного потенциала постсинаптического нейрона, известного как постсинаптический синаптический потенциал (PSP) .Таким образом, передача сигналов между нейронами связана с изменениями электрических свойств нейронов. Чтобы понять нейроны и нейронные цепи, необходимо понимать электрические свойства нервных клеток.
Потенциал покоя и потенциал действия
Потенциалы покоя. На рисунке 2 показан пример идеализированной нервной клетки. Во внеклеточную среду помещен микроэлектрод. Микроэлектрод – это не что иное, как небольшой кусок стеклянной капиллярной трубки, который растягивается под действием тепла, образуя очень тонкий наконечник, порядка 1 микрона в диаметре.Микроэлектрод заполняется проводящим раствором и затем подключается к подходящему записывающему устройству, например, осциллографу или самописцу. Когда электрод находится вне клетки во внеклеточной среде, регистрируется нулевой потенциал, поскольку внеклеточная среда является изопотенциальной. Однако, если электрод проникает в ячейку так, что кончик электрода теперь находится внутри ячейки, на записывающем устройстве наблюдается резкое отклонение. Регистрируется потенциал около -60 милливольт внутри отрицательного по отношению к внешней стороне.Этот потенциал называется потенциалом покоя и остается постоянным в течение неопределенных периодов времени при отсутствии какой-либо стимуляции. Если электрод удален, снова регистрируется нулевой потенциал. Потенциалы покоя – это не просто характеристики нервных клеток; все клетки тела обладают потенциалом покоя. Что отличает нервные клетки от других возбудимых мембран (например, мышечные клетки), так это то, что они способны изменять свой потенциал покоя. В случае нервных клеток для интеграции информации и передачи информации, а в случае мышечных клеток – для производства мышечных сокращений.
Потенциалы действия. На рис. 3 показан еще один эскиз идеализированного нейрона. Этот нейрон был пронзен одним электродом для измерения потенциала покоя и вторым электродом, называемым стимулирующим электродом. Стимулирующий электрод подключается к батарее через переключатель. Если батарея ориентирована так, что положительный полюс подсоединен к переключателю, замыкание переключателя сделает внутреннюю часть элемента несколько более положительной в зависимости от размера батареи.(Такое уменьшение поляризованного состояния мембраны называется деполяризацией ). На рисунке 3 представлена анимация, в которой переключатель многократно размыкается и замыкается, и каждый раз, когда он замыкается, в схему подключается батарея большего размера. Первоначально замыкание переключателя вызывает только небольшие деполяризации. Однако потенциалы становятся больше, и в конечном итоге деполяризация становится достаточно большой, чтобы вызвать потенциал действия , также известный как импульс или импульс .Потенциал действия связан с очень быстрой деполяризацией, позволяющей достичь пикового значения около +40 мВ всего за 0,5 миллисекунды (мс). За пиком следует столь же быстрая фаза реполяризации .
Напряжение, при котором деполяризация становится достаточной для запуска потенциала действия, называется порогом . Если для генерации надпороговой деполяризации используется батарея большего размера, единичный потенциал действия все равно генерируется, и амплитуда этого потенциала действия такая же, как и потенциал действия, запускаемый только пороговым стимулом.Простая запись на рисунке 3 иллюстрирует две очень важные особенности потенциалов действия. Во-первых, они вызываются методом по принципу «все или ничего» . Либо потенциал действия вызывается стимулами на уровне порога или выше, либо потенциал действия не вызывается. Во-вторых, потенциалы действия – это очень короткие события продолжительностью всего несколько миллисекунд. Запуск потенциала действия в некоторой степени аналогичен подаче спички на предохранитель. Определенная температура необходима для воспламенения предохранителя (т.е.е., предохранитель имеет порог). Спичка, выделяющая большее количество тепла, чем пороговая температура, не приведет к тому, что предохранитель будет гореть ярче или быстрее. Так же, как потенциалы действия вызываются по принципу «все или ничего», они также распространяются по принципу «все или ничего». Как только потенциал действия инициируется в одной области нейрона, такой как тело клетки, этот потенциал действия будет распространяться по аксону (как горящий предохранитель) и в конечном итоге проникнет в синапс, где он может инициировать процесс синаптической передачи.
В примере на рисунке 3 был сгенерирован только один потенциал действия, потому что продолжительность каждого из двух надпороговых стимулов была настолько короткой, что было достаточно времени только для инициирования одного потенциала действия (т. Е. Стимул закончился раньше, чем потенциал действия завершил свой цикл деполяризации-реполяризации). Но, как показано в анимации на Рисунке 4, более продолжительные стимулы могут привести к инициированию множественных потенциалов действия, частота которых зависит от интенсивности стимула.Таким образом, очевидно, что нервная система кодирует информацию не с точки зрения изменения амплитуды потенциалов действия, а, скорее, с точки зрения их частоты. Это очень универсальное свойство. Чем выше интенсивность механического стимула к сенсорному рецептору, тем больше количество потенциалов действия; чем сильнее растягивается рецептор растяжения мышцы, тем больше количество потенциалов действия; чем больше интенсивность света, тем большее количество потенциалов действия передается в центральную нервную систему.Точно так же в двигательной системе, чем больше количество потенциалов действия в двигательном нейроне, тем сильнее будет сокращение мышцы, которая получает синаптическую связь от этого двигательного нейрона. Инженеры называют этот тип кодирования информации частотно-импульсной модуляцией .
Синаптические потенциалы и синаптическая интеграция
На рисунке 5 показаны три нейрона. Зеленый цвет будет называться возбуждающим нейроном по причинам, которые вскоре станут ясны.Он соединяется с постсинаптическим нейроном синего цвета. Следы ниже (нажмите «Play») иллюстрируют последствия запуска потенциала действия в зеленом нейроне. Этот потенциал действия в пресинаптическом нейроне приводит к снижению мембранного потенциала постсинаптической клетки. Мембранный потенциал изменяется от значения покоя около -60 милливольт до более деполяризованного состояния. Этот потенциал называется возбуждающим постсинаптическим потенциалом (ВПСП) .Он является «возбуждающим», потому что перемещает мембранный потенциал к пороговому значению, и является «постсинаптическим», потому что это потенциал, зарегистрированный на постсинаптической стороне синапса. Обычно (и это важный момент) единичный потенциал действия в пресинаптической клетке не производит ВПСП, достаточно большого, чтобы достичь порога и запустить потенциал действия. Но, если в пресинаптической клетке запускаются несколько потенциалов действия, соответствующие множественные потенциалы возбуждения могут суммироваться посредством процесса, называемого временным суммированием , чтобы достичь порога и запустить потенциал действия.ВПСП можно рассматривать как «сигнал движения» постсинаптическому нейрону для передачи информации по сетевому пути.
Нейрон красного цвета на рисунке 5 называется тормозным нейроном. Как и зеленый нейрон, он также устанавливает синаптический контакт с синим постсинаптическим нейроном. Он также высвобождает химический передатчик-мессенджер, но последствия связывания передатчика от синей клетки с рецепторами на постсинаптической клетке противоположны последствиям передатчика, выпущенного зеленым нейроном.Следствием потенциала действия красного пресинаптического нейрона является увеличение мембранного потенциала синего постсинаптического нейрона. Мембранный потенциал более отрицательный, чем был раньше (гиперполяризация ), и поэтому мембранный потенциал дальше от порогового значения. Этот тип потенциала называется ингибирующим постсинаптическим потенциалом (IPSP) , потому что он предотвращает активацию постсинаптического нейрона потенциала действия. Это «стоп-сигнал» для постсинаптической клетки.Таким образом, зеленый нейрон говорит «вперед», а красный нейрон говорит «стоп». Что же делать постсинаптическому нейрону?
Нейроны похожи на счетные машины. Они постоянно суммируют возбуждающий и тормозящий синаптический вход во времени (временное суммирование) и по площади дендритов, получающих синаптические контакты ( пространственное суммирование) , и если это суммирование находится на пороговом уровне или выше, они запускают потенциал действия. Если сумма ниже порога, потенциал действия не инициируется.Этот процесс называется синаптическая интеграция и проиллюстрирован на рисунке 5. Первоначально два потенциала действия в зеленом нейроне производили суммирующие ВПСП, которые запускали потенциал действия в синем нейроне. Но если IPSP от тормозящего нейрона возникает непосредственно перед двумя потенциалами действия в возбуждающем нейроне, сумма одного IPSP и двух EPSP оказывается ниже порогового значения, и в постсинаптической клетке не возникает никакого потенциала действия. Тормозящий нейрон (и ингибирование в целом) – это способ стробирования или регулирования способности возбуждающего сигнала запускать постсинаптическую клетку.
Нейронные сети
Мотивы микросетей
Как указывалось ранее в этой главе, нейрон может получать контакты до 10 000 пресинаптических нейронов, и, в свою очередь, любой нейрон может контактировать до 10 000 постсинаптических нейронов. Комбинаторная возможность может привести к чрезвычайно сложным нейронным цепям или сетевым топологиям , что может быть очень трудно понять. Но, несмотря на потенциально огромную сложность, многое можно узнать о функционировании нейронных цепей, исследуя свойства подмножества простых конфигураций цепей.На рисунке 6 показаны некоторые из этих мотивов микросхем или микросетей. Несмотря на простоту, они могут делать многое из того, что должна делать нервная система.
Возбуждение с прямой связью . Позволяет одному нейрону передавать информацию своему соседу. Длинные цепочки из них могут использоваться для передачи информации через нервную систему.
Запрещение прямой связи .Пресинаптическая клетка возбуждает тормозящий интернейрон (интернейрон – это нейрон, расположенный между двумя нейронами), и этот тормозной интернейрон затем подавляет следующую клетку-последователь. Это способ выключения или ограничения возбуждения в нейроне ниже по потоку в нейронной цепи.
Конвергенция / Расхождение . Одна постсинаптическая клетка получает конвергентный вход от ряда различных пресинаптических клеток, и любой отдельный нейрон может создавать дивергентные связи со многими разными постсинаптическими клетками.Дивергенция позволяет одному нейрону общаться со многими другими нейронами в сети. Конвергенция позволяет нейрону получать входные данные от многих нейронов в сети.
Боковое торможение . Пресинаптическая клетка возбуждает тормозящие интернейроны, и они подавляют соседние клетки в сети. Как подробно описано ниже в этой главе, этот тип схемы может использоваться в сенсорных системах для улучшения контуров.
Запрет обратной связи / повторения .На панели E1 пресинаптическая клетка соединяется с постсинаптической клеткой, а постсинаптическая клетка, в свою очередь, соединяется с интернейроном, который затем подавляет пресинаптическую клетку. Этот контур может ограничивать возбуждение в пути. Некоторое начальное возбуждение будет отключено после того, как красный интернейрон станет активным. На панели E2 каждый нейрон в замкнутой цепи подавляет нейрон, с которым он связан. Казалось бы, эта схема ничего не делает, но, как будет показано далее в этой главе, она может привести к генерации сложных паттернов всплесковой активности.
Обратная связь / рекуррентное возбуждение . На панели F1 пресинаптический нейрон возбуждает постсинаптический нейрон, а этот постсинаптический нейрон возбуждает пресинаптический нейрон. Этот тип цепи может выполнять функцию переключателя, потому что, как только пресинаптическая клетка активируется, эта активация может сохраняться. Активация пресинаптического нейрона могла включить эту сеть, и она могла остаться. На панели F2 показаны варианты возбуждения обратной связи, при которых пресинаптический нейрон возбуждает постсинаптический нейрон, который может по обратной связи возбуждать сам себя (a, autapse ) или другие нейроны, которые в конечном итоге имеют обратную связь (b) с самим собой.
Эти простые мотивы – вездесущие компоненты многих нейронных цепей. Давайте рассмотрим несколько примеров того, на что способны эти сети.
Возбуждение с прямой связью и подавление с прямой связью
Одна из наиболее изученных микросхем – это схема, которая обеспечивает простое рефлекторное поведение. На рисунке 7 показана схема так называемого рефлекса рефлекса коленного рефлекса или растяжения. Невролог ударяет резиновым молоточком по колену, в результате чего нога разгибается.Этот тест используется как простой способ проверить целостность некоторых сенсорных и моторных путей в спинном мозге. Удар молотка растягивает мышцу и приводит к возникновению потенциалов действия в сенсорных нейронах внутри мышцы, которые чувствительны к растяжению. (Потенциалы действия представлены маленькими яркими «огоньками» на анимации.) Потенциалы действия инициируются по принципу «все или ничего» и распространяются в спинной мозг, где аксон разделяется (раздваивается) на две ветви.
Давайте сначала обсудим ветвь слева, которая формирует синаптическую связь (зеленый треугольник) с двигательным нейроном Extensor (E) (окрашен в синий цвет). Потенциал действия сенсорного нейрона вторгается в синаптический терминал сенсорного нейрона, вызывая высвобождение медиатора и последующее возбуждение двигательного нейрона. Растяжение мышцы приводит к возникновению потенциала действия в двигательном нейроне (МН), который затем распространяется по периферическому нерву и вторгается в синапс в мышце, вызывая высвобождение передатчика и потенциал действия в мышце.Потенциал действия в мышечной клетке приводит к сокращению мышцы и разгибанию конечности. Итак, здесь у нас есть простая схема возбуждения с прямой связью, которая опосредует поведение.
Теперь давайте рассмотрим правую ветвь аксона сенсорного нейрона на рисунке 7. Потенциал действия сенсорного нейрона вторгается в синаптический терминал сенсорного нейрона, вызывая высвобождение медиатора и последующее возбуждение постсинаптического интернейрона, окрашенного в черный цвет.Этот нейрон называется интернейроном , потому что он расположен между одним нейроном (здесь SN) и другим нейроном (здесь MN). Возбуждение интернейрона приводит к инициированию действия и последующему высвобождению передатчика из пресинаптического терминала интернейрона (черный треугольник), но для этой ветви цепи передатчик ведет к IPSP в постсинаптическом сгибателе (F ) мотонейрон (окрашен в красный цвет). Функциональные последствия этого подавления прямой связи заключаются в уменьшении вероятности того, что двигательный нейрон-сгибатель станет активным и вызовет несоответствующее сгибание ноги.
Сходимость и расхождение
Упрощенная схема, опосредующая рефлекс растяжения, представлена на рисунке 8. Однако правильное функционирование схемы рефлекса растяжения также зависит от конвергенции и расхождения. Один сенсор имеет несколько ветвей, которые расходятся и образуют синаптические связи со многими отдельными моторными нейронами (щелкните «Дивергенция»). Следовательно, когда мышца сокращается в результате нажатия невролога, это происходит потому, что несколько мышечных волокон активируются одновременно несколькими двигательными нейронами.Кроме того, когда мышца растягивается, активируется не один, а несколько сенсорных нейронов, и все сенсорные нейроны проецируются в спинной мозг, где они сходятся к отдельным моторным нейронам-разгибателям (нажмите «Конвергенция»). Итак, рефлекс растяжения возникает из-за комбинированных эффектов активации нескольких сенсорных нейронов и моторных нейронов-разгибателей.
Боковое торможение
Улучшение края.Боковое торможение очень важно для обработки сенсорной информации. Одним из примеров является явление в зрительной системе, называемое усилением краев. На рисунке 9 показаны две полосы: темно-серая полоса слева и светло-серая полоса справа. Хотя темная полоса и светлая полоса имеют одинаковую яркость по всему полю, при внимательном рассмотрении обнаруживается, что светло-серая полоса кажется несколько светлее на границе темно-серой полосы, чем в других областях поля.Напротив, темно-серая полоса на границе выглядит несколько темнее, чем в других областях темного поля. Это явление улучшения границ, которое помогает визуальной системе извлекать важную информацию из визуальных сцен. Усиление краев опосредуется, по крайней мере частично, боковым торможением в сетчатке.
Давайте сначала рассмотрим схему без бокового торможения (Рисунок 10, щелкните «Без бокового торможения»). Свет падает на сетчатку (Часть A), а его интенсивность можно описать ступенчатым градиентом (Часть B).Для упрощения предположим, что темно-серая область имеет интенсивность пяти единиц, а светло-серая область – десять единиц. Градиент света активирует фоторецепторы, и фоторецепторы создают синаптические связи с нейронами второго порядка. Предположим, что интенсивность света 5 единиц приводит к 5 всплескам / с, а интенсивность света 10 единиц приводит к 10 всплескам / с (Часть C) в фоторецепторах, и что синаптическая сила достаточна (здесь обозначена как +1) так что интенсивность света 5 единиц приводит к 5 импульсам / с, а интенсивность света 10 единиц приводит к 10 импульсам / с (Часть C) соответственно в нейронах второго порядка.Если никакой дальнейшей обработки информации не происходит, воспринимаемый градиент будет точно таким же, как градиент интенсивности света (часть B, красный след). Но это не то, что воспринимается, и латеральное торможение объясняет разницу.
Теперь рассмотрим расширенную схему с боковым торможением (нажмите «С боковым торможением»). Каждый из фоторецепторов устанавливает тормозные синаптические связи со своим соседним нейроном второго порядка. Сила подавления (обозначается -0.2) меньше силы возбуждения (пожертвовано +1). Прежде чем смотреть на границу, рассмотрите выход схемы в однородных областях каждого поля. Далеко справа от границы все клетки получают одинаковое возбуждение и одинаковое торможение. Без бокового торможения интенсивность света в 10 единиц произвела бы 10 импульсов в секунду в нейроне второго порядка. Но из-за тормозящей связи от соседних нейронов справа и слева выходная мощность снижается до 6 импульсов в секунду.То же самое верно для клеток, расположенных далеко слева от границы, но величина возбуждения меньше и, соответственно, величина торможения меньше. Ключевая обработка происходит на границе или краю. Обратите внимание, что нейрон справа от границы получает такое же подавление от нейрона справа от нее, но получает меньшее подавление от нейрона слева от него на другой стороне границы. Следовательно, он получает больше чистого возбуждения и имеет выходной сигнал 7 пиков / с, а не 6 пиков / с его соседа справа.Теперь посмотрите на нейрон слева от границы. Он получает слабое торможение от своего соседа слева, но более сильное торможение от своего соседа справа по другую сторону границы. Следовательно, он получает меньше чистого возбуждения и имеет выходную мощность 2 пика / с, а не 3 пика / с его соседа слева. Таким образом, в результате бокового торможения информация, передаваемая нервной системе, и воспринимаемый градиент будут версией исходной с усиленной границей или краем (рис. 10B)!
ДиапазонМаха.Простая схема сетчатки с боковым торможением может объяснить феномен усиления краев. Этим также можно объяснить визуальную иллюзию, известную как полосы Маха. На рисунке 11 показан градиент светлых и темных вертикальных полос, поперек которых проходит тонкая горизонтальная линия. Похоже, что горизонтальная линия имеет неравномерное распределение интенсивности: она темнее в области светлых вертикальных градиентов и светлее в области темных вертикальных градиентов. Это визуальная иллюзия .Иллюзию можно раскрыть, поместив маску поверх вертикального градиента. (Нажмите «Play», чтобы добавить маску.) Теперь вы можете видеть, что горизонтальная полоса имеет равномерную интенсивность. В некоторых областях он воспринимается более темным, потому что клетки сетчатки, которые реагируют на более темную область горизонтальной полосы, сильно подавляются клетками, реагирующими на яркую область вертикальной полосы. Напротив, полоса воспринимается ярче в некоторых областях, потому что клетки сетчатки, которые реагируют на более светлую область горизонтальной полосы, только слабо ингибируются клетками, реагирующими на темную область вертикальной полосы.
Обратная связь / повторное подавление
Запрет обратной связи в микросхемах. Подавление обратной связи играет общую роль в гашении возбуждения через нейронную цепь. Классическим примером является клетка Реншоу в спинном мозге. Аксон ветви спинномозгового мотонейрона. Одна ветвь иннервирует мышцу, как описано ранее (например, рис. 7), а другая ветвь создает возбуждающую синаптическую связь с интернейроном, называемым клеткой Реншоу.Интернейрон, в свою очередь, подавляет двигательный нейрон, замыкая петлю. Другой пример подавления обратной связи обнаружен в гиппокампе. Пирамидные клетки СА3-типа создают возбуждающие связи с клетками корзины и обратную связь с клетками корзины, чтобы ингибировать клетки СА3. Термин повторяющееся ингибирование применяется к простым схемам подавления обратной связи, таким как цепь Реншоу в спинном мозге и цепь корзиночных клеток в гиппокампе.
Подавление обратной связи в наноцепях.Подавление обратной связи распространено не только во многих нейронных цепях; он также широко распространен в биохимических цепях. Здесь он может служить подложкой для генерации колебаний. Они могут охватывать несколько временных масштабов от секунд до дней в зависимости от молекулярных компонентов схемы.
Рисунок 12. Из Byrne, Canavier, Lechner, Clark and Baxter, 1996. |
- Эндогенное взрывное поведение в нейронах.Идеализированный нейрон, описанный ранее в этой главе, молчал в отсутствие стимуляции (например, рисунок 3). Однако некоторые нейроны активируют потенциалы действия в отсутствие стимуляции, и в некоторых случаях паттерны возбуждения могут демонстрировать взрывной паттерн, в котором последовательные высокочастотные всплески активности сменяются периодами покоя. Такие нейронные свойства могут быть важны для создания ритмического поведения, такого как дыхание. На рисунке 12 показан пример записи нейрона беспозвоночного, имеющего эндогенный импульсный ритм.Этот конкретный нейрон называется параболическим всплеском, потому что характер интервалов между всплесками является длинным в начале и в конце цикла всплеска, но очень коротким в середине цикла. Клетка запускает всплеск потенциалов действия, а затем замолкает, но вскоре происходит еще один всплеск, и этот процесс продолжается бесконечно каждые десять-пятнадцать секунд. Взрыв происходит, даже если нейрон хирургическим путем удален из ганглия и помещен в культуру, чтобы не было синаптических связей с другими нейронами.Таким образом, для этого ритма не требуется нейронная сеть – она эндогенная. Но это действительно связано с наноконтуром внутри клетки. На рисунке 13A показана очень упрощенная версия этой сети, в которой подчеркивается ключевой принцип работы. Критическим для этой сетевой функции является канал в
мембрана (обозначена g SI ), проницаемая для Ca 2+ . Поскольку концентрация Ca 2+ относительно высока во внеклеточной среде и низка внутри клетки, Ca 2+ будет двигаться вниз по градиенту концентрации и тем самым деполяризует клетку.В конце концов, деполяризация достигает порога, и клетка начинает срабатывать. Обжиг приводит к дополнительному притоку Ca 2+ (зеленая стрелка) и накоплению Ca 2+ внутри клетки. Ключевым этапом является то, что накопление Ca 2+ ингибирует (красная стрелка) дальнейший приток кальция и прекращает всплеск. Всплеск остается прекращенным, пока уровни внутриклеточного Ca 2+ остаются повышенными. Но уровни Ca 2+ не остаются повышенными надолго.Они восстанавливаются внутриклеточными буферами и удаляются из клетки с помощью насосов (I NaCa и I CaP ). Когда внутриклеточные уровни Ca 2+ снижаются, ингибирование канала снимается (растормаживание ), и нейрон снова начинает деполяризоваться, и начинается новый всплеск. По сути, то, что мы видим здесь на уровне наносети, является повторением сети, препятствующей обратной связи (рис. 13B). Первоначальный процесс возбуждения приводит к активации тормозного процесса, который отключается от него и прекращает его.В такой сети возникнут колебания, если возбуждающее побуждение будет непрерывным, но тормозящий процесс ослабевает по своей эффективности.
Циркадные ритмы.Второй пример подавления обратной связи – это наноцепь регуляции генов, лежащих в основе циркадных ритмов. Циркадный ритм позвоночных обусловлен работой группы нейронов в области мозга, называемой супрахиазматическим ядром, которая расположена чуть выше зрительного нерва. Эти нейроны оказывают сильное влияние как на высвобождение гормонов, таких как мелатонин, кортизол, так и на вегетативные функции, такие как температура тела (рис. 14). Несмотря на огромное влияние этого генератора, его работа сводится к очень простой схеме, и действительно, не к нейронной схеме, а к другой наноконтуре.Похоже, что основной механизм сохраняется у всех видов животных, включая человека. На рисунке 15 представлена упрощенная принципиальная схема основных компонентов. Участвует несколько генов, но основной механизм включает ген под названием на , где на – на период. Этот ген был впервые идентифицирован у плодовой мухи Drosophila , но также присутствует у позвоночных. на генов приводит к производству на матричных РНК. на мРНК покидает ядро и попадает в цитоплазму, где приводит к синтезу белка PER.PER диффундирует или транспортируется обратно в ядро, где репрессирует дальнейшую транскрипцию гена на . Концептуально эта система очень похожа на механизм разрывного нейрона, рассмотренный выше. Ген активируется, он производит сообщение и белок, а белковая обратная связь подавляет экспрессию гена. Но как цикл повторяется? Ключевым механизмом является деградация PER. Белок PER разлагается и разлагается в течение 24 часов.Так как белок PER разрушается, подавление или репрессия снимается (растормаживание), позволяя этому гену снова начать создавать информационную РНК и белок. Итак, как только этот цикл начинается, он повторяется снова и снова через 24 часа. Это основной механизм, лежащий в основе циркадных ритмов и мощного воздействия, которое они оказывают на ряд различных физиологических систем. По сути, все наши циркадные ритмы начинаются с сети подавления молекулярной обратной связи.Рисунок 14. По материалам Hastings et al., Nature Rev. Neurosci. , 2003.
Запрет обратной связи в кольцевых цепях.Повторяющееся торможение может, по крайней мере в принципе, объяснить генерацию сложных двигательных паттернов, примером которых является передвижение на четвероногих ногах. Четвероногие животные интересны тем, что четвероногие способны не только двигать своими четырьмя ногами, но и генерировать различные типы циклов активности, называемые походками. На рисунке 16 показаны четыре аллюра. Первая панель – это прогулка (тап по собаке). Последовательность начинается с разгибания левой передней конечности. Затем следуют разгибания правой задней конечности, правой передней конечности и левой задней конечности.На рыси (вторая панель рисунка 16) (постукивание по собаке) левая передняя и правая задние конечности находятся в фазе друг с другом и на 180 градусов не совпадают по фазе с правой передней и левой задней конечностями. В связке (третья панель) (постукивание по собаке) левая передняя и правая передние конечности находятся в фазе, но на 180 градусов не совпадают по фазе с левой задней и задней задними конечностями. Галоп (четвертая панель) (постукивание по собаке) – вариант скачек, в котором есть небольшая разница фаз между правой и левой передними и задними конечностями.
Как нервная система генерирует эти походки? И нужны ли для каждой отдельные нейронные цепи? К сожалению, нейробиологи не знают ответов на эти вопросы, но поучительно изучить некоторые возможности. Это подход в области нейробиологии под названием Вычислительная и теоретическая нейробиология . Один из способов создания походки проиллюстрирован на рисунке 17. Возьмите четыре отдельных нейрона, каждый с эндогенной импульсной активностью, подобной тому, который показан ранее на рисунке 12, и назначьте активность в каждом из этих нейронов контролю конкретной конечности.Нейроны могут быть «запущены» так, чтобы у них были соответствующие фазовые соотношения для генерации походки, такие как граница, показанная на рисунке 16. Трудность будет заключаться в запуске нейронов точно в точное время. Другой проблемой могут быть небольшие «дрейфы» в колебательных периодах четырех независимых нейронов, которые со временем приведут к тому, что паттерн станет несогласованным (рис. 18). Эта собака не выиграет ни одной гонки и, вероятно, не сможет ходить.
Итак, очевидно, что нейроны должны быть связаны.Один из способов сделать это – использовать повторяющуюся цепь ингибирования, состоящую из четырех связанных нейронов, чтобы сформировать так называемую «кольцевую» цепь, где каждый нейрон в цепи имеет эндогенную импульсную активность, и каждый нейрон связан с другим с помощью тормозной синаптической связи ( Рисунок 19A).
Рисунок 19. По материалам Canavier, Butera, Dror, Baxter, Clark and Byrne, 1997. |
Чтобы получить правильные фазовые соотношения для походок, вместо того, чтобы назначать нейрон 3 правую переднюю конечность, он назначается для управления правой задней конечностью, а нейрон 4 назначается для управления правой передней конечностью ( простой поворот схемы) (Рисунок 19Б).При реализации с помощью компьютерного моделирования эта единственная схема способна генерировать четвероногие ворота. Более того, одна и та же схема с небольшими изменениями свойств отдельных нейронов может генерировать каждую из четырех походок, показанных на рисунке 17 (рисунок 20).
Этот результат указывает на важный момент, касающийся нейронных сетей.Чтобы понять их, необходимо понимать не только топологию сети, но и природу связей между нейронами (являются ли они возбуждающими или тормозящими), а также свойства отдельных узлов (т. Е. нейроны). Кроме того, это моделирование иллюстрирует явление, называемое динамической реконфигурацией . Для генерации этих четырех разных походок не обязательно иметь четыре разные сети – все это можно сделать с помощью одной схемы.Фактическая схема, генерирующая квадрапедальную походку, более сложна, чем схема на рис. 19. Заинтересованный читатель может обратиться к недавнему обзору Оле Кейна (см. Дополнительную литературу).
Обратная связь / рекуррентное возбуждение
Периодическое возбуждение в наноцепях и микросхемах имеет решающее значение для процессов обучения и памяти. Обучение включает в себя изменения биофизических свойств нейронов и изменения синаптической силы. Накапливающиеся данные указывают на то, что положительная обратная связь внутри биохимических каскадов и генных сетей является важным компонентом для индукции и поддержания этих изменений.Более того, периодическое возбуждение обнаруживается по крайней мере в некоторых микросхемах, участвующих в процессах памяти. Яркий пример находится в области CA3 гиппокампа.
Рисунок 21. Изменено из Byrne and Roberts, 2009. |
На рисунке 21 показаны основные характеристики рекуррентной возбуждающей цепи CA3. Шесть различных пирамидных нейронов гиппокампа обозначены как U , V , W , X , Y и Z .Каждый из этих нейронов получает синаптическую связь от пресинаптических нейронов, обозначенных a , b , c , d , e и f. Эти пресинаптические нейроны могут быть активными или неактивными с 0 и черным цветом, представляющим неактивный нейрон, и 1 и зеленым цветом, указывающим активный. Важным аспектом этой схемы является то, что синаптические связи входного пути достаточно сильны, чтобы активировать (активировать) пирамидный нейрон, с которым они связаны.Например, если активирован нейрон a , будет активирован нейрон Z , который представлен как 1 на панели вывода. Эта топология – не что иное, как возбуждение с прямой связью. Взаимное возбуждение делает эту схему особенной. Например, нейрон Z и другие пирамидные нейроны имеют коллатерали аксонов, которые посредством обратной связи соединяются друг с другом. Но они не только устанавливают связь между собой. Каждый нейрон устанавливает связь с каждым из четырех других пирамидных нейронов в цепи.Таким образом, каждая пирамида получает сходящуюся информацию от всех других клеток в сети, и, в свою очередь, выход каждого пирамидального нейрона расходится, чтобы установить синаптические связи со всеми другими пирамидными нейронами в цепи. (Следовательно, этот повторяющийся мотив возбуждения вложил в себя мотивы конвергенции и расхождения.) Матрица связности состоит из 36 элементов.
Чтобы эта сеть могла чему-либо научиться, в схему необходимо встроить правило обучения синаптической пластичности.Широко признано правило Хебба . По сути, в нем говорится, что синапс изменит свою силу, если этот синапс активен (то есть высвобождает передатчик), и в то же время активна постсинаптическая клетка. Комбинация этого правила обучения и повторяющегося возбуждающего контура приводит к некоторым интересным эмерджентным свойствам. Например, если нейрон Z активируется входом a, , сила его соединения с самим собой (синапс 1 ) изменится, как показано зеленым синапсом на рисунке 21 (анимация находится в 2-секундном цикле, чтобы иллюстрируют контрольные и модифицированные синапсы).Однако синапс 1 не будет единственным синапсом, который будет усилен. Например, синапс 13 также будет усилен, потому что нейрон Z был активен в то же время, когда нейрон X был активирован входом c . Напротив, синапс 7 не усиливается, потому что нейрон Y не был активен в то же время, что и нейрон Z . Чистый эффект этой конвергенции и расхождения и правила обучения состоит в том, что исходный входной шаблон активности будет сохранен как изменения в элементах матрицы связности.Поэтому этот канал был назван сетью автоматического связывания . Важная концепция здесь заключается в том, что «память» находится не в каком-то одном синапсе; это распределенных в сети .
Сводка
Значительный прогресс был достигнут в понимании того, как разные простые нейронные сети участвуют в обработке информации и посредническом поведении. Возбуждение с прямой связью и торможение с прямой связью опосредуют рефлекторное поведение.Боковое торможение важно для усиления краев. Повторяющееся возбуждение – важный механизм памяти. Повторяющееся торможение может иметь важное значение для формирования локомоторного поведения. В этих микросхемах заложены сходимость и расхождение. Одни и те же типы сетевых мотивов воспроизводятся в биохимических и генных сетях.
Следующий уровень понимания находится на уровне нейронных сетей, которые опосредуют более сложные, так называемые функции мозга высшего порядка.Их понимание становится возможным благодаря использованию электрофизиологических и оптических методов записи и современных методов визуализации, таких как функциональная магнитно-резонансная томография (фМРТ) и диффузионно-тензорная визуализация (DTI) . ФМРТ позволяет исследователям идентифицировать области мозга, которые заняты когнитивными задачами, тогда как DTI позволяет визуализировать пути, связывающие одну область мозга с другой. Рисунок 22 (любезно предоставлено Тимом Элмором, доктором философии, отдел.из нейрохирургии, Медицинская школа Техасского университета в Хьюстоне) – вид сбоку человеческого мозга, показывающий пути, соединяющие корковые области, выявленные с помощью DTI !.
Рисунок 22. Предоставлено Тимом Элмором, доктором философии. |
Распознавание объектов – это пример прогресса в понимании макросхем.Как показано на рисунке 23, обработка визуальной информации начинается в сетчатке, а затем затрагивает несколько областей коры, таких как затылочная кора и височная кора. Внутри этой макросхемы находятся модули, извлекающие информацию более высокого порядка. Предположительно, каждый модуль включает в себя сотни, если не тысячи отдельных микросхем. Задача на будущее – определить, как эти модули работают и как они взаимодействуют с другими модулями. Хотя соединения с прямой связью присутствуют, соединения с обратной связью и боковые соединения широко распространены.Задача огромна, но, возможно, достижению цели будет способствовать использование преимуществ того, что было изучено о принципах работы наносхем и микросхем. Чтобы понять макросхемы, необходимо знать больше, чем топологию сетевых соединений. Необходимо будет знать, как функционирует каждый модуль, и о динамике межмодульных соединений.
Рисунок 23.Из Феллемана и Ван Эссена, 1991. |
Проверьте свои знания
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
C. Запрет обратной связи
D. Боковое ингибирование
E.Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью. Этот ответ НЕПРАВИЛЬНЫЙ.
B. Ингибирование с прямой связью
C. Запрет обратной связи
D. Боковое ингибирование
E. Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
А.Возбуждение с прямой связью
B. Запрещение прямой связи. Этот ответ НЕПРАВИЛЬНЫЙ.
C. Запрет обратной связи
D. Боковое ингибирование
E. Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
с.Подавление обратной связи. Этот ответ НЕПРАВИЛЬНЫЙ.
D. Боковое ингибирование
E. Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
C. Запрет обратной связи
D. Боковое торможение. Ответ ПРАВИЛЬНЫЙ!
E.Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
C. Запрет обратной связи
D. Боковое ингибирование
E. Расхождение. Этот ответ НЕПРАВИЛЬНЫЙ.
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
А.Пластичность
B. Интеграция
C. Конвергенция
D. Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
A. Пластичность. Этот ответ НЕПРАВИЛЬНЫЙ.
B. Интеграция
C. Конвергенция
Д.Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
A. Пластичность
B. Интеграция. Этот ответ ПРАВИЛЬНЫЙ!
C. Конвергенция
D. Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
А.Пластичность
B. Интеграция
C. Конвергенция Этот ответ НЕПРАВИЛЬНЫЙ.
D. Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
A. Пластичность
B. Интеграция
C. Конвергенция
Д.Частотно-импульсная модуляция. Этот ответ НЕПРАВИЛЬНЫЙ.
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
A. Пластичность
B. Интеграция
C. Конвергенция
D. Частотно-импульсная модуляция
E. Растормаживание Этот ответ НЕПРАВИЛЬНЫЙ.
Сетевой мотив, лежащий в основе циркадных ритмов:
А.Временное суммирование
Б. Пространственное суммирование
C. Запрет обратной связи
D. Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование. Этот ответ НЕПРАВИЛЬНЫЙ.
Б. Пространственное суммирование
C. Запрет обратной связи
Д.Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование
B. Пространственное суммирование. Этот ответ НЕПРАВИЛЬНЫЙ.
C. Запрет обратной связи
D. Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
А.Временное суммирование
Б. Пространственное суммирование
C. Запрет обратной связи. Ответ ПРАВИЛЬНЫЙ!
D. Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование
Б. Пространственное суммирование
C. Запрет обратной связи
Д.Возбуждение обратной связи. Этот ответ НЕПРАВИЛЬНЫЙ.
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование
Б. Пространственное суммирование
C. Запрет обратной связи
D. Возбуждение обратной связи
E. Боковое торможение. Ответ НЕПРАВИЛЬНЫЙ.
Сеть с автоматической ассоциацией важна для:
А.Рефлексы
B. Улучшение кромки
C. Память
D. Передвижение
E. Нейронные колебания
Сеть с автоматической ассоциацией важна для:
A. Рефлексы. Этот ответ НЕПРАВИЛЬНЫЙ.
B. Улучшение кромки
C. Память
D. Передвижение
E. Нейронные колебания
Сеть с автоматической ассоциацией важна для:
А.Рефлексы
B. Улучшение края Этот ответ НЕПРАВИЛЬНЫЙ.
C. Память
D. Передвижение
E. Нейронные колебания
Сеть с автоматической ассоциацией важна для:
А. Рефлексы
B. Улучшение кромки
C. Память Этот ответ ПРАВИЛЬНЫЙ!
D. Передвижение
E.Нейронные колебания
Сеть с автоматической ассоциацией важна для:
А. Рефлексы
B. Улучшение кромки
C. Память
D. Передвижение. Этот ответ НЕПРАВИЛЬНЫЙ.
E. Нейронные колебания
Сеть с автоматической ассоциацией важна для:
А. Рефлексы
Б.Улучшение края
C. Память
D. Передвижение
E. Нейрональные колебания. Этот ответ НЕПРАВИЛЬНЫЙ.
пожертвований Neuroscience Online поможет финансировать разработку новых функций и контента. |
Знают ли ноги, что делает язык?
Abstract
Понимание адаптивного поведения требует точно контролируемого предъявления мультисенсорных стимулов в сочетании с одновременным измерением нескольких поведенческих модальностей.Таким образом, мы разработали устройство виртуальной реальности, которое позволяет одновременно измерять проверку вознаграждения, обычно используемую меру в парадигмах ассоциативного обучения, и навигационное поведение, а также точно контролируемое представление визуальных, слуховых и поощрительных стимулов. Крысы выполняли задачу виртуальной пространственной навигации, аналогичную лабиринту Морриса, где только дистальные зрительные или слуховые сигналы обеспечивали пространственную информацию. Карты пространственной навигации и проверки вознаграждения показали обучение, зависящее от опыта, и учитывали дистальные визуальные подсказки.Однако они показали диссоциацию, в результате чего дистальные слуховые сигналы не поддерживали пространственную навигацию, но поддерживали пространственно локализованную проверку вознаграждения. Эти данные показывают, что крысы могут перемещаться в виртуальном пространстве с помощью только дистальных визуальных сигналов, без значительных вестибулярных или других сенсорных входов. Более того, они обнаруживают одновременную диссоциацию двух моделей поведения, ориентированных на вознаграждение.
Образец цитирования: Cushman JD, Aharoni DB, Willers B, Ravassard P, Kees A, Vuong C, et al.(2013) Мультисенсорный контроль мультимодального поведения: знают ли ноги, что делает язык? PLoS ONE 8 (11): e80465. https://doi.org/10.1371/journal.pone.0080465
Редактор: Анна Дунаевская, Медицинский центр Университета Небраски, Соединенные Штаты Америки
Поступила: 21 августа 2013 г .; Одобрена: 2 октября 2013 г .; Опубликован: 4 ноября 2013 г.
Авторские права: © 2013 Cushman et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Эта работа была поддержана грантами для MRM от Национального научного фонда (NSF) Career # 0969034 (www.nsf.gov), NIH / CRCNS # 1-R01-MH-092925-01 (www.nih .gov), фонд Уайтхолла (www.whitehall.org) и фонд В.М. Кека (www.wmkeck.org). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Адаптивное поведение определяется широким спектром мультисенсорных входных стимулов, например слуховой и зрительный, и выражается в виде разнообразных поведенческих модальностей [1]. Чтобы полностью понять, как несколько форм поведения объединяются для создания адаптивного поведения, необходимо точно контролировать мультисенсорные стимулы и измерять их влияние одновременно на несколько поведенческих выходов.В частности, пространственное обучение изучается только с использованием моторной отдачи конечностей, например, в водном лабиринте Морриса [2], хотя другие поведенческие модальности также могут содержать пространственную информацию. Из-за технических ограничений одновременное измерение нескольких пространственно-модулированных поведений было невозможно, поскольку исследования обычно фокусировались на единственном сенсорном входе и единственной поведенческой модальности в данном устройстве. Поведение подхода, управляемое стимулами, предсказывающими вознаграждение, такими как журнал о еде или проверка трубки вознаграждения, широко изучалось в парадигмах ассоциативного обучения [3] – [6], но неясно, показывает ли проверка вознаграждения пространственную модуляцию, как это было раньше. измеряется только в месте доставки награды в аппарате для кондиционирования.Чтобы преодолеть это ограничение, мы разработали мультисенсорный аппарат виртуальной реальности (VR), который позволяет одновременно измерять как навигационное поведение, так и проверку вознаграждения, а также точно контролируемое представление визуальных, слуховых и поощрительных стимулов.
Виртуальная реальность у грызунов недавно стала захватывающим и мощным инструментом, поскольку она облегчает электрофизиологические и оптические измерения, которые выигрывают от ограничения движения головы животного и обеспечивают точный контроль над сенсорными стимулами [7] – [13].Однако до сих пор приложения виртуальной реальности грызунов ограничивались визуальной модальностью, а используемые поведенческие задачи выполнялись либо на одномерных линейных треках [8] – [10], [12], [14], либо на двумерных плоскостях бесконечного размера. [7]. Более того, ни один из этих предыдущих подходов не предусматривал одновременной проверки вознаграждения. Используя наш аппарат виртуальной реальности, мы разработали задачу виртуальной пространственной навигации, смоделированную по образцу водного лабиринта Морриса, которая требует гибкой навигации от нескольких начальных точек до скрытой зоны вознаграждения на основе дистальных визуальных и / или слуховых сигналов [15].Наши результаты показывают, что крысы легко научились пространственной навигации в виртуальной реальности, несмотря на отсутствие значительных вестибулярных сигналов. Кроме того, проверка вознаграждения одновременно выражалась во время навигации, и наблюдалось значительное зависимое от опыта обучение как в навигационных картах, так и в картах проверки вознаграждения, однако эти два пространственно модулированных поведения демонстрировали диссоциацию, в результате чего дистальные слуховые сигналы не поддерживали пространственную навигацию, но поддерживали пространственную модулированная проверка вознаграждения.
Результаты
Мы разработали устройство виртуальной реальности с несколькими основными усовершенствованиями, которые позволили точно представить визуальные и слуховые стимулы и одновременно измерить вознаграждение во время навигации (подробности см. В Методах S1 и на Рисунке S1).Аппарат был неинвазивным и не требовал фиксации головы, что позволяло проводить длительные испытания в условиях низкого стресса и выражения естественного поведения, такого как вставание на дыбы, уход за шерстью и отдых. Этот аппарат обеспечивал три различных сенсорных режима: визуальный, слуховой и поощрительный. Конструкция экрана обеспечивала высокий уровень погружения в виртуальную среду, поскольку визуальные стимулы могли проецироваться повсюду, над крысой и непосредственно рядом с ней. Поддержка сферической беговой дорожки была очень тихой (44 дБ), что еще больше снизило нагрузку и позволило нам представить двухмерные позиционные слуховые сигналы, используя систему объемного звучания с семью динамиками и специальным звуковым программным обеспечением, которое использует Ambisonics более высокого порядка (Blue Ripple’s Rapture 3D см. рисунок S3).В отличие от традиционной акустической системы объемного звучания, наша система создает звуковой ландшафт, подобный визуальному ландшафту, создаваемому удаленными источниками. В лабиринте виртуальной реальности также доставлялись точно контролируемые поощрительные стимулы. Емкостной сенсорный датчик был прикреплен к трубке с сахаром и водой с наградой для измерения поведения при упреждающей проверке вознаграждения. Таким образом, этот метод позволял непрерывно измерять проверку вознаграждения с высоким временным разрешением (60 Гц) в любом месте мультисенсорного виртуального пространства и одновременно измерять навигационные характеристики и обучение животного (надежность измерения 1 оборота сферы. имело стандартное отклонение 0.01 оборот). Для изучения пространственной навигации мы сначала разработали процедуру, чтобы научить крыс взаимодействовать с виртуальными средами и ограничивать их навигацию конечным двумерным пространством и избегать виртуальных границ (см. Рисунок S2 и видео S1). Затем мы убедились, что крысы могут использовать нашу систему объемного звучания для управления как навигацией, так и проверкой вознаграждения до ближайших слуховых и визуальных маяков (см. Рисунок S3).
Быстрое обучение пространственной навигации в виртуальном пространстве
Мы обучили крыс задаче пространственной навигации (см. Видео S2, рис. 1A), смоделированной после стандартной задачи пространственной памяти, обычно называемой водным лабиринтом Морриса [15].Крысы стартовали в одном из четырех стартовых мест в каждом испытании. Их задача заключалась в навигации на основе дистальных сигналов к виртуальной «скрытой зоне вознаграждения», заранее определенному, немаркированному месту в лабиринте по отношению к дистальным аудиовизуальным сигналам (рис. 1B-E). После успешной навигации они были вознаграждены сахарной водой через трубку для лизания и телепортировались в другое случайное начальное место, чтобы начать следующее испытание после 2-секундного интервала между испытаниями. Их первоначальный шаблон поиска был случайным в сеансе 1, но стал довольно точным к сеансу 6 (рис. 1B против 1C).И задержка, чтобы найти скрытую зону вознаграждения, и расстояние, которое нужно пройти, чтобы добраться до нее, снизились до асимптотического уровня производительности в течение трех сеансов (рис. 1D, E). Чтобы проверить точность их когнитивной карты, размер зоны вознаграждения был уменьшен до 20 см в радиусе, и тренировка продолжалась до тех пор, пока не был выбран критерий: два последовательных дня из 40 испытаний в течение 30 минут. Наблюдалось явное увеличение времени, проведенного вокруг зоны вознаграждения во время выполнения этой более сложной задачи (рис. 1F).Чтобы количественно оценить это, мы рассчитали время занятости для каждого квадранта (см. Методы). Крысы проводили значительно больше времени в целевом квадранте по сравнению с другими квадрантами (рис. 1H). Наконец, было проведено испытание с зондом, в котором зона вознаграждения была инактивирована, и крысам позволяли исследовать в течение приблизительно 270 секунд. Крысы выборочно искали на участке зоны вознаграждения, проводя значительно больше времени в целевом квадранте (рис. 1G, H).
Рисунок 1. Навигационные характеристики в задаче виртуальной аудиовизуальной пространственной навигации.
(A) Схема виртуальной среды с указанием дистальных слуховых и визуальных сигналов, скрытой зоны вознаграждения и четырех начальных точек. (B) Пример путей от одной крысы из сеанса 1 st . Цвет каждого пути указывает начальную точку, цвет которой обозначен стрелками в A. (C) Пример путей от одной и той же крысы из сеанса 6 -го . (D) Кривая задержки получения вознаграждения по сеансам. F (5,35) = 4,266, p = 0,0061, сеанс 1 против сеанса 3: p <0.05, N = 6. (E) Кривая накопления расстояния, пройденного до вознаграждения за сеансы. F (5,35) = 3,00, p = 0,0296, сеанс 1 против сеанса 5: p <0,05. (F) 2-мерная гистограмма средней занятости, усредненной по последним четырем сессиям задач асимптотической производительности с меньшей зоной вознаграждения. (G) Пример пробного пути зонда. (H) Процентный квадрант измеряет время занятости во время выполнения последних четырех заданий и во время пробного исследования. Эффект квадранта: F (3,23) = 10,15, p = 0,007, F (3,23) = 10.9, p = 0,0005 соответственно.
https://doi.org/10.1371/journal.pone.0080465.g001
Одновременное измерение пространственно-модулированной проверки вознаграждения во время навигации
Чтобы понять, как приобретается поведение проверки вознаграждения, мы вычислили среднее расстояние от каждой проверки до зоны вознаграждения, нормированное на расстояние, ожидаемое случайным образом распределенными кликами (называемое нормализованным расстоянием проверки, см. Методы). Этот метод не учитывал вклад улучшенных характеристик навигации, так что пространственное уточнение поведения проверки можно было анализировать изолированно.Этот анализ показал, что проверка смещалась значительно ближе к зоне вознаграждения между сеансами приобретения, и это сохранялось во время асимптотической производительности с меньшей зоной вознаграждения (рис. 2А). Таким образом, по мере того, как крысы усваивали навигационный компонент задачи, они также смещали распределение своего проверочного поведения в сторону зоны вознаграждения. Важно отметить, что это было сверх того, чего можно было ожидать только благодаря улучшению их пространственной навигации. Кроме того, общая частота проверок снизилась (до 27% от начального значения) по мере того, как они освоили задачу (рис. 2B).Эти данные свидетельствуют о том, что частота проверок модулируется неопределенностью местоположения вознаграждения, так что по мере увеличения точности проверки ее общая частота снижается.
Рисунок 2. Анализ поведения проверки вознаграждения во время задачи виртуальной аудиовизуальной пространственной навигации.
(A) Кривая сбора данных нормализованного контрольного расстояния между сеансами и во время окончательного выполнения задачи. Это рассчитывается как: Среднее фактическое расстояние / Среднее случайное расстояние x 100. Обратите внимание, что 6 -й день сбора данных был потерян из-за неисправности емкостного датчика.«Финал» обозначает среднюю производительность за последние четыре сеанса задач. Эффект сеанса: F (5,35) = 3,384, p = 0,018, p <0,05 для 1 st по сравнению с 5 сеансом th , N = 6. (B) Нормализованная частота проверки по сеансам. Эффект сеанса: F (5,35) = 4,452, p = 0,0049, p <0,05 для 1 st по сравнению с 4 th сеанс (C) 2-мерная гистограмма нормализованной частоты проверок, усредненной по крысам во время заключительного задания представление. (D) Проверьте модуляцию скорости как функцию радиального расстояния от зоны вознаграждения в радиальных ячейках 4 см для окончательного выполнения задачи.Влияние интервала, F (14,89 = 9,241, p <0,0001, p <0,05 для ближайшего интервала относительно интервалов с 3-го по 15-й. (E) 2-мерная карта p-значений производительности одной крысы во время выполнения окончательной задачи. Красным обозначены области, где поведение проверки было значительно выше вероятности (p <0,01), а синим цветом обозначены области, где поведение проверки было значительно ниже вероятности. (F) Пример фактического поведения проверки (черная линия) как функция радиального расстояния от зоны вознаграждения относительно случайного поведения проверки (зеленая линия с заштрихованной SE).Красные точки представляют собой точки, значительно превышающие вероятность, а синие точки - точки, значительно меньшие вероятности.
https://doi.org/10.1371/journal.pone.0080465.g002
Анализ окончательной асимптотической производительности с меньшей зоной вознаграждения показал, что частота проверки вознаграждения была немного увеличена в целевом квадранте, но это не достигло статистической значимости. (Эффект квадранта: F (3,23) = 1,234, p = 0,3319, процент проверок в целевом квадранте: 28% + / – 1.74 SE), что, по-видимому, противоречит их пространственным навигационным критериям, показывающим предпочтение квадранта вознаграждения. Для дальнейшего анализа мы вычислили распределение частоты проверок вознаграждения в зависимости от позиции (рис. 2C), которое показало, что частота проверок была повышена в непосредственной близости от зоны вознаграждения. Поскольку частота проверок вознаграждений была очень низкой (0,36 +/– 0,19 Гц), было трудно надежно оценить уровень значимости проверки вознаграждений в 2D-ячейках. Следовательно, использовалась одномерная мера, в которой использовались радиальные интервалы, сосредоточенные вокруг зоны вознаграждения (определяется как: (Фактическая частота проверок в корзине – рандомизированная частота проверок в этой корзине) / рандомизированная частота проверок, см. Методы).Частота проверок была увеличена вокруг зоны вознаграждения, при этом ближайший 4-сантиметровый интервал показывает значительное увеличение частоты проверок (рис. 2D). Это показало, что проверка вознаграждения повышается в очень мелком пространственном масштабе только вокруг зоны вознаграждения, но не во всем квадранте NE. Типичный пример производительности одной крысы показан на рисунках 2E и F. Проверка была модулирована значительно выше вероятности в непосредственной близости от зоны вознаграждения (рисунок 2E). Точно так же проверка была значительно выше вероятности в первых шести радиальных ячейках, окружающих зону вознаграждения (рис. 2F).
Крысы полагаются на дистальные зрительные, а не на дистальные слуховые сигналы для пространственной навигации
Чтобы понять лежащий в основе мультисенсорный вклад в пространственную навигацию, мы систематически удаляли визуальные или слуховые сигналы (рис. 3А). Чтобы гарантировать сходство опыта и мотивации в разных условиях, использовался блокированный дизайн (см. Методы). Примеры путей (рис. 3В) показали, что крысы переходили в зону вознаграждения для аудиовизуальных и только визуальных испытаний, но показали в основном случайный шаблон поиска в испытаниях только со звуком.Испытания только аудио имели значительно увеличенную задержку и расстояние до вознаграждения, однако эти измерения не различались между аудиовизуальными и только визуальными испытаниями (рис. 3C). Точно так же процент времени, проведенного в целевом квадранте, был на уровне случайности в испытаниях только со звуком, в отличие от аудиовизуальных и визуальных испытаний, которые были значительно выше вероятности (Рисунок 3D). Эти результаты предполагают, что крысы полагались почти исключительно на дистальные визуальные сигналы, а не на слуховые, чтобы перейти к месту награждения.Этот тест был как можно более кратким, чтобы исследовать ранее приобретенную пространственную стратегию крыс, а не усвоить новые стратегии, которые возникнут при дальнейшем обучении. К сожалению, это привело к недостаточным данным для проведения статистически надежного анализа частоты проверок, и поэтому мы провели следующие эксперименты, чтобы решить эту проблему.
Рис. 3. Мультисенсорный вклад в виртуальную пространственную навигацию и проверку вознаграждения.
(A) Схема аудиовизуального (AV), только визуального (V) и слухового (A) виртуальных пространственных лабиринтов.Обозначения как на рисунке 1. (B) Пример путей для трех типов испытаний от одной крысы. Цвет каждого пути указывает начальное местоположение, обозначенное цветом стрелок в A. (C) Средняя задержка и расстояние до вознаграждения для каждого типа испытания. Эффект типа исследования, F (2,17) = 7,555, p = 0,01, F (2,17) = 8,911, p = 0,006, соответственно. A по сравнению с AV и V: p <0,05 для обоих показателей, N = 6. (D) Процент занятости в целевом квадранте для трех типов испытаний. Эффект типа исследования: F (2,17) = 13,19, p = 0,0064.A против AV: p = 0,013, A и V: p = 0,018.
https://doi.org/10.1371/journal.pone.0080465.g003
Диссоциация между пространственной навигацией и проверкой вознаграждения
Могут быть две возможные причины неспособности крыс ориентироваться, используя только дистальные слуховые сигналы в аудиовизуальной задаче. Во-первых, усвоению слуховых сигналов можно помешать или затмить [16] присутствие зрительных сигналов. Во-вторых, использование четырех дистальных слуховых сигналов могло привести к насыщению или загромождению слуховой обработки, что затруднило различение отдельных звуков.Чтобы исключить обе эти возможности, мы обучили крыс двум новым задачам пространственного обучения. Одна среда содержала только два дистальных слуховых сигнала, а другая – только две дистальных визуальных сигнала (рис. 4A) в той же пространственной конфигурации относительно скрытой зоны вознаграждения (см. Методы). Затем эти задания были обучены отдельно, сначала последовательно со звуковым заданием. Мы сосредоточили наш анализ на последних четырех сессиях асимптотической производительности в этих двух задачах. Задержка до вознаграждения была стабильной на протяжении этих сеансов и была значительно больше в слуховой области по сравнению с визуальной задачей (Эффект сеанса, F (3,24) = 0.938, р = 0,4392; Эффект задачи: F (1,24) = 8,989, p = 0,0171, взаимодействие: F (3,24) = 1,809, p = 0,1725, задержка для слуховой задачи: 36,7 ± 6,7 с, задержка для визуальной задачи: 18,96 ± 1,12 s, см. видео S3 и S4). Крысы продемонстрировали стратегию кружения в слуховой задаче, бегая на фиксированном расстоянии от визуально определяемого края стола, тогда как в визуальной задаче они продемонстрировали четкое свидетельство прямой навигации к квадранту вознаграждения из всех четырех начальных точек (рис. 4B, C). Крысы проводили значительно больше времени в целевом квадранте при выполнении зрительной, но не слуховой задачи (рис. 4E).Эти данные свидетельствуют о том, что крысы были способны формировать пространственную карту на основе двух дистальных зрительных сигналов, но не двух дистальных слуховых сигналов. Однако удивительно, что частота проверки вознаграждения была значительно увеличена в целевом квадранте не только для визуальной задачи (средняя частота проверки: 0,20 ± 0,06 Гц), но и для слуховой задачи (средняя частота проверки: 0,22 ± 0,08 Гц, рис. 4D, E). . Радиальный анализ количества проверок вознаграждения и информационного содержания показал очень схожий профиль в обеих задачах с большим подъемом в непосредственной близости от зоны вознаграждения (Рисунок 4F, Влияние типа испытания на информационное содержание: p = 0.78, визуальный: 0,149 бит, слуховой: 0,152 бит). Таким образом, при наличии дистальных слуховых сигналов крысы действительно демонстрируют повышенную частоту проверки вознаграждения вблизи зоны вознаграждения, несмотря на отсутствие доказательств пространственного обучения, основанного на традиционных навигационных критериях.
Рис. 4. Навигационные характеристики и проверка вознаграждения во время выполнения слуховых и визуальных задач виртуальной пространственной навигации.
(A) Схема слуховой задачи и визуальной задачи. Символы как на рисунке 3. (B) Примеры путей для каждой задачи.(C) 2-мерная гистограмма занятости, усредненная по крысам за 4 сеанса для каждого типа испытаний. (D) 2-мерная гистограмма нормализованной частоты проверок, усредненной по крысам за эти сеансы. Белые области указывают на недостаточное покрытие. (E) Процент занятости и частота проверок в целевом квадранте для двух задач. Двусторонний дисперсионный анализ для целевого эффекта по сравнению с другими квадрантами и типов слуховых и зрительных испытаний: эффект квадранта F (1,8) = 95,16, p <0,001, эффект типа задачи: F (1,8) = 2,08, p < 0,001, Взаимодействие квадранта и типа задачи: F (1,8) = 2.08, p = 0,002; Эффект квадранта для слуховой задачи: p = 0,097, процентное наполнение целевого квадранта: 23,9 ± 0,5% по сравнению со средней нецелевой занятостью: 25,36 ± 0,17%, обратите внимание, что занятость в целевом квадранте немного снижается, поскольку животные, попадающие в зону вознаграждения, телепортировался; для визуальной задачи: p <0,001, процентная занятость целевого квадранта: 59,6 ± 1,15% по сравнению со средней нецелевой занятостью: 13,5 ± 0,38%, N = 5. (F) Нормализованная частота проверок как функция расстояния от зоны вознаграждения в радиальных бункерах для обоих пробных типов.Эффект расстояния от награды: F (14,112) = 67,11, p <0,0001, p> 0,05 для эффектов типа задачи и взаимодействия.
https://doi.org/10.1371/journal.pone.0080465.g004
Обсуждение
Мы разработали новое мультисенсорное устройство виртуальной реальности, которое позволило нам представить точно контролируемые аудиовизуальные стимулы и стимулы вознаграждения и одновременно измерять поведение проверки вознаграждения вместе с виртуальной пространственной навигацией. В отличие от большинства существующих систем, наша система была неинвазивной, что сводило к минимуму стресс и позволяло проводить долгосрочные измерения в условиях низкого напряжения.Крысы легко освоили задачу виртуальной пространственной навигации, смоделированную по образцу водного лабиринта Морриса. Это была аппетитная версия обычно используемой аверсивной задачи в водном лабиринте, где от них требовалось перейти к немаркированной зоне вознаграждения, определяемой исключительно дистальными визуальными и / или слуховыми сигналами, чтобы получить жидкое вознаграждение. Важно отметить, что виртуальный лабиринт впервые позволил нам убедиться, что не было других сигналов, определяющих пространственное расположение награды, чего трудно достичь в реальном мире.Эта задача двумерной навигации не позволяла крысам использовать стратегии навигации по ориентирам, которые используются в типичных экспериментах мышей с фиксированной головой на одномерном виртуальном линейном треке [14]. Пространственная навигация в виртуальной реальности была продемонстрирована на людях [17], [18], однако это первая демонстрация на грызунах. Наши результаты показывают, что надежные карты пространственной навигации могут быть сформированы у крыс в отсутствие значимых вестибулярных сигналов, которые, как предполагалось, играют важную роль в пространственном обучении во многих теориях [19], [20].Это свидетельствует о гибкости механизмов, лежащих в основе пространственного обучения, что имеет важное значение для природы этих механизмов [12], [13], [21], а также практическое значение для использования виртуальной реальности в электрофизиологических исследованиях гиппокампа [ 7] – [13].
Временной ход пространственного обучения был довольно быстрым, сравнимым с таковым в реальном водном лабиринте, хотя автоматизированный и аппетитный характер устройства виртуальной реальности позволял проводить гораздо больше испытаний за один сеанс.Наши результаты показали, что пространственная навигация была основана на дистальных визуальных сигналах, а не на дистальных слуховых сигналах, когда оба метода обучались одновременно. Это было подтверждено, когда крыс тренировали в отдельных лабиринтах, где пространственная информация в дистальном направлении предоставлялась исключительно визуальными или слуховыми сигналами. Мы обнаружили, что они были неспособны сформировать пространственное представление на основе двух дистальных слуховых сигналов и вместо этого приняли стратегию стереотипного кружения, аналогичную паттерну, наблюдаемому у крыс с поражениями гиппокампа [22].Лишь несколько исследований изучали навигацию только по дистальным слуховым сигналам у грызунов и получили смешанные результаты [23], [24]. Примечательно, что мы продемонстрировали, что крысы могут точно перемещаться по звуковому маяку, хотя эта производительность была слабее, чем навигация по визуальному маяку (рисунок S3). Напротив, они успешно ориентировались только с двумя дистальными визуальными ориентирами. Это согласуется с визуальным преобладанием, наблюдаемым в реальных задачах у грызунов [23], [24], а также людей [25] – [28]. Это можно объяснить более слабой способностью крыс определять ориентацию слуха по сравнению со зрительным стимулом [29].Таким образом, хотя крысы ведут ночной образ жизни, в этих контролируемых условиях они в подавляющем большинстве полагались на визуальные, а не слуховые сигналы для навигации.
Во время изучения аудиовизуального лабиринта проверка вознаграждения постепенно смещалась в сторону зоны вознаграждения, больше, чем можно было бы предсказать с помощью одной только улучшенной навигации. Это первое свидетельство того, что проверка вознаграждения также значительно пространственно модулируется, и ее пространственная точность улучшается с опытом. Во время асимптотического выполнения в аудиовизуальном лабиринте проверка вознаграждения показала очень тонкую пространственную модуляцию, поскольку она повышалась только в непосредственной близости от зоны вознаграждения.Удивительно, но проверка вознаграждения также была увеличена вокруг зоны вознаграждения для слухового лабиринта. Фактически, проверка вознаграждения была одинаково пространственно точной как в слуховом, так и в чисто визуальном лабиринте (рис. 4E, F). Таким образом, несмотря на отсутствие доказательств пространственного обучения, основанного на навигационных критериях, проверка вознаграждения в значительной степени пространственно модулировалась дистальными слуховыми сигналами.
Эта замечательная диссоциация утверждает, что наши слуховые сигналы были достаточно заметными и точными, чтобы поддерживать пространственно модулированное поведение, что исключает более тривиальные объяснения их неспособности поддерживать пространственную навигацию.Однако, что более важно, это доказывает, что природа мультисенсорной обработки информации, которая лежит в основе каждого выходного поведения, показывает расхождение в некоторой точке потока обработки. Это согласуется с гипотезой о том, что пространственная навигация и проверка вознаграждения, обычно рассматриваемые как результат ассоциативного обучения [3] – [5], управляются системами параллельной памяти, которые работают в соответствии со своими собственными базовыми правилами или стилями обработки [1] . Эти две системы либо частично параллельны, т.е.е. каждая система имеет доступ к идентично обработанной информации и использует ее по-разному или более полностью параллельно, т.е. каждая система может представлять мультисенсорную информацию принципиально разными способами. Однако также возможно, что эта обработка не всегда параллельна, поскольку навигация и проверка вознаграждения регистрируются на основе дистальных визуальных сигналов, хотя для этого потребуется, чтобы проверка поведения переключала лежащие в ее основе механизмы обработки на основе модальностей стимулов, которые она имеет. имеется в наличии.
В ассоциативном обучении считается, что мультисенсорная информация интегрирована в конфигурационные представления, посредством которых соединения нескольких сенсорных элементов связаны вместе в единое целое, или гештальт [30] – [32]. Ни один сенсорный элемент точно не определял местоположение вознаграждения в задачах аудио или визуальной навигации, и поэтому наблюдаемое нами пространственно модулированное проверяющее поведение должно быть обусловлено ассоциациями с конфигурацией мультисенсорных стимулов в непосредственной близости от зоны вознаграждения.Важно отметить, что эти ассоциации могут формироваться независимо от модальности стимула, поскольку менее пространственно информативных дистальных слуховых сигналов достаточно для поддержания кондиционирования. Пространственная навигация, напротив, предполагает формирование пространственной когнитивной карты, которая представляет окружающую среду в аллоцентрической метрической системе координат [19], [33] – [35]. Таким образом, интригующая возможность состоит в том, что проверка вознаграждения управляется конфигурационной когнитивной картой, тогда как навигация управляется пространственной когнитивной картой.С другой стороны, может быть только одна когнитивная карта, управляющая как навигацией, так и проверкой вознаграждения, но порождающая различное поведение из-за более высоких метаболических затрат на первую, чем на последнюю. Например, ошибки в вычислении навигационного пространства связаны с большими энергетическими затратами, поскольку они приведут к навигации к неправильным местоположениям, в то время как ошибочная проверка вознаграждения не несет таких затрат. Следовательно, порог для навигационных решений должен быть высоким, что может привести к сильной зависимости от более пространственно информативной визуальной информации.
Основные нейронные схемы пространственной навигации и ассоциативного обучения в течение некоторого времени интенсивно исследуются, при этом энторинально-гиппокампальная система считается основным посредником как пространственной навигации, так и конфигурационных ассоциаций [1], [2], [20] , [30], [36] – [41]. Таким образом, наши результаты поднимают интригующую возможность того, что одна и та же структура одновременно участвует в генерации выходных данных для двух поведенческих систем. Будущие исследования, использующие мультисенсорную виртуальную реальность в сочетании с электрофизиологическими методами записи [12] во множестве соответствующих областей мозга, смогут более глубоко проникнуть в основные нейронные механизмы этой параллельной обработки информации.Кроме того, способность обучать крыс сложным пространственным задачам в виртуальной реальности дает возможность напрямую объединить исследования на грызунах и людях таким образом, который ранее был невозможен [17], [18], [42] – [44].
Материалы и методы
Животные
Для этих экспериментов использовали девять крыс-самцов Long Evans возрастом примерно 3 месяца в начале поведенческого обучения. Их поддерживали в нормальном 12-часовом цикле свет / темнота с поведенческим обучением и тестированием во время светлой фазы.Во время поведенческого обучения животных ограничивали в пище и воде (16–18 г корма, 25–35 мл воды в день). Все процедуры были выполнены в соответствии с руководящими принципами NIH и одобрены Комитетом по исследованиям на животных Калифорнийского университета в Лос-Анджелесе.
Общие процедуры подготовки к обучению
По прибытии крыс обрабатывали 15-30 минут в день. Это продолжалось не менее 5 дней, после чего мы начали более конкретные процедуры предварительной подготовки в течение 10–15 дней для возможного обучения виртуальной реальности.Это включало в себя три основных процедуры: 1) привыкание к привязи примерно на 30 минут в день, 2) привыкание к привязи к привязи на вершине сферической беговой дорожки и 3) предварительная тренировка ассоциации «тон награды – сахар воды». Последнее было сделано в камере кондиционирования рядом с аппаратом VR, где крысу обучали связывать тон поощрения с доставкой сахарной воды в течение четырех-пяти дней. Наградным сигналом был звуковой сигнал частотой 200 мс и частотой 1 кГц. За этим последовало открытие клапана подачи сахарной воды в течение 400 мс, повторенное 5 раз.
Задача виртуальной пространственной навигации
Мы разработали задачу пространственного обучения по образцу водного лабиринта Морриса [15]. Виртуальный мир (рис. 1A) состоял из круглого стола радиусом 1 метр, расположенного на высоте 125 см над полом в центре комнаты размером 4,5х4,5 м с отчетливыми визуальными сигналами на каждой стене, а также четырьмя отдельными сложными слуховыми сигналами (северный звук: Частотная развертка от 1 до 5 кГц, повторяется один раз в секунду; восточный звук: сложный звук с пиком на частоте 2,3 кГц, повторяется три раза в секунду; южный звук: 10 кГц щелчок повторяется 10 раз в секунду; западный звук: сложный тон, содержащий повторяющиеся 14-20 кГц 1.5 раз в секунду). На виртуальном столе не было пространственно информативных подсказок (рис. 1А). Крыса стартовала с одной из 4 случайных стартовых позиций лицом к стене. Северо-восточный квадрант таблицы был обозначен как целевой квадрант. В центре этого квадранта находилась немаркированная зона вознаграждения радиусом 30 см. При входе в эту зону выдавалось до 5 поощрительных импульсов. Чтобы обеспечить визуальную обратную связь, при входе появилась белая точка, охватывающая зону вознаграждения. После выдачи 5 наград или если крыса покинула зону награды, инициировался 2-секундный период затемнения, а затем крыса была телепортирована в одно из 4 случайных начальных мест, обращенных к стене.Крыс запускали максимум 45 минут или 200 импульсов вознаграждения. Крыс обучали в течение 6 дней приобретения с зоной вознаграждения радиусом 30 см. После этого размер зоны вознаграждения был уменьшен до 25 см в течение одного дня, а затем до 20 см в течение нескольких дней до достижения критерия двух дней из 200 импульсов вознаграждения в течение 30 минут. Через 24 часа после последнего дня обучения было проведено испытание с зондом, в котором зона вознаграждения была инактивирована, и животное могло исследовать в течение 4,5 минут. Затем мы разработали задачу для систематического удаления визуальных или слуховых сигналов с использованием блокированного дизайна с 8 попытками аудиовизуальных сигналов, 8 попытками только со слуховыми сигналами и 8 попытками только с визуальными сигналами.В каждом блоке каждое начальное местоположение использовалось дважды в псевдослучайном порядке. Для этого использовали шесть крыс.
Чтобы определить, возможно ли пространственное обучение, основываясь исключительно на дистальных слуховых сигналах, без какого-либо потенциального затенения визуальными сигналами, мы разработали задачу с двумя дистальными слуховыми сигналами и без дистальных визуальных сигналов. Два новых и отчетливых слуховых сигнала были размещены к северо-западу и юго-востоку от виртуального стола, а зона вознаграждения 25 см находилась в центре северо-западного квадранта (северо-западный звук: сложный звук с частотой около 8 кГц, повторяемый три раза в секунду; SW-звук: колеблющаяся развертка. от 2 до 7 кГц повторяется 0.35 раз в секунду, рис. 4A). Эта схема была основана на предыдущем исследовании, которое показало доказательства пространственной навигации, основанной исключительно на дистальных звуковых сигналах [24]. Также была создана виртуальная среда с идентичной компоновкой, за исключением того, что использовались две визуальные реплики вместо двух слуховых. Первоначальное обучение происходило на виртуальном столе, размеры которого были идентичны предыдущей задаче аудиовизуальной пространственной навигации. Для чисто слуховой задачи это не дало никаких доказательств пространственного обучения, а вместо этого указывало на то, что крысы приняли общую стратегию поиска – бег на фиксированное расстояние от края стола.Поэтому, чтобы снизить эффективность этой стратегии, мы увеличили радиус виртуального стола до 1,2 м и уменьшили зону вознаграждения до 20 см, оставив при этом относительное расположение зоны вознаграждения и дистальных слуховых сигналов нетронутыми. Кроме того, из-за большей сложности задачи количество импульсов поощрения было увеличено с 5 до 10. Обучение в виртуальной среде с двумя дистальными зрительными сигналами проходило по той же процедуре, что и обучение в слуховой версии. Данные, представленные на рисунке 4, относятся к последним 4 дням выполнения обеих этих задач для каждой крысы.Для этого были использованы пять крыс, которые ранее были обучены задаче пространственной навигации.
Анализ данных и статистические методы
Порог скорости 2 см / с применялся ко всем показателям занятости, нормализованной скорости проверки и квадрантам, чтобы исключить периоды неподвижности. Что касается показателей производительности, мы рассчитали среднее значение по всем испытаниям для каждой крысы в течение сеанса и выполнили последующий анализ, используя эти значения для всех крыс. Обрезка краев рассчитывалась путем деления общего движения сферической беговой дорожки на границы виртуального стола на общее движение сферической беговой дорожки во время каждой тренировки.Используя метод повторной выборки, мы рассчитали пространственное распределение случайных проверок вознаграждения путем случайного перераспределения количества проверок вдоль пути крысы от одного испытания к другому. Нормализованная мера расстояния проверки была рассчитана путем деления среднего радиального расстояния проверок вознаграждения на среднее радиальное расстояние проверок вознаграждения с повторной выборкой (Фактическое среднее расстояние / Среднее расстояние после повторной выборки x 100). Нормализованная частота проверок рассчитывалась путем деления частоты проверок в каждой ячейке на площадь под кривой скорости проверки по всем ячейкам в пределах 80 см от зоны вознаграждения для каждой крысы.Модуляция частоты проверок выше вероятности была рассчитана путем вычитания нормализованной средней повторной частоты проверок в каждом интервале из нормализованной частоты проверок в каждом интервале, а затем деления на нормализованную среднюю повторную частоту проверок в каждом интервале ((Фактическая нормализованная частота проверок – Нормализованная частота проверок после повторной дискретизации) / Восстановленная нормализованная частота проверки x 100). Для отдельного примера на рисунке 2 E значения F P были получены путем сравнения фактических данных с повторной выборкой проверки трубки вознаграждения. Размер ячейки 4 см х 4 см использовался для всех мер поведения в 2-мерной плоскости.Точно так же для всех мер, которые зависели от расстояния до мер зоны вознаграждения, использовался радиальный размер ячейки 4 см. Пространственное распределение частоты проверок трубки вознаграждения было вычислено с использованием гауссовского сглаживающего ядра на 6 см (двумерные данные) и 2,6 см (радиальные данные) на гистограммах занятости и проверки трубки вознаграждения. Для визуализации двухмерных гистограмм скорости проверки (рис. 2C, E и рис. 4D) мы применили порог занятости 0,25 с на ячейку на каждую крысу. Информационное содержание гистограмм скорости проверки радиальной трубки вознаграждения (в битах) было определено как
., где i – номер интервала, P i – вероятность занятости интервала i, , полученная из o j занятость пространственного интервала j , λ i – средняя частота проверок для ячеек i, и общая средняя частота проверок.Для квадрантного анализа времени занятости, пройденного расстояния и проверки вознаграждения во время выполнения задач пространственного обучения данные изнутри зоны вознаграждения и эквивалентного местоположения в других квадрантах были удалены перед любыми вычислениями. Планки погрешностей на рисунках представляют собой стандартную ошибку среднего, вычисленного для всех крыс. Для статистического анализа мы использовали ANOVA с альфа = 0,05. Когда это было оправдано, за этим следовали апостериорный анализ Тьюки и поправки Бонферони для повторных сравнений измерений.
Дополнительная информация
Рисунок S1.
Мультимодальный, неинвазивный, мультисенсорный аппарат виртуальной реальности. (A) Поперечное сечение системы VR, показывающее общую раму, зеркало, микропроектор, трубку для вознаграждения, удерживающие механизмы для крысы, сферическую беговую дорожку и воздушную подушку. (B) Вид сверху на расположение динамиков, окружающих устройство. (C) Изображение системы VR сзади, когда крыса выполняет виртуальную случайную задачу поиска пищи. (D) Вид виртуальной случайной среды кормодобывания, созданной программным обеспечением.(E) Фотография сделана внутри VR с точки зрения крысы с той же точки, что и в d. (F) Крыса в системе VR. Обратите внимание на петли и ремни, которые позволяют крысе принимать естественную позу.
https://doi.org/10.1371/journal.pone.0080465.s001
(TIF)
Рисунок S2.
Крысы быстро учатся ориентироваться и избегать краев в конечной двумерной виртуальной среде. (A) Схема для: Сеанса 1 в виртуальной среде Большого стола с 5 наградами (LT5) и Сессии 11 в виртуальной среде Малого стола с 1 наградой (ST1).(B) Пример 30-минутного пути от одной крысы в сеансах 1 и 11 соответственно. (C) Средняя занятость для всех крыс в сеансе 1 и 11 соответственно. (D) Кривая регистрации процента расстояния, пройденного до края платформы, называемая обрезкой края. Эффект сеанса: F (2,32) = 4,037, p = 0,0038, первый сеанс против пятого: t = 2,812, p <0,05, N = 4. (E) Кривая сбора данных для задержки между вознаграждениями. Эффект сеанса: F (2,32) = 3,913, p = 0,0012, первый сеанс против четвертого: t = 3.056, р <0,05.
https://doi.org/10.1371/journal.pone.0080465.s002
(TIF)
Рисунок S3.
Проверка системы объемного звучания с семью динамиками. (A) Схема задач визуального и звукового маяка. Стрелки указывают случайную начальную ориентацию крыс в каждом испытании. Полосатый кружок указывает на наличие визуального маяка. Значок звука указывает на наличие звукового маяка. (B) Примеры путей для двух пробных типов. Цвет каждого пути указывает приблизительную начальную ориентацию, цвет обозначен стрелками в a.На этом и на всех последующих графиках области в зоне вознаграждения не анализируются. (C) 2-мерная гистограмма средней занятости, усредненной по всем крысам. (D) 2-мерная гистограмма нормализованной частоты проверок, усредненной по крысам. Белые бункеры не получили достаточной выборки. (E) Средняя задержка и расстояние до вознаграждения в визуальных (V) и слуховых (A) задачах. Эффект типа исследования: t = 2,453, p = 0,070 и t = 3,945, p = 0,0169 соответственно; A в сравнении с AV и V, p <0,05, N = 3. (F) Интенсивность звука (дБ) на расстоянии от источника звука в VR и RW.(G) Интенсивность звука (дБ) источника звука при различных ориентациях в VR.
https://doi.org/10.1371/journal.pone.0080465.s003
(TIF)
Видео S2.
На этом видео показана крыса, выполняющая задачу виртуальной пространственной навигации. Крысы начинают игру в одном из четырех случайных начальных мест, обращенных в противоположную сторону от центра виртуальной платформы, а затем должны перейти к месту получения награды на основе отдаленных сигналов. После получения награды наступает двухсекундный период затемнения, а затем крыса телепортируется в одно из четырех случайных начальных мест.
https://doi.org/10.1371/journal.pone.0080465.s006
(MP4)
Благодарности
Мы хотели бы поблагодарить Джейсона Мура, Чжипинга Чена и Дэвида Хо за их квалифицированную техническую помощь.
Заявление об ограничении ответственности: Эти результаты были опубликованы в виде тезисов на ежегодном собрании Общества нейробиологии, рефераты № 812.01 и 812.10 (2012).
Вклад авторов
Задумал и спроектировал эксперименты: JDC DBA MRM.Проведены эксперименты: Джойнт ПР АК ЦВ БП. Проанализированы данные: DBA JDC. Написал статью: JDC DBA MRM. Создал оборудование для этих экспериментов: DBA BW. Написал программу для этих экспериментов: DBA BW. Участвовал в строительстве метизов: КА MRM.
Ссылки
- 1. White NM, McDonald RJ (2002) Множественные параллельные системы памяти в мозге крысы. Neurobiol Learn Mem 77: 125–184.
- 2. Моррис Р.Г., Гарруд П., Роулинз Дж. Н., О’Киф Дж. (1982) У крыс с поражениями гиппокампа нарушена навигация по месту.Природа 297: 681–683.
- 3. Balsam PD, Drew MR, Gallistel CR (2010) Время и ассоциативное обучение. Comp Cogn Behav Rev 5: 1-22.
- 4. Петерсон Г.Б., Акилт Дж. Э., Фроммер Г. П., Херст Е. С. (1972) Обусловленный подход и контактное поведение в отношении сигналов для питания или подкрепления стимуляцией мозга. Наука 177: 1009–1011.
- 5. Schultz W, Apicella P, Scarnati E, Ljungberg T (1992) Нейрональная активность в вентральном полосатом теле обезьяны, связанная с ожиданием вознаграждения.J Neurosci 12: 4595–4610.
- 6. Pan WX, Brown J, Dudman JT (2013) Нейронные сигналы угасания в тормозной микросхеме вентрального среднего мозга. Nat Neurosci 16: 71–78.
- 7. Holscher C, Schnee A, Dahmen H, Setia L, Mallot HA (2005) Крысы могут перемещаться в виртуальных средах. J Exp Biol 208: 561–569.
- 8. Harvey CD, Collman F, Dombeck DA, Tank DW (2009) Внутриклеточная динамика пространственных клеток гиппокампа во время виртуальной навигации.Природа 461: 941–946.
- 9. Dombeck DA, Harvey CD, Tian L, Looger LL, Tank DW (2010) Функциональная визуализация клеток места гиппокампа при клеточном разрешении во время виртуальной навигации. Nat Neurosci 13: 1433–1440.
- 10. Harvey CD, Coen P, Tank DW (2012) Последовательности, зависящие от выбора, в теменной коре головного мозга во время задачи решения виртуальной навигации. Природа 484: 62–68.
- 11. Келлер Г.Б., Бонхёффер Т., Хубенер М. (2012) Сигналы сенсомоторного несоответствия в первичной зрительной коре головного мозга ведущей мыши.Нейрон 74: 809–815.
- 12. Равассард П. К., Виллерс Б., Хо Д., Ахарони Д. А., Кушман Д. Д. и др. (2013) Мультисенсорный контроль пространственно-временной селективности гиппокампа. Наука в прессе.
- 13. Chen G, King JA, Burgess N, O’Keefe J (2013) Как видение и движение сочетаются в коде места гиппокампа. Proc Natl Acad Sci U S A 110: 378–383.
- 14. Янгстром И.А., Строубридж Б.В. (2012) Визуальные ориентиры облегчают пространственную навигацию грызунов в средах виртуальной реальности.Выучите Mem 19: 84–90.
- 15. Моррис Р. (1984) Разработка процедуры водного лабиринта для изучения пространственного обучения у крыс. J Neurosci Methods 11: 47–60.
- 16. Mackintosh NJ (1976) Затенение и интенсивность стимула. Обучение и поведение животных 4: 186–192.
- 17. Bohbot VD, Lerch J, Thorndycraft B, Iaria G, Zijdenbos AP (2007) Различия серого вещества коррелируют со спонтанными стратегиями в задаче виртуальной навигации человека.Журнал неврологии 27: 10078–10083.
- 18. Джейкобс WJ, Laurance HE, Thomas KGF (1997) Место обучения в виртуальном пространстве I: приобретение, затенение и передача. Обучение и мотивация 28: 521–541.
- 19. Макнотон Б.Л., Батталья Ф.П., Дженсен О., Мозер Е.И. и др. (2006) Интеграция путей и нейронная основа «когнитивной карты». Nat Rev Neurosci 7: 663–678.
- 20. Muller RU, Poucet B, Fenton AA, Cressant A (1999) Является ли гиппокамп крысы частью специализированной навигационной системы? Гиппокамп 9: 413–422.
- 21. Эйхенбаум Х. (2000) Гиппокамп: отображение или память? Curr Biol 10: R785–787.
- 22. Eichenbaum H, Stewart C, Morris RG (1990) Репрезентация гиппокампа в обучении на месте. J Neurosci 10: 3531–3542.
- 23. Россье Дж., Хэберли С., Шенк Ф. (2000) Слуховые сигналы поддерживают навигацию по месту у крыс, когда они связаны с визуальным сигналом. Behav Brain Res 117: 209–214.
- 24. Сазерленд Р.Дж. (1984) Навигация с помощью крыс в бассейне.Канадский журнал психологии 38: 322–347.
- 25. Ховард И. П., Темплтон В. Б. (1966) Ориентация человека в пространстве. Лондон; Нью-Йорк: Вили. 533 с. п.
- 26. Welch RB, Warren DH (1980) Непосредственный перцептивный ответ на межсенсорное несоответствие. Psychol Bull 88: 638–667.
- 27. Эрнст М. О., Бултхофф Х. Х. (2004) Объединение органов чувств в устойчивое восприятие. Тенденции Cogn Sci 8: 162–169.
- 28. Талсма Д., Сеньковски Д., Сото-Фарако С., Волдорф М.Г. (2010) Многогранное взаимодействие между вниманием и мультисенсорной интеграцией.Тенденции Cogn Sci 14: 400–410.
- 29. Heffner RS, Heffner HE (1992) Визуальные факторы в локализации звука у млекопитающих. J Comp Neurol 317: 219–232.
- 30. Фанселоу М.С. (2000) Контекстуальный страх, гештальт-воспоминания и гиппокамп. Behav Brain Res 110: 73–81.
- 31. Руди Дж. У., Сазерленд Р. Дж. (1989) Формирование гиппокампа необходимо крысам для обучения и запоминания конфигурационных различений. Behav Brain Res 34: 97–109.
- 32.Пирс Дж. М. (2002) Оценка и развитие коннекционистской теории конфигурационного обучения. Anim Learn Behav 30: 73–95.
- 33. Галлистель ЧР (1990) Организация обучения. Кембридж, Массачусетс: Bradfor / MIT Press.
- 34. О’Киф Дж, Надел Л. (1978) Гиппокамп как когнитивная карта. Оксфорд: Издательство Оксфордского университета.
- 35. Ченг К. (1986) Чисто геометрический модуль в пространственном представлении крысы. Познание 23: 149–178.
- 36. Dupret D, O’Neill J, Pleydell-Bouverie B, Csicsvari J (2010) Реорганизация и реактивация карт гиппокампа предсказывают производительность пространственной памяти. Nat Neurosci 13: 995–1002.
- 37. Hollup SA, Molden S, Donnett JG, Moser MB, Moser EI (2001) Поместите поля пирамидных клеток гиппокампа крысы и пространственное обучение в водном лабиринте. Eur J Neurosci 13: 1197–1208.
- 38. Ито Р., Роббинс Т.В., Макнотон Б.Л., Эверитт Б.Дж. (2006) Селективные эксайтотоксические поражения гиппокампа и базолатеральной миндалины оказывают диссоциативное влияние на сигнал аппетита и кондиционирование места на основе интеграции путей в новой процедуре Y-лабиринта.Eur J Neurosci 23: 3071–3080.
- 39. Мехта М.Р., Макнотон Б.Л. (1997) Расширение и сдвиг пространственных полей гиппокампа: доказательства синаптического потенциала во время поведения. Нью-Йорк, штат Нью-Йорк: Plenum Press.
- 40. Mehta MR, Barnes CA, McNaughton BL (1997) Зависимое от опыта асимметричное расширение полей мест гиппокампа. Proc Natl Acad Sci U S A 94: 8918–8921.
- 41. Mehta MR, Quirk MC, Wilson MA (2000) Зависимая от опыта асимметричная форма рецептивных полей гиппокампа.Нейрон 25: 707–715.
- 42. Astur RS, Ortiz ML, Sutherland RJ (1998) Характеристика работы мужчин и женщин в виртуальном водном задании Морриса: большая и достоверная разница между полами. Behav Brain Res 93: 185–190.
- 43. Экстром А.Д., Букхаймер С.Ю. (2007) Восстановление пространственной и временной эпизодической памяти задействует диссоциированные функциональные сети в человеческом мозге. Обучение и память 14: 645–654.
- 44. Doeller CF, King JA, Burgess N (2008) Параллельные стриатальные и гиппокампальные системы для ориентиров и границ в пространственной памяти.Proc Natl Acad Sci U S A 105: 5915–5920.
