
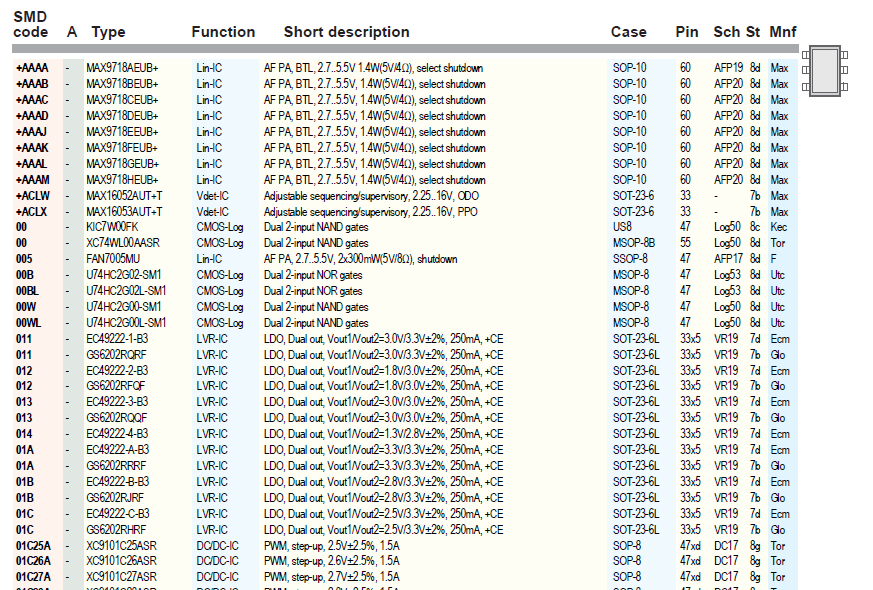
Сокращенная маркировка SMD радиодеталей (marking SMD) | hardware
Типы миниатюрных SMD-компонентов, коварно закодированные производителями трехсимвольной и двухсимвольной (а иногда кодировка состоит из одного символа!) маркировкой, без специальных справочников распознать очень непросто. У меня накопилось несколько ссылок на онлайновые справочники такого рода кодировки, и решил их для удобства выложить в виде отдельного обзора.
http://www.s-manuals.com/smd
Довольно удобный справочник, оформленный в виде квадратной таблицы по двум первым символам кодировки. В ячейках таблицы находятся ссылки на более детальную таблицу, в которой имеется наименование и назначение радиокомпонента, его производитель и даже ссылка на даташит.
http://microsin.ru/phpscr/showsmd02.php
Справочник из журнала Радиокомпоненты”, 1..4 номера 2003 г. и 1, 2 номера 2004 г. – таблица, удобная для поиска по загруженной странице в браузере.
The SMD Codebook
Справочник построен из набора таблиц, каждая таблица соответствует первому символу кодировки. Указаны наименование компонента, производитель, код картинки с цоколевкой, тип корпуса, краткое описание (или эквивалент). Есть размеры многих SMD-корпусов.
Surface Mount Device identification
Справочник, не такой полный, как другие, но тоже достойный внимания. Указаны принципы маркировки SMD резисторов и конденсаторов.
[Ссылки]
1. Таблицы соответствия микросхем 561 и 1561 серий импортным микросхемам 4000 серии.
 Таблица соответствия отечественных микросхем серий TTL импортным микросхемам 74-й серии.
Таблица соответствия отечественных микросхем серий TTL импортным микросхемам 74-й серии.3. Мини-справочник по микросхемам.
4. Сокращенная кодировка компонентов Analog Devices.
Микросхемы в корпусах SOT26 и SOT23-6 и их маркировка.
В современной электронике множество электронных компонентов производится в миниатюрных корпусах TSOP6, SSOT6, SOT23-6, SOT23-5, SOT26. В связи с малыми габаритами радиодеталей в данных корпусах, производитель, зачастую, маркирует компоненты кодовым обозначением. В сервисных центрах и ремонтных мастерских возникают трудности при опознании неисправных электронных компонентов с кодовой маркировкой.
Следующая таблица поможет для опознания парт номера электронного компонента по его зашифрованной кодовой маркировке, для дальнейшего поиска документации (DataSheet) и подбору аналога.
В таблице представлены ШИМ контроллеры, DC/DC преобразователи в пяти и шести выводных SMD корпусах SOT23-5, SOT26, SOT23-6, TSOP6.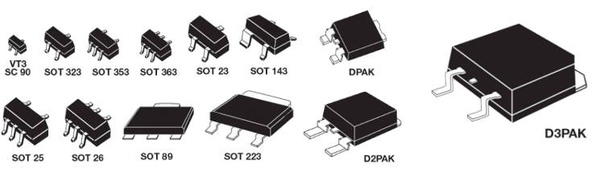
| Полный партномер | Производитель | Маркировка на корпусе |
| ACT511US | Active-Semi | FSGT |
| AOZ1083CI | Alpha & Omega Semiconductor | BA… |
| AP3012K | BCD Semiconductor | E6B |
| AP3031K | BCD Semiconductor | GEC |
| AP3103K | BCD Semiconductor | GHL |
| APW7077AR | Anpec | A77S |
| APW7077AZ | Anpec | A77B |
| FAN6862 | Fairchild | AB… |
| FSP3307 | Foslink Semiconductor | GEC |
| GR8830 | Grenergy | 30… |
| GR8836 | Grenergy | 36… |
| HT2263MP | HotChip | 63… |
| HT2273 | HotChip | 73… |
| HX1001-AE | Hexin | HX-VG |
| IN1M101-T6G | Infinno | M101 |
| IRMLS1503 | Micro6 | 2B… |
| IRMLS1503 | Micro6 | B… |
| IRMLS1902 | Micro6 | 2A… |
| IRMLS1902 | Micro6 | A… |
| IRMLS2002 | Micro6 | 2G… |
| IRMLS2002 | Micro6 | G… |
| IRMLS4502 | Micro6 | 2F… |
| IRMLS4502 | Micro6 | F… |
| IRMLS5703 | Micro6 | 2D… |
| IRMLS5703 | Micro6 | D… |
| IRMLS6702 | Micro6 | 2C… |
| IRMLS6702 | Micro6 | C… |
| IRMLS6802 | Micro6 | 2E… |
| IRMLS6802 | Micro6 | E… |
| IRMLS6803 | Micro6 | 2H… |
| IRMLS6803 | Micro6 | H… |
| L2263TP | Shenzhen LIZE Electronic Technology | L63… |
| LD7510GL | Leadtrend | .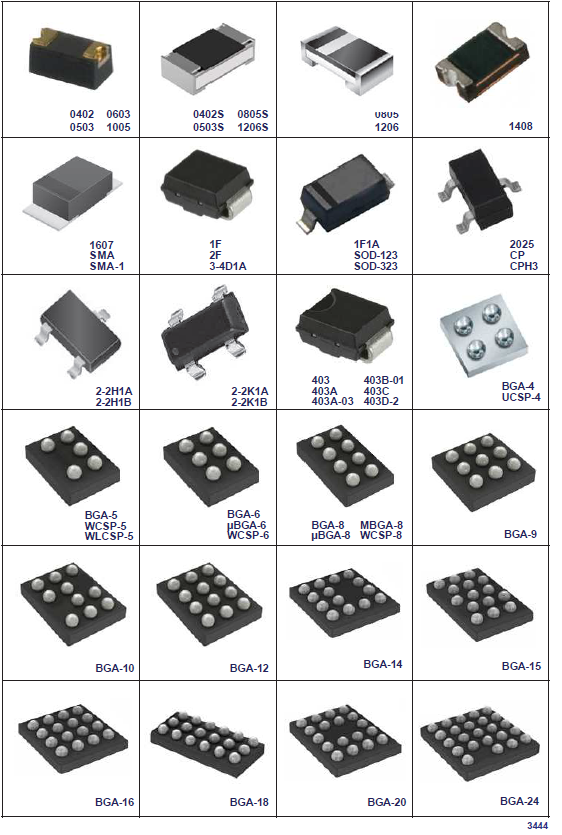 ..P/10 ..P/10 |
| LD7510JGL | Leadtrend | …P/10J |
| LD7513GL | Leadtrend | …P/13 |
| LD7530 | Leadtrend | …30 |
| LD7530 | Leadtrend | …P/30 |
| LD7530A | Leadtrend | …30A |
| LD7530A | Leadtrend | …P/30A |
| LD7530APL | Leadtrend | sL01 |
| LD7531A | Leadtrend | 31A |
| LD7531GL | Leadtrend | …P/31 |
| LD7531PL | Leadtrend | …P/31 |
| LD7535 | Leadtrend | 35. |
| LD7535A | Leadtrend | 35A |
| LD7535BL | Leadtrend | S12W |
| LD7535IL | Leadtrend | …P/35 |
| LD7536R | Leadtrend | 36R |
| LD7536RGL | Leadtrend | …P/36R |
| LD7537R | Leadtrend | 37R |
| LD7550BL | Leadtrend | . ..P/50 ..P/50 |
| LD7550IL | Leadtrend | …P/50 |
| LP2273 | LP2273 | |
| MP2104DJ | Monolithic Power Systems | C2 |
| NCP1250ASN | ON Semiconductor | 25A |
| NCP1250BSN | ON Semiconductor | 252. |
| NCP1251 | ON Semiconductor | 5AA… |
| NCP1251 | ON Semiconductor | 5A2… |
| NCP1251 | ON Semiconductor | 5AC… |
| NCP1251 | ON Semiconductor | 5AD… |
| NCP1251 | ON Semiconductor | 5AF… |
| NCP1251 | ON Semiconductor | 5UA… |
| NCP1251 | ON Semiconductor | 5U2… |
| NCP1251 | ON Semiconductor | 5UC… |
| NCP1251 | ON Semiconductor | 5UD… |
| NCP1251 | ON Semiconductor | 5UF… |
| NCP1400ASN25 | ON Semiconductor | DAV… |
| NCP1400ASN33 | ON Semiconductor | DAJ… |
| NCP1529ASN | ON Semiconductor | DXJ… |
| OB2252MP | Lite-on | 52… |
| OB2262MP | Lite-on | 62… |
| OB2263MP | Lite-on | 63… |
| OB2273MP | Lite-on | 73… |
| OB2361MP | Lite-on | 61… |
| PAM2301 | Diodes Incorporated | BA… |
| PT4101 | Powtech | 4101.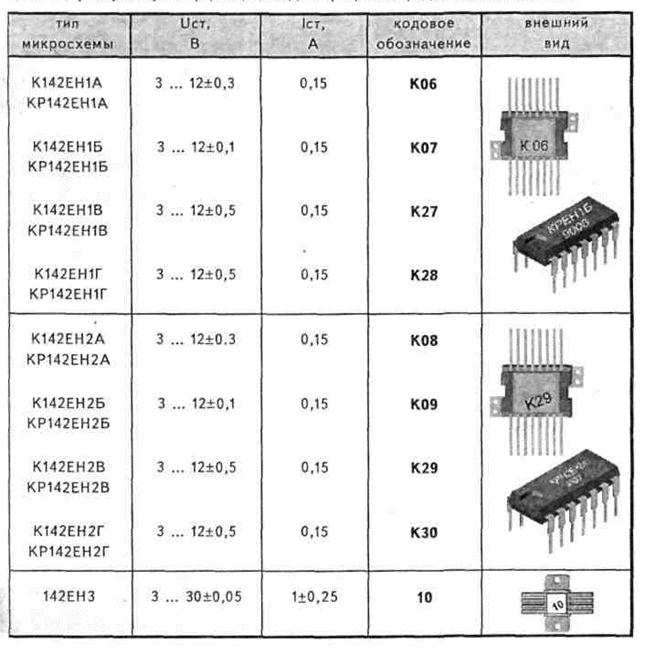 |
| PT4103 | Powtech | 4103. |
| PT4201E23F | Powtech | 4201. |
| R7731A | Richtek Technology Corporation | IDP=… |
| R7731AGE | Richtek Technology Corporation | IDP=… |
| R7731GE | Richtek Technology Corporation | 0Q= |
| R7731PE | Richtek Technology Corporation | 0Q= |
| SD4870TR | Silan Microelectronics | 4870. |
| SD4871TR | Silan Microelectronics | 4871. |
| SD6855 | Silan Microelectronics | 6855. |
| SD6856 | Silan Microelectronics | 6856. |
| SF1530LGT | SiFirst | 30… |
| SF1531LGT | SiFirst | 31… |
| SG5701TZ | System General Corp. | AAE… |
| SG6848T | System General Corp. | AAHBW |
| SG6848T | System General Corp. | Ea… |
| SG6848T | System General Corp. | AAHB… |
| SG6848T | System General Corp. | EA… |
| SG684965TZ | System General Corp. | BB |
| SG6858TZ | Fairchild | AAI… |
| SG6859ATZ | Fairchild | AAJF… |
| SG6859TZ | Fairchild | AAJ.. |
| SG6860TY | Fairchild | AAQ… |
| SGP400 | Fairchild | AAK… |
| SP6850AS26RG | Sync Power | 85A… |
| SP6850BS26RG | Sync Power | 850… |
| SP6853S26RGB | Sync Power | 853… |
| STC4054GR | STMicroelectronics | LTH7 |
| SY8009A | Silergy Corp. | AD… |
| SY8009B | Silergy Corp. | CU… |
| UC3863G | Unitrode | U863 |
| UC3863L | Unitrode | U863 |
| UC3873G | Unitrode | U873 |
| UC3873L | Unitrode | U873 |
| XN1049TP | 49… |
Маркировка радиоэлементов (импортных, активных) « РадиоГазета – принципиальные схемы для меломанов и аудиофилов
Опубликовано: 18 сентября, 2017 • Рубрика: Разное
В последние годы производители полупроводников оптимизировали номенклатуру своих изделий, и количество предлагаемых устройств несколько сократилось. Однако, это трудно заметить при просмотре каталогов компонентов, где количество различных устройств только одного типа может составлять не менее нескольких сотен. Для крупного, профессионального поставщика в каталогах будет доступно несколько тысяч полупроводников.
Однако, это трудно заметить при просмотре каталогов компонентов, где количество различных устройств только одного типа может составлять не менее нескольких сотен. Для крупного, профессионального поставщика в каталогах будет доступно несколько тысяч полупроводников.
Именно поэтому при подборе элементов даже опытным радиоинженерам следует проявлять осторожность, потому что легко ошибиться, когда имеется так много компонентов одного типа, многие из которых имеют схожую маркировку. Иначе вы рискуете купить неправильный прибор/компонент или правильный компонент, но неправильную его версию.
Анатомия маркировки
Ошибок не будет, если вы понимаете основную анатомию маркировки полупроводникового компонента. Конечно, всех проблем это не решит, но три составные части маркировки надо знать обязательно.
Обычно в маркировке есть префикс, который предоставляет некоторую базовую информацию об устройстве, но используемые методы кодирования очень просты и никогда не рассказывают вам о конкретном устройстве. Однако при покупке компонентов префикс может быть (и довольно часто) очень важен.
Однако при покупке компонентов префикс может быть (и довольно часто) очень важен.
Вторая часть является основной (как бы серийный номер изделия) и имеет три или четыре цифры.
Третья часть – суффикс, предоставляет некоторую дополнительную информацию об устройстве, но он не всегда присутствует, особенно у транзисторов и диодов. Он необходим только при наличии двух или более разных версий устройства.
Опять же, это важно при покупке компонентов, и вы можете легко получить неправильную версию, если у устройства будет неправильный суффикс. Есть много примеров идентичных устройств, которые имеют разные суффиксы.
Менеджмент «среднего звена»
Основная часть – это наиболее простая часть маркировки полупроводниковых элементов. Первое устройство такого типа, которое должно быть зарегистрировано, может иметь номер «0001», следующий — «0002» и т. д.
На практике это работает не совсем так, и некоторые производители транзисторов начинают маркировку своих изделий с «100», а не «001».
Существенным недостатком такого метода маркировки является наличие большего числа полупроводниковых приборов, чем доступных номеров (3-х или 4-х значных).
Для примера, устройство, промаркированное «555», может быть популярной интегральной схемой таймера (ИС), транзистором с европейским типом номера и, возможно, чем-то другим, например, другим типом интегральной схемы или оптическим устройством.
Таким образом, базовая числовая маркировка важна, но сама по себе недостаточна для точной идентификации элемента.
Чтобы выбрать подходящий элемент нужно обязательно обращать внимание и на другие части маркировки.
Начать с начала
Первая часть маркировки (префикс) выполняет две функции, и для европейских производителей эта часть маркировки даёт некоторую базовую информацию о типе устройства. Она чем-то похожа и берёт истоки у маркировки вакуумных ламп, но применительно к твёрдотельным устройствам первая буква указывает на тип используемого полупроводникового материала или тип интегральной схемы:
| Первый символ | Тип элемента |
| A | Германий |
| B | Кремний |
| C | Арсенид галия |
| F | Интегральная логическая схема |
| R | Фотоэлемент |
| S | Цифровая интегральная схема |
| T | Линейная интегральная схема |
Вторая буква указывает тип устройства, так как в таблице 2.
| Второй символ | Тип прибора |
| A | Сигнальный диод |
| B | Варикап |
| C | Маломощный транзистор для аудио |
| D | Мощный транзистор для аудио |
| E | Туннельный диод |
| F | Маломощный высокочастотный транзистор |
| L | Мощный высокочастотный транзистор |
| P | Фототранзистор |
| S | Транзистор для ключевых схем |
| T | Тиристор |
| Y | Фильтр |
| Z | Стабилитрон |
Заметим, что элементы для промышленных применений имеют в маркировке три буквы.
Для примера, BC550 представляет собой небольшой кремниевый транзистор для аудио или других низкочастотных приложений, в то время как BF181 представляет собой маломощный кремниевый транзистор для использования на радиочастотах.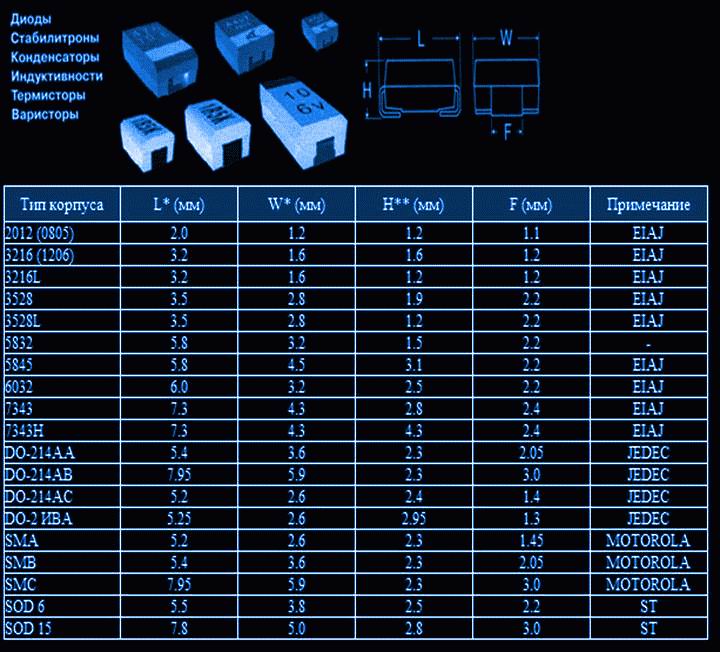
На один меньше
Простые полупроводники американских производителей маркируются по системе JEDEC (Joint Electron Devices Engineering Council) и имеют префикс, состоящий из цифры, за которой следует буква N . Цифра на единицу меньше количества выводов, которое имеет устройство, что на практике означает 1 — для диодов и стабилитронов (т.е. два вывода), «2» для обычных транзисторов и «3» или более для специальных устройств, таких как двухзатворные МОП-транзисторы и прочее.
Таким образом, 1N4148 является устройством, которое имеет два вывода, что обычно означает диод. Это на самом деле небольшой диод, но эта информация не отображается в маркировке типа JEDEC, которая получается менее информативна, чем европейская Pro Electron.
Сейчас не часто встречается маркировка японской системы JIS (Японские промышленные стандарты), но первая цифра в ней снова является числом, которое на один меньше, чем количество выводов у элемента. Затем следуют две буквы, которые идентифицируют общий тип устройства:
| Маркировка | Тип устройства |
| SA | Высокочастотный PNP транзистор |
| SB | Высокочастотный NPN транзистор |
| SC | PNP транзистор для аудио |
| SD | NPN транзистор для аудио |
| SE | Диод |
| SJ | P-канальный полевой транзистор (в том числе и MOSFET) |
| SK | N-полевой транзистор (в том числе и MOSFET) |
| SR | Фильтр |
Как нетрудно заметить, для обычных типов транзисторов первые две цифры всегда получаются «2S» и, возможно, они немного бесполезны, поэтому эти две цифры часто опускаются при маркировке элементов.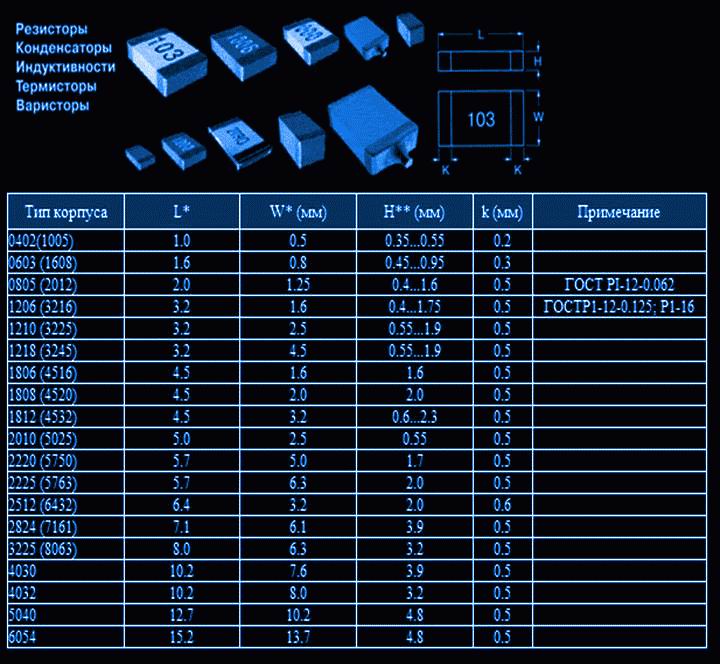
Производитель
Большинство электронных компонентов маркируются согласно перечисленным стандартным методам. Но бывают и исключения. (рис.1).
Здесь префикс TIP этого силового транзистора указывает, что он является мощным транзистором в пластиковом корпусе от Texas Instruments. Однако впереди производитель нанёс логотип MOSPEC, поэтому префикс стал вторым элементом маркировки.
Такое часто встречается в маркировке интегральных микросхем, где к стандартной маркировке типа производитель добавляет свою кодировку.
Рис.2. Эта интегральная схема имеет обозначение «LM» в качестве префикса, что указывает на то, что это изделие фирмы National Semiconductor.
Как несколько примеров: префиксы «CA» и «MC» используются соответственно фирмы KCA и Motorola. Из-за того, что один и тоже элемент может выпускаться разными производителями и маркироваться по своему, возникают трудности с идентификацией элементов.
Конечно, наличие на рынке нескольких производителей порождает конкуренцию, что, как следствие, снижает цены на радиоэлементы.
При просмотре каталога интегральных микросхем, вероятно, лучше всего игнорировать префикс и сосредоточиться на двух других элементах маркировки. Тем более, что часто поставщики компонентов не гарантируют поставку устройств от конкретных производителей. Если вы заказываете (скажем) MC1458CP. но вам прислали СА1458Е. или наоборот, нет повода беспокоиться. Обе микросхемы являются 1458 — двойными операционными усилителями, и нет никакой практической разницы между ними. MC1458CP производится Motorola или Texas Instruments, а СА1458Е – фирмой RCA.
Полный список префиксов производителей смотрите на сайте: https://en.wikibooks.org/wiki/Practical_Electronics/Manufacturers_Prefix
Многообразие вариантов
Большинство транзисторов не имеют суффикса в маркировке. Там, где он присутствует, суффикс обычно представляет собой одну букву и указывает на коэффициент усиления или другой какой-то параметр. Обычно буквой «А» маркируются транзисторы с низким коэффициентом усиления, буквой «В» со средним и буквой «С» с высоким коэффициентом усиления. Конкретные значения или диапазон указывается в даташите на элемент.
Обычно буквой «А» маркируются транзисторы с низким коэффициентом усиления, буквой «В» со средним и буквой «С» с высоким коэффициентом усиления. Конкретные значения или диапазон указывается в даташите на элемент.
Поэтому, если на схеме указан транзистор с суффиксом «В», заменить его безопасно можно на транзистор с суффиксом «С». При замене на элемент с суффиксом «А» может не хватить его усиления и устройство откажется работать или будет часто уходить в перегрузку.
Бывают ситуации (к счастью, довольно редкие), когда суффикс указывает на расположение выводов элемента. Для транзисторов это обозначения «L» или «K». Большинство транзисторов имеют одну типовую конфигурацию выводов. Но если ваше устройство не работает по непонятным причинам, проверьте, не попались ли вам транзисторы с такими суффиксами.
С интегральными микросхемами ситуация противоположная. Тут производители часто используют суффикс для обозначения типа корпуса. И если вы при заказе проигнорируете суффикс или укажите неверный, вы рискуете получить микросхему в таком исполнении, которое будет не совместимо с вашим вариантом печатной платы.
Ситуация осложняется тем, что стандартов на суффиксы нет и каждый производитель использует свои типы маркировки. Так что будьте предельно внимательны при заказе микросхем!
Маркировка частоты
Некоторые интегральные схемы имеют суффикс, который указывает на тактовую частоту устройства. Эта система используется совместно с памятью и некоторыми другими компьютерными чипами, такими как микроконтроллеры и микропроцессоры. В большинстве случаев дополнительные цифры на самом деле являются расширением основной части маркировки, а не суффиксом, так как в маркировке суффикс будет присутствовать и, как говорилось выше, скорее всего будет обозначать тип корпуса.
Некоторые микроконтроллеры PIC, например, имеют в обозначении что-то вроде « -20», добавленное к базовому типу номера. Дополнительная маркировка указывает максимальную тактовую частоту (в мегагерцах) для чипа. Вы можете вполне безопасно использовать элемент с более высокой тактовой частотой, чем тот, который указан в списке компонентов.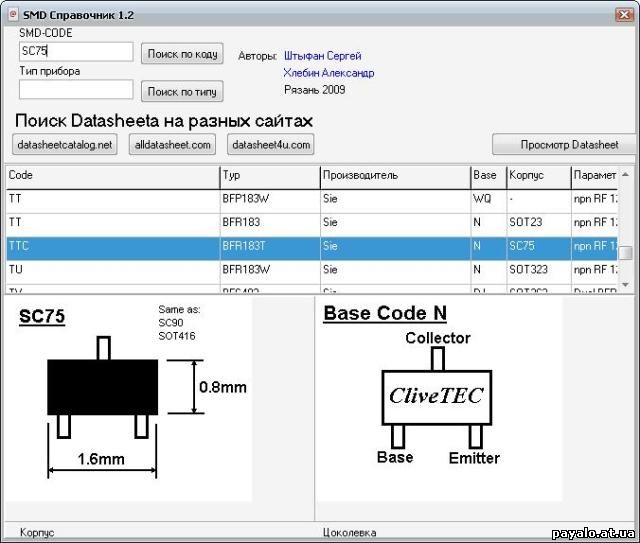 Однако, более быстрые версии, как правило, значительно дороже, чем медленные.
Однако, более быстрые версии, как правило, значительно дороже, чем медленные.
И технологии…
Но, увы, не всё так просто. Особенно с интегральными микросхемами. 74-я серия (TTL) логических интегральных схем была основной, прародительницей других серий и первоначально маркировалась по изложенным правилам: префикс-основная часть-суффикс. При маркировке последующих, улучшенных серий, от стандартной маркировки производители начали отклоняться — между префиксом «74» и базовым номером стали добавлять маркировку, обозначающую семейство микросхем:
Эта маркировка может указывать на технологию изготовления и, как следствие, на скорость (частоту), напряжения питания и другие параметры.
Поэтому исходное устройство 7420 сегодня может маркироваться как 74HC20, 74MCT20 и 74LS20. Это всё различные семейства микросхем, которые несовместимы между собой. Поэтому и тут при заказе важно выбрать правильный тип!
И тока!
Подобная ситуация есть и у всенародно любимых интегральных стабилизаторов L78XX и L79XX. Здесь к базовому обозначению добавляются две цифры, указывающие на выходное напряжение стабилизаторов: L7805 — выходное напряжение 5В, L7912 — выходное напряжение -12В.
Здесь к базовому обозначению добавляются две цифры, указывающие на выходное напряжение стабилизаторов: L7805 — выходное напряжение 5В, L7912 — выходное напряжение -12В.
Но в середине номера могут присутствовать буквы, которые обозначают максимальный выходной ток стабилизатора. Возможны три варианта маркировки, как представлено в таблице:
| Символ | Максимальный ток |
| L | 0.1 A (100mA) |
| M | 0.5A (500mA) |
| S | 2A |
Так стабилизатор с маркировкой «78L15» будет выдавать на выходе напряжение 15В и максимальный ток 100мА.
Проявляйте внимательность при чтении каталогов производителей и соблюдайте осторожность при заказе радиоэлектронных элементов!
Статья подготовлена по материалам журнала «Практическая электроника каждый день»
Автор: Роберт Пенфолд
Вольный перевод: Главный редактор «РадиоГазеты»
Похожие статьи:
Что такое даташит (datasheet)? Поиск информации о радиодеталях.
Ищем данные о характеристиках радиодеталей
При ремонте и конструировании современной радиоэлектронной аппаратуры очень часто возникает необходимость в информации о конкретных радиоэлементах: диодах, транзисторах, микросхемах и многих других деталях.
Производством и разработкой электронных компонентов занимаются сотни различных фирм, а предлагаемый ассортимент постоянно увеличивается и обновляется.
В настоящее время рынок радиоэлектронных компонентов заполнен разношёрстным импортом. Каких только обозначений не встретишь на корпусах современных радиоэлементов: 2SB764, LA78040, BA1404, LM1117, SN74HC05N, 1N5822, PAM8403, CD5954, MC34063AP, список можно продолжать до бесконечности.
Как же не запутаться в этих цифро-кодовых обозначениях и найти информацию по конкретному компоненту?
Для опытных радиолюбителей это не проблема. Для начинающих электронщиков разобраться в том, что же скрывается в миниатюрном пластиковом корпусе с непонятной маркировкой порой не так-то просто.
Узнать подробную информацию об электронном компоненте можно из его “даташита” (от англ. – datasheet). Он же справочный лист, техническая документация или описание электронного компонента или изделия. В нём приводятся все характеристики прибора, например, для транзистора – тип проводимости, цоколёвка, тип корпуса, размеры, кодовое обозначение, приводятся всевозможные характеристики, графики зависимостей и многое другое. Имея подробную информацию о радиоэлементе можно быстрее найти ему замену .
Особенно важна информация по современным микросхемам. В описании, как правило, приводятся стандартные схемы включения с обозначением номиналов и параметров элементов обвязки. Также указывается сферы применения данной микросхемы и её особенности. Для начинающего радиолюбителя такая информация крайне важна, поскольку позволяет понять назначение и функционал микросхемы, узнать её схему включения, величину номинального и максимального питающего напряжения, назначение выводов и т.д.
Умение работать с технической документацией, это одно из важных качеств специалиста, работающего с электронной техникой.
Где же можно найти описания (datasheet) для радиодеталей?
Очень большое количество описаний всевозможных полупроводников можно найти на сайте www.alldatasheet.com
На момент написания статьи на сайте доступно более 20 миллионов описаний радиоэлементов. Каждый месяц база пополняется более чем на 30 000 описаний! В сутки ресурс обрабатывает более 370 000 поисковых запросов пользователей!
Было бы глупо не воспользоваться возможностями такого мощного сайта.
Как же пользоваться данным сайтом?
Зайдя на главную страницу сайта, мы увидим поле ввода поискового запроса.
К примеру, вводим в поисковую форму – PB137 и жмём кнопку Поиск (Search).
Поиск выдал нам два результата.
Далее жмём на значке . Откроется новая страница.
На новой странице щёлкаем по изображению, которое выглядит как документ.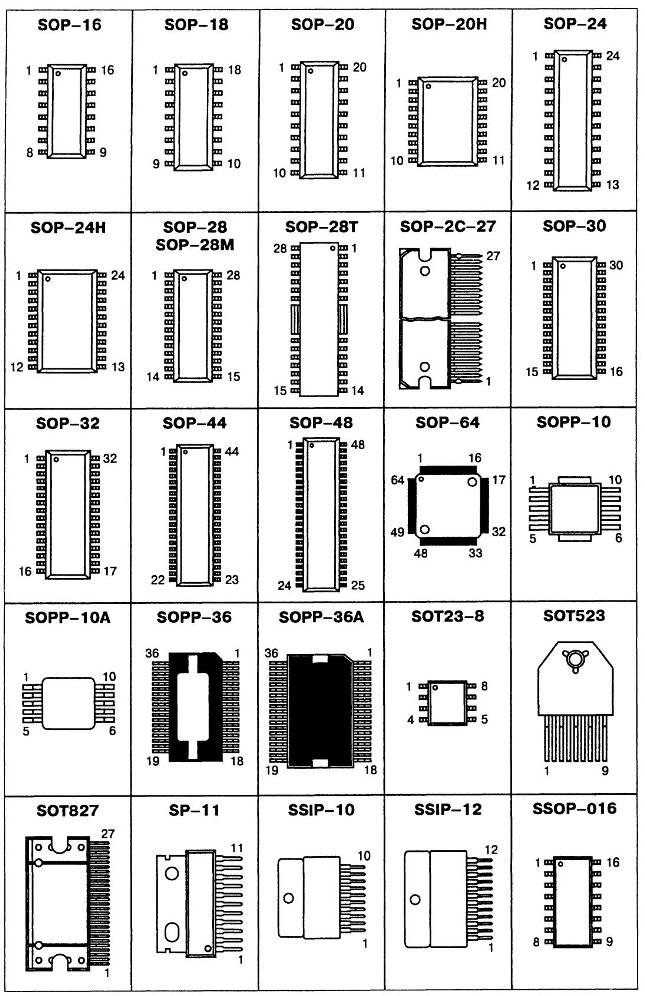
После этого откроется ещё одна страница и во внутреннем окне начнётся процесс загрузки PDF документа с информацией на электронный компонент.
После полной загрузки даташита его можно просмотреть. При необходимости его можно сохранить на компьютере, как и любой другой PDF файл. Сделать это можно, нажав на кнопку в виде дискеты, которая расположена на панели инструментов.
Появиться окно, где необходимо указать, где сохранить PDF файл и как он будет назван. Кроме такого способа сохранить даташит есть ещё один. Жмём правой кнопкой мыши на любом месте документа и в выпадающем меню выбираем “Сохранить как…”. Всё довольно знакомо.
Также можно распечатать даташит прямо из браузера. Для этого жмём кнопку с изображением принтера и указываем настройки печати.
В PDF документе приводится описание микросхемы PB137: структурная схема, стандартная схема включения, электрические характеристики, краткое описание назначения микросхемы, изображение корпуса прибора, таблицы с параметрами.
К сожалению все документы на иностранном языке (в основном на английском). При переводе интересующей информации можно пользоваться on-line переводчиками, например, переводчиком от Google.
Главная » Радиоэлектроника для начинающих » Текущая страница
Также Вам будет интересно узнать:
Технические справочники
Что-то не так?
Пожалуйста, отключите Adblock.
Портал QRZ.RU существует только за счет рекламы, поэтому мы были бы Вам благодарны если Вы внесете сайт в список исключений. Мы стараемся размещать только релевантную рекламу, которая будет интересна не только рекламодателям, но и нашим читателям. Отключив Adblock, вы поможете не только нам, но и себе. Спасибо.
Как добавить наш сайт в исключения AdBlock QRZ. RU > Технические справочникиclass=”small”>
RU > Технические справочникиclass=”small”>
Радиолюбительский разговорник EW1MM (WinWord)
Полупроводниковая техника
- Справочник по полупроводниковым диодам
- Справочник по полупроводниковым светоизлучающим приборам
- Справочник по полевым транзисторам
- Справочник по биполярным транзисторам
- Маркировка диодов
- Цветовая и кодовая маркировка транзисторов
- Краткие справочные данные зарубежных транзисторов
- Характеристики динисторов и тиристоров
- Справочник по отечественным и зарубежным полупроводниковым приборам: транзисторам, тиристорам, диодам – 8.4М
Микросхемы
- Номенклатура и аналоги отечественных микросхем
- Справочник по микросхемам ТТЛ серий
- Справочник по низкочастотным цифровым КМОП микросхемам
- Справочник по цифровым КМОП микросхемам
- Справочник по полупроводниковым аналоговым коммутаторам
- Справочник по операционным усилителям
- Справочник по стабилизаторам напряжения
- Справочник по микросхемам памяти
- Замена импортных ТТЛ микросхем отечественными аналогами
- Замена импортных КМОП микросхем отечественными аналогами
- Замена импортных операционных усилителей отечественными аналогами
- Замена импортных компараторов отечественными аналогами
- Справочник по электронной технике для студентов ВУЗов
- Справочник по аналоговым микросхемам для аудиоаппаратуры – 11.
 6М
6М - Справочник по микросхемам для теле- и видеоаппаратуры – 46.9М
- Справочник по цифровым логическим микросхемам ч1 – 26.8М
- Справочник по цифровым логическим микросхемам ч2 – 39.8М
- Справочник по полупроводниковым приборам – 24.1М
Светоизлучающие и фоточувствительные приборы
- Характеристики светоизлучающих диодов импортного производства
- Характеристики светоизлучающих диодов отечественного производства :: (Вариант в XLS)
- Характеристики светоизлучающих диодов отечественного производства –
Каталог по светодиодам, Нижний Новгород, PDF, 27 стр. - Характеристики инфракрасных светоизлучающих диодов АЛ164…172
- Фотодиоды КДФ115 – фотоэлектрические параметры
- Фототранзисторы
- Введение
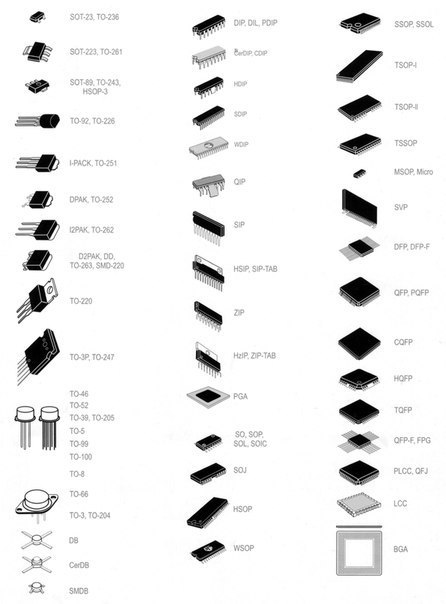
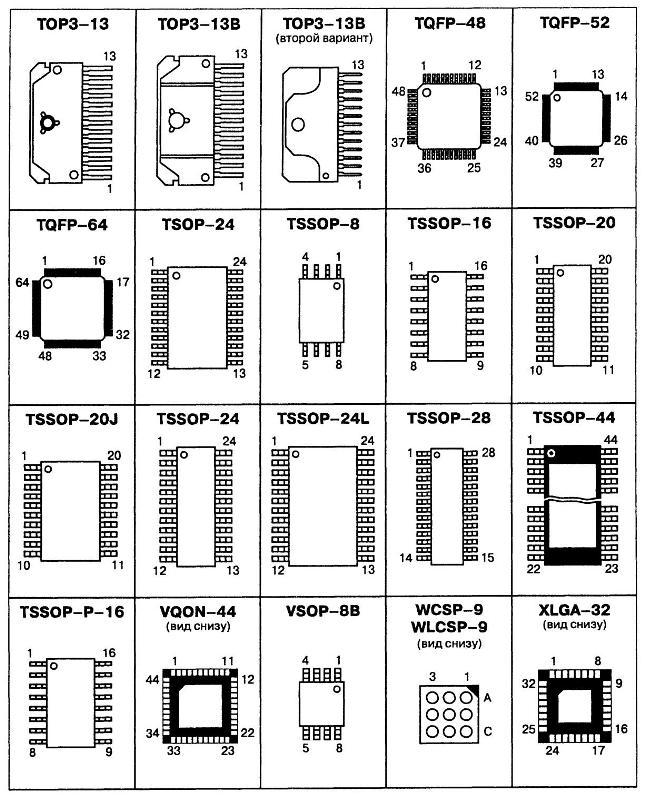
- Корпуса компонентов для поверхностного монтажа (SMD)
- Сквозная нумерация наиболее популярных корпусов SMD
- Ряды (резисторы и конденсаторы)
- Резисторы.
 Цветовая маркировка
Цветовая маркировка - Резисторы. Цветовая маркировка фирмы Philips
- Резисторы. Нестандартная цветовая маркировка
- Резисторы. кодовая маркировка
- Кодовая маркировка прецизионных высокостабильных резисторов фирмы Panasonic
- Перемычки и резисторы с “нулевым” сопротивлением
- Резисторы. Кодовая маркировка фирмы Philips
- Резисторы. Кодовая маркировка фирмы Bourns
- Конденсаторы. Допуски
- Конденсаторы. Температурный коэффициент емкости (ТКЕ)
- Конденсаторы. Цветовая маркировка
- Конденсаторы. Кодовая маркировка
- Конденсаторы. Кодовая маркировка электролитических конденсаторов для поверхностного монтажа (SMD)
- Индуктивности. Цветовая маркировка
- Индуктивности. Кодовая маркировка
- Транзисторы. Кодовая маркировка. Корпус КТ-26 (ТО-92)
- Транзисторы. Цветовая маркировка. Корпус КТ-26 (ТО-92)
- Транзисторы. Маркировка. Корпус КТ-27 (ТО-126)
Ламповая техника
- Термины и определения справочников по радиолампам
- Общие данные о лампах
- Рекомендации по применению
- Тренировка радиоламп
- Отечественные радиолампы
- Зарубежные радиолампы
- Приёмно-усилительные и генераторные лампы (список)
- Генераторные лампы
- Импульсные генераторные лампы
- Импульсные модульные лампы
- Ультрафиолетовые лампы серии ДРТ для стирания ПЗУ
- Импульсные лампы серии ИФК
- NEW! Приемные электронно-лучевые трубки.
 Справочник, Москва “Радио и связь”, Н.И. Вуколов, А.И. Гербин, Г.С. Котовщиков, 1993 г. – формат DjVU [что это такое], 289 стр (В этом справочнике представлены характеристики практически всех известных электронно-лучевых трубок)
Справочник, Москва “Радио и связь”, Н.И. Вуколов, А.И. Гербин, Г.С. Котовщиков, 1993 г. – формат DjVU [что это такое], 289 стр (В этом справочнике представлены характеристики практически всех известных электронно-лучевых трубок)
Осциллографические трубки
Аудиотехника
- Справочник по динамическим громкоговорителям
ВЧ техника
- Справочник по коаксиальным кабелям
- Справочник по высокочастотным трансформаторным устройствам (в одном файле 6 мб | разными файлами)
- Электрические характеристики радиочастотных коаксиальных кабелей со сплошной ПЭ изоляцией
- Пересчет dBm в вольты и ватты на нагрузке 50 ОМ
- Катушки индуктивности – физическая природа
- hot! Соединители радиочастотные коаксиальные (ВЧ-разъемы)
Антенны
- hot! Радиопрозрачные канатики Мастрант
- Компьютерное моделирование антенн на программе MMANA
Книга предоставлена автором на исключительных правах публикации только на сервере QRZ. RU ! Все копии файлов с любых других сайтов кроме сайта автора или QRZ.RU – краденые!
RU ! Все копии файлов с любых других сайтов кроме сайта автора или QRZ.RU – краденые!
Эта книжка посвящена описанию работы с одной из лучших на сегодняшний день и, что особенно следует подчеркнуть, бесплат.ной компьютерной программой моделирования антенн MMANA. Краткое описание программы было опубликовано в журнале «Радио» в 2001 г. (июнь.сентябрь). В этом издании приведено пол.ное ее описание. Особое внимание уделено тонкостям работы с MMANA, которые не освещенны в журнальном варианте, кроме того разобраны типичные ошибки, приведены ответы на часто встре.чающиеся вопросы. Наличие большой библиотеки файлов готовых антенн позволяет не только подобрать подходящую антенну, но и проверить на кон.кретных примерах уровень освоения программы. Также даны, хотя и не относящиеся непосредственно к MMANA, но желательные для уверенной работы и правильного понимания полученных результатов, основы компьютерного моделирования антенн.
- Характеристики основных типов военных антенн
Справочник по ферритам
- Ферритовые кольца в спортивной аппаратуре
- Ферромагнитные материалы
- Изделия завода “Ферроприбор”
Компьютерная техника
- Разводка контактов различных интерфейсов, разъемов, компьютерных шин и т.
 п.
п. - Компьютерные кабели – распиновка. Как сделать кабель последовательного интерфейса и много других
- Компьютерные адаптеры – схемы, описание, спецификации
- Различные схемы (активные фильтры и проч.)
- Справочные таблицы
Прочее
- Микрофоны электретные (формат XLS)
- Микрофоны электретные часть 2
- Определение номинала резистора по цветовой маркировке
- Схемы и параметры резисторных сборок М019НР1, М019НР2, М020НР1, М021НР1, М022НР1, М023НР1, М024НР1, М025НР1, М026НР1, М027НР1, М042НР1, М043НР1, М044НР1, М050НР1.
- Коммутационные переключатели 220В
- Изготовление сетевого предохранителя на любой ток
- Справочные данные трансформаторов ТА, ТН, ТАН, ТПП
- Батарейки и аккумуляторы
- Расчет трансформаторов – формулы, таблицы. Скан страниц из книги 50 х годов Гинзбурга. (DjVU)
- 10 лучших бесплатных онлайн симуляторов электроцепи
- hot! Телевизионные стандарты
- hot! Сокращения и условные обозначения
ГОСТ
- ГОСТ 23849-87 Аппаратура радиоэлектронная бытовая.
 Методы измерения электрических параметров усилителей сигналов звуковой частоты.
Методы измерения электрических параметров усилителей сигналов звуковой частоты. - ГОСТ 7845-92 – Системы вещательного телевидения. Основные параметры. Методы измерений.
| Благодарим! |
Информация для этого раздела предоставлена:
|
Рейтинг платформ поиска электронных компонентов / Хабр
Во всём мире несколько десятков миллионов наименований электронных компонентов.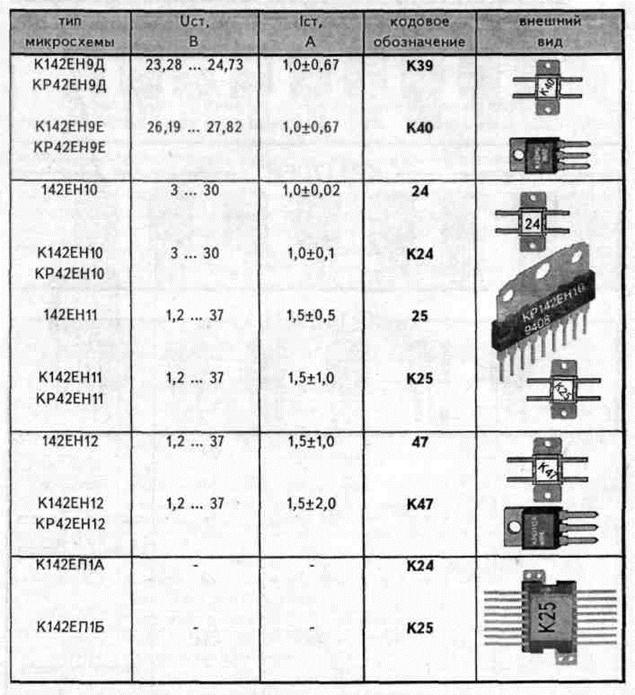 Для эффективной работы с таким количеством информации невозможно обойтись без специальных инструментов поиска. Поэтому, для того чтобы упростить связь между покупателем, продавцом и производителем, существуют так называемые «Поисковики электронных компонентов». В данный момент можно насчитать около двадцати подобных платформ как российских, так и иностранных.
Для эффективной работы с таким количеством информации невозможно обойтись без специальных инструментов поиска. Поэтому, для того чтобы упростить связь между покупателем, продавцом и производителем, существуют так называемые «Поисковики электронных компонентов». В данный момент можно насчитать около двадцати подобных платформ как российских, так и иностранных.В силу рода своей деятельности часто использую данные платформы для поиска и заказа компонентов. Сначала через параметрический поиск на digikey ищем нужный компонент, после через chipfind находим российского поставщика. Узнали себя? Если да, то эта статья для Вас.
Любители в основном пользуются платформами, которые хорошо разрекламированы, просто не зная о существовании возможности выбора. Специалисты же пользуются не одной, а целым набором. В этой статье постараюсь ответить на вопрос: почему не существует универсальной платформы для поиска электронных компонентов?
Все платформы электронных компонентов можно разделить по происхождению на российские и иностранные. К российским, т.е. ориентированными только на пользователей на территории Российской Федерации, относятся такие платформы как efind, chipfind, optochip итд. Иностранные: Digikey, mouser итд работают по всему миру.
К российским, т.е. ориентированными только на пользователей на территории Российской Федерации, относятся такие платформы как efind, chipfind, optochip итд. Иностранные: Digikey, mouser итд работают по всему миру.
Методика
Анализ электронных платформ проводился по 16 критериям, определяющим исключительно функциональные возможности платформы. Наличие реализованной функции приносит платформе 1 балл, а отсутствие 0 баллов. Всё просто. Если функция заявлена разработчиками платформы, но находится на стадии разработки, то платформа получает 0.5 балла. За рамками данного анализа остаются субъективные критерии: дизайн, удобство использования, стоимость доступа. По результатам анализа будет составлен рейтинг.
Список участвующих платформ
Скрытый текстВ список выше не попали следующие платформы: icmania.ru, rel.ru, chipportal.ru, zFind.ru по причине недоступности их сайта на момент написания статьи (январь 2018 года)
Результаты анализа «Российских платформ»
Результаты анализа «Иностранных платформ»
Сводная таблица, результаты анализа* Представленная информация актуальна на январь 2018 года
1 – присутствие русского языка на иностранных платформах, присутствие английского языка на русских платформах;
2 – частичный перевод;
3 – без учёта услуг рекламных услуг;
4 – поиск реализован всего для нескольких разделов;
5 – только раздел электронных компонентов;
$ – платный доступ
Итоговый рейтинг
Доступ в Brokerforum.
 com платный, после 15 дней пробного периода составит 400$ на полгода. О возможности доступа для физических лиц информация отсутствует
com платный, после 15 дней пробного периода составит 400$ на полгода. О возможности доступа для физических лиц информация отсутствуетВыводы
Рассмотренные платформы по функциональному признаку можно разделить на агрегаторы и поисковики. Агрегаторы получают информацию о наличии электронных компонентов от поставщиков или путём парсинга информации с других платформ, а том числе с digikey и mouser. Поисковики устроены значительно сложнее агрегаторов, кроме информации об остатке на складах предоставляют инструменты для поиска электронных компонентов по техническим характеристикам (параметрических поиск).
У российских платформ есть общий недостаток – отсутствие английской версии сайта, дополнительных тематических услуг, форумов, справочников, мобильного приложения. По иностранным платформам можно сказать, что у них есть только один недостаток – абсолютное отсутствие электронных компонентов отечественного производства.
Из всех рассмотренных платформ только одна (Optochip. org) обладает возможностью параметрического поиска отечественных компонентов. На трёх платформах (eFind.ru, ChipFind.ru, Optochip.org) есть возможность подбора аналога импорт/отечественная продукция, отечественная продукция/импорт. Также только три платформы обладают групповым поиском компонентов (Optochip.org, FindChips.com, Farnell.com). На своём сайте разместили тематическую литературу: Optochip.org, compel.ru, Digikey.com, Farnell.com. Форум для пользователей платформы присутствует у eFind.ru, Digikey.com, Farnell.com. Весьма интересным оказался результаты «Дополнительные тематические услуги» — все иностранные компании оказывают дополнительные услуги (аналитика, продвижение, доставка итд) из российских только одна Optochip.org. Также у большинства иностранных платформ присутствует мобильное приложение.
org) обладает возможностью параметрического поиска отечественных компонентов. На трёх платформах (eFind.ru, ChipFind.ru, Optochip.org) есть возможность подбора аналога импорт/отечественная продукция, отечественная продукция/импорт. Также только три платформы обладают групповым поиском компонентов (Optochip.org, FindChips.com, Farnell.com). На своём сайте разместили тематическую литературу: Optochip.org, compel.ru, Digikey.com, Farnell.com. Форум для пользователей платформы присутствует у eFind.ru, Digikey.com, Farnell.com. Весьма интересным оказался результаты «Дополнительные тематические услуги» — все иностранные компании оказывают дополнительные услуги (аналитика, продвижение, доставка итд) из российских только одна Optochip.org. Также у большинства иностранных платформ присутствует мобильное приложение.
На данный момент не существует ни одной российской платформы электронных компонентов, ориентированной на международный рынок. А все существующие иностранные платформы не ориентированы на российский. Именно поэтому не существует универсальное платформы для поиска электронных компонентов.
Именно поэтому не существует универсальное платформы для поиска электронных компонентов.
Не сочтите за рекламу, лично я пользуюсь следующим набором: Digikey, Brokerforum $, Optochip. Для тех, кто не хочет платить за доступ могу порекомендовать следующий набор: Digikey или Mouser, Alibaba, Optochip.
Маркировка точками микропроцессоров К1801ВМ1 – Soviet CPU Collection
На профильных форумах среди коллег неоднократно поднимался вопрос о маркировке 16-разрядных однокристальных микропроцессоров 1801ВМ1. Микросхемы маркировались точками. Производители этих процессоров – “Ангстрем” (Зеленоград), “Экситон” (г.Павловский Посад). Микросхемы с маркировкой буквой Г имели дополнительную 69 команду MUL-умножение.
Давайте расставим все точки над 1801
Маркировка 1801ВМ1
Однозначного ответа при обсуждении на форумах не получалось. Поиск на просторах интернета и в литературе – справочниках того периода – тоже больше запутывал, чем вносил ясность. Единственное место, где могла быть информация, это какие-либо официальные документы, например, ТУ. Но где взять ТУ? В свободном доступе такой информации не нашел. Во многих крупных городах во времена СССР существовали организации ЦНТИ – центры научно-технической информации. В некоторых городах они существуют до сих пор. Я решил посетить эту контору в нашем городе. Оказалось, что там масса разной интересной информации. Есть ТУ даже на совсем редкие процессоры, типа 1801ВЕ1 и 581ВЕ1. Что обнаружилось при прочтении ТУ – бК0.348.570-02ТУ-81?
Единственное место, где могла быть информация, это какие-либо официальные документы, например, ТУ. Но где взять ТУ? В свободном доступе такой информации не нашел. Во многих крупных городах во времена СССР существовали организации ЦНТИ – центры научно-технической информации. В некоторых городах они существуют до сих пор. Я решил посетить эту контору в нашем городе. Оказалось, что там масса разной интересной информации. Есть ТУ даже на совсем редкие процессоры, типа 1801ВЕ1 и 581ВЕ1. Что обнаружилось при прочтении ТУ – бК0.348.570-02ТУ-81?
ТУ было введено впервые 01.10.1981г. Срок действия ТУ с 1.12.81г. до 1.07.85г.
В первом ТУ речь шла только о микросхемах с буквами А и Б. Маркировались они следующим образом: А – одна точка, Б – две точки.
Максимальные частотные характеристики были следующие: К1801ВМ1А – 5 мГЦ, К1801ВМ1Б – 2,5 мГЦ.
Первое упоминание о микросхемах с буквой Г появилось в изменении №3 от 1.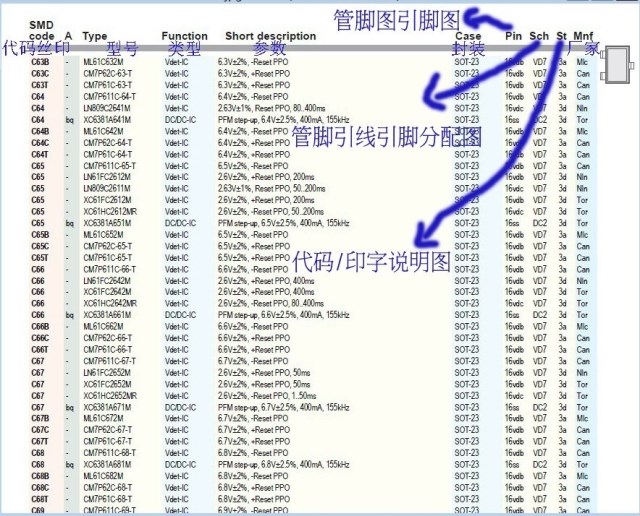 09.83г. Изменение было предписано внедрить с 1.01.84г.
09.83г. Изменение было предписано внедрить с 1.01.84г.
В этом изменении к ТУ предписывалось маркировать микросхемы следующим образом: А – одна точка, Б – две точки, В – три точки, Г – четыре точки.
В этом же изменении появилось упоминание о 69 команде MUL- умножении.
Частотные характеристики:
Фото микросхем того времени:
Последнее из приведенных фото – с платы ЧПУ Электроника НЦ-80. Предоставлено Сергеем Устюговым.
Следующий вариант в маркировке микросхем К1801ВМ1 появился в изменении №8 от 21.04.89г.
В данном изменении предписывалось маркировать А – без точек, Б – одной точкой, Г – двумя точками. В этом изменении было предписано следующее указание о внедрении: «Срок внедрения изменений в производстве устанавливается заводом-дублером». Скорее всего изменение было внедрено с начала следующего года. То есть с 1990г. В СССР новые дела было принято начинать либо с понедельника, либо с нового года :)
Скорее всего изменение было внедрено с начала следующего года. То есть с 1990г. В СССР новые дела было принято начинать либо с понедельника, либо с нового года :)
Частотные характеристики:
Фото имеющихся у меня микросхем этого периода:
Еще в ТУ встречаются такие строки: “Микросхемы К1801ВМ1Г поставляются только для НИИП и НИИНЦ и для изделия ”Электроника МС 2106”. Согласно ТУ выпускался также бескорпусной вариант микросхемы с маркировкой Б1801ВМ1Г.
Обсудить можно здесь
Defense Logistics Agency запускает возможность маркировки ДНК для усиления цепочки поставок микросхем
FORT BELVOIR, Вирджиния, 15 декабря 2014 г. / PRNewswire-USNewswire / – Агентство оборонной логистики прилагает усилия, чтобы упростить обнаружение и предотвращение попадания поддельных микросхем в свою цепочку поставок.
Сегодня агентство начало реализацию собственной инициативы по борьбе с подделкой микросхем, получившей название ДНК-маркировка. Эта возможность будет подтверждать подлинность приобретенных микросхем, одновременно повышая их надежность по всей цепочке поставок.Новые меры по контролю качества будут проводиться в Центре тестирования электронных продуктов DLA в DLA Land and Maritime, полевом агентстве, расположенном в Колумбусе, штат Огайо.
«Микросхемы интегрированы во многие системы вооружения, эксплуатируемые нашими военными службами, поэтому обеспечение надежных поставщиков имеет решающее значение для поддержания наших элитных готовых к выполнению задач сил», – сказал командующий сухопутными и морскими войсками DLA контр-адмирал Джон Кинг. «Внедрение этой возможности маркировки ДНК позволит DLA активизировать борьбу с попаданием контрафактных деталей и негативным влиянием на нашу цепочку поставок и, в конечном итоге, на наших клиентов.«
До внедрения этой возможности агентство полагалось на партнеров по отрасли для выполнения процесса маркировки ДНК. Это было эффективным в сдерживании фальшивомонетчиков, но добавляло больше времени на доставку микросхем покупателям и увеличивало расходы DLA.
Это было эффективным в сдерживании фальшивомонетчиков, но добавляло больше времени на доставку микросхем покупателям и увеличивало расходы DLA.
«Создавая возможности для органической или внутренней маркировки ДНК, DLA сократит время доставки и сократит затраты, усилит контроль цепочки поставок, повысит контроль качества и создаст возможность распространить маркировку ДНК на другие части, которые считаются высоким риском подделки», сказал директор центра тестирования продуктов DLA Кейт Робинетт.«Размещение ДНК-маркировки в Центре тестирования электронной продукции идеально подходит. Центр выполняет различные тесты, чтобы гарантировать, что электронные предметы, закупаемые DLA, соответствуют требованиям боевиков и требованиям к качеству. Маркировка ДНК расширит эту возможность».
Испытательный центр будет маркировать все микросхемы, приобретенные DLA, в классе федеральных поставок (FSC) 5962 с использованием технологии защиты от подделки, которая получена из дезоксирибонуклеиновой кислоты (ДНК) растений.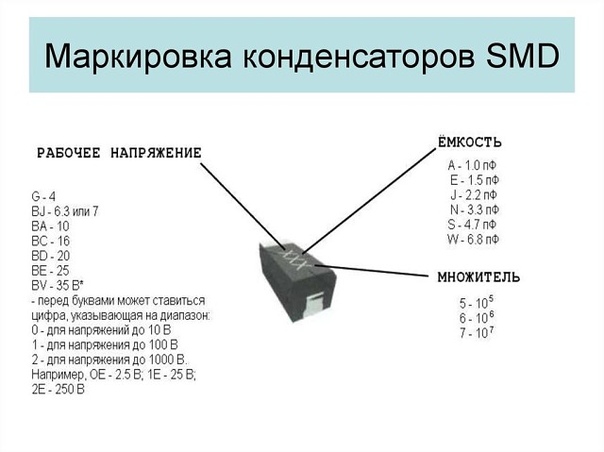 Предполагается, что в год будет маркироваться около 85 000 микросхем.
Предполагается, что в год будет маркироваться около 85 000 микросхем.
заключается в нанесении идентификатора ботанической ДНК на поверхность микросхемы для подтверждения подлинности. Уникальный код или отпечаток пальца, который отпугивает фальшивомонетчиков, встроен в чернила метки ДНК. Этот код не может быть воспроизведен, переработан или скопирован в цифровом виде. Метку ДНК можно обнаружить с помощью ручного сканера для легкой идентификации в цепочке поставок. Метку ДНК также можно удалить тампоном для судебно-медицинской экспертизы, которая предоставляет подробную информацию о микросхеме, такую как поставщик, код клетки, а также номер детали и партии.Также может быть получена дополнительная информация, такая как данные контракта, дата присуждения контракта, номер, национальный складской номер, количество и время включения микросхемы в цепочку поставок DLA.
Метка ДНК несет эту аутентификационную информацию на протяжении всего срока службы микросхемы и имеет возможность отслеживать и проверять движение микросхемы от момента поступления в цепочку поставок DLA до конечного пользователя.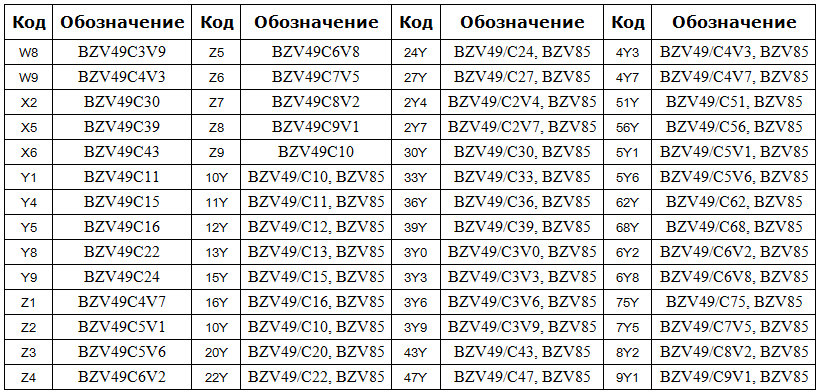 Это преимущество прослеживаемости имеет решающее значение при расследовании нарушений качества и мошенничества. До этого во время этих расследований часто было трудно получить информацию о поставщиках, когда микросхему вынимали из упаковки и устанавливали на систему оружия или помещали в бункеры на военно-ремонтных предприятиях.
Это преимущество прослеживаемости имеет решающее значение при расследовании нарушений качества и мошенничества. До этого во время этих расследований часто было трудно получить информацию о поставщиках, когда микросхему вынимали из упаковки и устанавливали на систему оружия или помещали в бункеры на военно-ремонтных предприятиях.
Как агентство боевой поддержки Министерства обороны, DLA предоставляет армии, военно-морскому флоту, военно-воздушным силам, корпусу морской пехоты, другим федеральным агентствам, а также объединенным и союзным силам различные логистические, закупочные и технические услуги. Агентство закупает и поставляет почти 100 процентов предметов потребления, необходимых для работы американских вооруженных сил, от продуктов питания, топлива и энергии до униформы, медицинских принадлежностей, строительного и заградительного оборудования. DLA также поставляет более 85 процентов запасных частей для военных.
Со штаб-квартирой в Форт-Бельвуар, штат Вирджиния, DLA насчитывает около 25 000 сотрудников по всему миру и обслуживает более 2440 систем вооружения. Для получения дополнительной информации о DLA посетите www.dla.mil, www.facebook.com/dla.mil или http://twitter.com/dlamil.
Для получения дополнительной информации о DLA посетите www.dla.mil, www.facebook.com/dla.mil или http://twitter.com/dlamil.
– http://photos.prnewswire.com/prnh/20130918/MM82677LOGO
Чтобы просмотреть исходную версию на PR Newswire, посетите: http://www.prnewswire.com/news-releases/defense-logistics-agency-launches-dna-marking-capability-to-strengthen-microcircuit-supply-chain- 300009908.html
ИСТОЧНИК Агентство оборонной логистики
Ссылки по теме
http://www.dla.mil
US DLA рассматривает возможность расширения планов маркировки ДНК
Агентство оборонной логистики США (DLA) рассматривает возможность расширения использования маркировки ДНК для защиты запасных частей и компонентов от подделки.
Использование ДНК-маркеров для предотвращения попадания поддельных микросхем, предназначенных для использования военными США в цепочку поставок, было впервые санкционировано ближе к концу прошлого года, когда DLA потребовало, чтобы технология применялась ко всем классам федеральных поставок (FSC) 5962 электронные микросхемы.
Мандат оказался неоднозначным, поскольку существует только один поставщик, способный удовлетворить требования – Applied DNA Sciences (APDN), который уже подписал более двух десятков контрактов на поставку своих маркеров Signature DNA поставщикам микросхем.
В то время, когда было объявлено требование, Ассоциация полупроводниковой промышленности США (SIA) заявила, что эта мера не решит проблему подделок, заявив, что ее «легко обойти, потому что фальшивомонетчику нужно только имитировать материал маркера».В нем также говорится, что это обременит производителей без снижения риска и фактически создает монополию на поставку маркеров.
Несмотря на критику DLA, теперь заявило, что оно рассматривает возможность расширения диапазона компонентов, требующих маркировки защиты от подделки, чтобы включить в него следующие элементы, согласно недавней презентации, сделанной на информационном форуме электронной и электротехнической промышленности в 2013 году:
- FSC 3110, Подшипники (авиация)
- FSC 4730, Фитинги, шланги и трубки (наземные и морские или L&M)
- FSC 5325, крепежные детали (поддержка войск)
- FSC 5935, Электрические соединители (L&M)
- FSC 5961, Полупроводниковые приборы (L&M)
Расширение будет означать, что эти категории продуктов будут закупаться правительством США только в том случае, если поставщик соглашается применять маркеры, как это уже имеет место в случае микросхем FSC 5692.

Агентство управляет 548 различными классами компонентов и деталей, охватывающих 5,1 миллиона отдельных предметов, и хотя электронные компоненты считаются подверженными наибольшему риску в случае подделки, оно также обеспокоено аксессуарами двигателя, трубами и фитингами, оборудованием, абразивными материалами и компонентами автомобилей.
«Контрафакция представляет собой растущий риск для готовности к миссии, безопасности персонала и национальной безопасности», – заявило DLA на мероприятии, отметив, что глобализация создала уязвимости в цепочке поставок.
Маркировка ДНКявляется предпочтительным выбором для защиты оригинальных деталей, поскольку она относительно недорогая, не может быть скопирована, обеспечивает аутентификацию на уровне судебной экспертизы, не влияет на функциональность товара и может использоваться для проверки товаров в нескольких точках цепи поставок. DLA уже заявило, что возместит поставщикам стоимость материала ДНК-маркера.
APDN явно в восторге от этой новости, и генеральный директор Джеймс Хейворд сказал, что «коммерческий опыт компании в маркировке всего, от медных кабелей до пластиковых оплеток и тканей, можно передать этим FSC.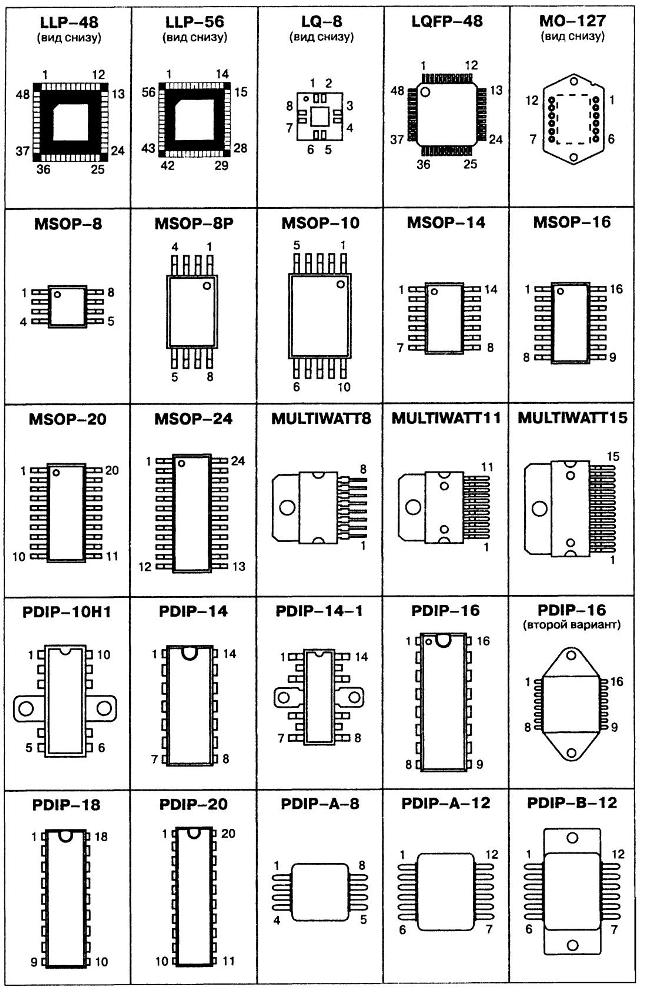 «
«
Цепочка поставок электроники продолжает борьбу с контрафактными деталями
Прошел почти год с тех пор, как президент Обама подписал Закон о национальной обороне (NDAA), и его конечное влияние на компании в цепочке поставок электроники все еще остается горячей темой. Этой осенью дистрибьюторы, в частности, были заняты подготовкой к шквалу действий, связанных с разделом 818 NDAA, который касается новых требований правительства к закупке электронных компонентов для использования в оборонных системах и оборудовании – правила, которые включают расширенные процедуры проверки и отчетности и более тщательный контроль со стороны Министерство внутренней безопасности над странами, считающимися значительными источниками контрафактных компонентов.По состоянию на середину сентября одно из новейших требований, связанных с разделом 818, было от правительственного агентства по логистике обороны, требуя маркировки ДНК на электронных микросхемах.
В конце лета, Electronic Design / SourceESB встретился с некоторыми ключевыми сторонами, вовлеченными в проблему – независимыми дистрибьюторами, уполномоченными дистрибьюторами, представителями правительства и поставщиками дополнительных услуг – чтобы узнать, что они делают, чтобы подготовиться к новым правилам и чтобы узнать их мнение о состоянии проблемы контрафакта в конце 2012 года.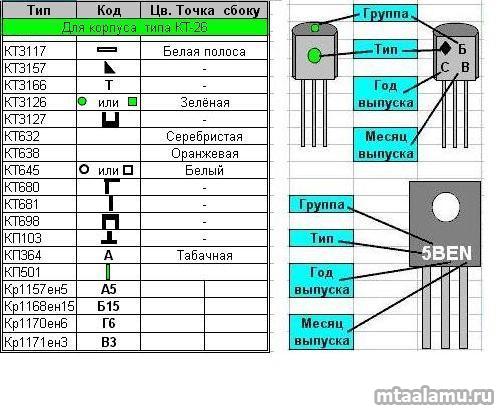 Мы поговорили с Дженис Мераглиа, вице-президентом по военным и правительственным программам прикладных наук о ДНК, которая создает решения для обеспечения безопасности и аутентификации на основе ботанической ДНК для широкого круга отраслей; Стив Мартин, исполнительный вице-президент по продажам Components Direct, авторизованного дистрибьютора электронных компонентов с излишками и отслужившими свой срок; Кристин Мец, ответственный за технические и качественные процессы в Управлении оборонной логистики федерального правительства; Мэтт Харцелл, главный операционный директор независимого дистрибьютора NF Smith & Associates; и Кевин Синк, вице-президент по общему качеству официального дистрибьютора TTI Inc.Хартцелл и Синк также являются членами комитета G19 SAE International, который занимается вопросами предотвращения, обнаружения и реагирования электронной промышленности на угрозу контрафакта.
Мы поговорили с Дженис Мераглиа, вице-президентом по военным и правительственным программам прикладных наук о ДНК, которая создает решения для обеспечения безопасности и аутентификации на основе ботанической ДНК для широкого круга отраслей; Стив Мартин, исполнительный вице-президент по продажам Components Direct, авторизованного дистрибьютора электронных компонентов с излишками и отслужившими свой срок; Кристин Мец, ответственный за технические и качественные процессы в Управлении оборонной логистики федерального правительства; Мэтт Харцелл, главный операционный директор независимого дистрибьютора NF Smith & Associates; и Кевин Синк, вице-президент по общему качеству официального дистрибьютора TTI Inc.Хартцелл и Синк также являются членами комитета G19 SAE International, который занимается вопросами предотвращения, обнаружения и реагирования электронной промышленности на угрозу контрафакта.
Следующие комментарии являются выдержками из наших разговоров.
Electronic Design / SourceESB: Прошлогоднее принятие NDAA с его разделом 818, в котором подробно описаны действия по ограничению потока поддельных электронных компонентов в цепочку поставок оборонной продукции, привлекло новое внимание к проблеме контрафактной продукции. Как повышенное внимание помогает в борьбе с контрафактными электронными компонентами?
Как повышенное внимание помогает в борьбе с контрафактными электронными компонентами?
Мэтт Хартцелл: Учитывая интенсивную направленность слушаний NDAA и Комитета по вооруженным силам Сената, которые предшествовали этому, было написано и опубликовано огромное количество информации о проблеме подделки на электронном рынке. А за последние несколько лет количество сообщений [о случаях поддельных деталей] резко возросло.
Во всяком случае, я думаю, что проблема настолько сосредоточена, что сейчас она вроде как в статус-кво.К сожалению, одной контрафактной детали слишком много, поэтому мы должны проявлять бдительность в наших усилиях, чтобы не допустить попадания контрафактных деталей в цепочку поставок.
Steve Martin: Проблема столь же серьезна, как и прежде, и может быть еще серьезнее прямо сейчас, если вы посмотрите на цепочку поставок в целом. Есть много людей, которые пытаются удовлетворить потребности клиентов, но у них нет средств обнаруживать плохие детали.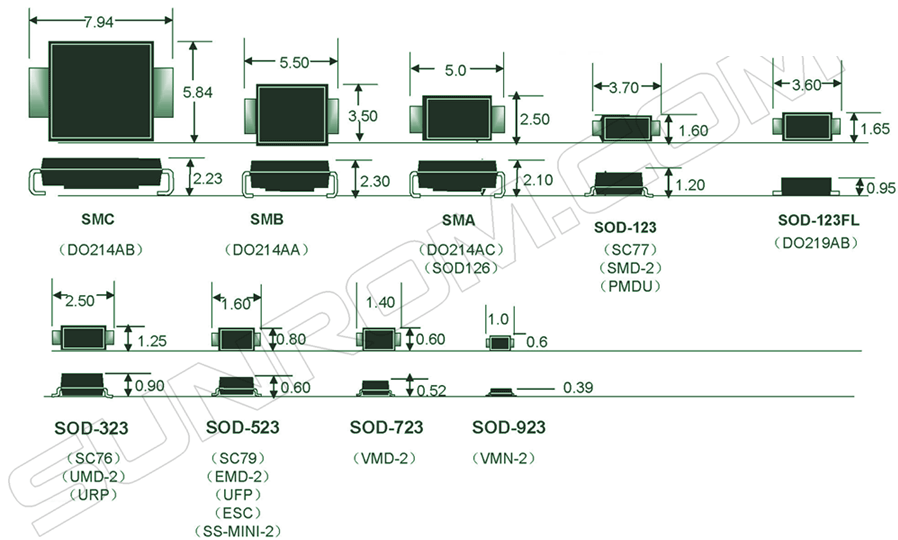 [Клиенты] должны убедиться, что они сотрудничают с правильными компаниями.
[Клиенты] должны убедиться, что они сотрудничают с правильными компаниями.
Кевин Синк: Я думаю, что сейчас он достиг критической массы. Раздел 818 NDAA и майский отчет сенатского комитета по вооруженным силам – вместе эти две вещи как бы вынудили [компании к действиям]. Клиенты требуют, чтобы поставщики заявили, что у них действует программа борьбы с контрафактной продукцией. Я, наверное, получаю три таких [запроса] в неделю, тогда как три года назад я ни одного не получал.
Electronic Design / SourceESB: Но, как авторизованному дистрибьютору, TTI действительно не нужно беспокоиться о плане борьбы с подделкой, верно? Какова ваша роль в движении по борьбе с контрафактом и, в частности, в комитете G19 САЕ?
Приемник: Почти все клиенты говорят [всем]: «Мы хотим убедиться, что вы знаете о проблеме и у вас есть план.'[Они также] хотят знать, покупали ли вы когда-нибудь вне авторизованного канала и как вы с этим справляетесь. Некоторые требуют, чтобы вы вообще не покупали за пределами авторизованного канала – что для меня нормально, потому что мы этого не делаем.
Некоторые требуют, чтобы вы вообще не покупали за пределами авторизованного канала – что для меня нормально, потому что мы этого не делаем.
В комитете [G19] есть три уполномоченных представителя по распространению…. Мы представляем авторизованное распространение и, в этом случае, гарантируем, что написанные правила справедливо защищают клиентов, когда они совершают покупки вне разрешенного распределения. Мы, конечно, не поддерживаем это, но мы хотим, чтобы правила были справедливыми.
Откровенно говоря, до недавнего времени многие клиенты не понимали разницы между авторизованным и независимым дистрибьютором … Например, недавно у меня был клиент, который хотел потребовать от каждого поставщика проведения интенсивного визуального осмотра продукта – и он не делал различий в том, что покупалось у независимых дистрибьюторов, официальных дистрибьюторов или производителей. Какая трата денег, когда производитель, создавший продукт, и официальный дистрибьютор, который продает продукт, доказывают, что это такое.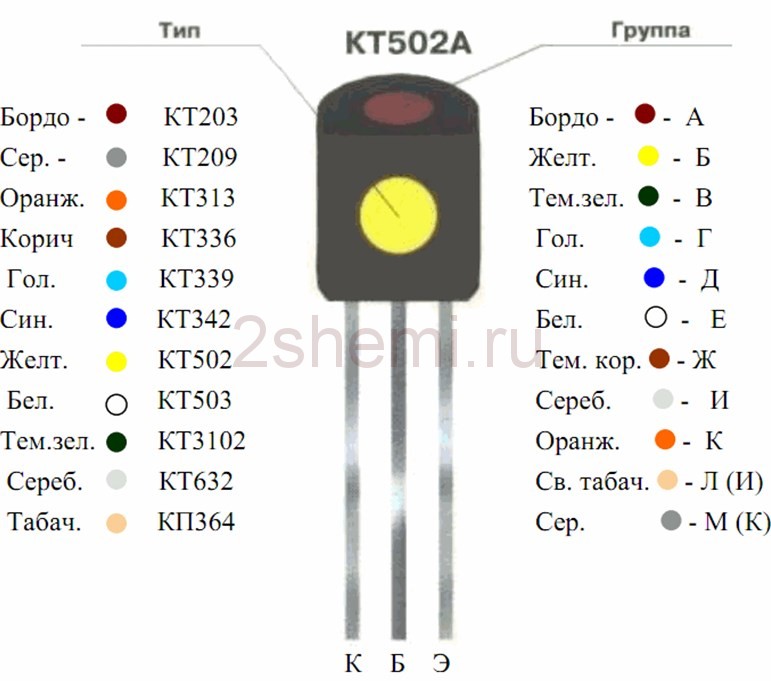
Electronic Design / SourceESB: Обсуждение контрафактной электроники часто по многим причинам сосредоточено исключительно на военном и аэрокосмическом рынках. Насколько серьезна проблема для других отраслей?
Hartzell: Это то, что кипело до того, как кипящий поток на слушаниях в Сенате. [Многие люди] начали верить, что в прошлом году это была большая проблема для военных и правительства. Но если вы занимаетесь этим бизнесом в течение длительного времени, вы знаете, что есть люди, которые долгое время пытались быстро заработать и продавать товары ниже приемлемого качества.Не менее важно, чтобы коммерческие клиенты знали, что их продавец пользуется репутацией, применяет процессы [устранения] подделок и может поддерживать продаваемые ими продукты. Их риск, который не опасен для жизни, если вы не говорите о коммерческой авиации или аналогичных отраслях, заключается в том, что продукт не будет работать, загорится или причинит физический ущерб. Меньше всего [коммерческие клиенты] хотят столкнуться с оглаской провального продукта или, что еще хуже, отзывом.
Меньше всего [коммерческие клиенты] хотят столкнуться с оглаской провального продукта или, что еще хуже, отзывом.
Martin: Причина, по которой [проблема] была определена Министерством обороны, состоит в том, что у них большое влияние.Но неважно, потребительское это приложение, медицинское, оборонное или автомобильное, у всех одинаковый уровень беспокойства.
Электронный дизайн / ИсточникESB: Что будет дальше на горизонте борьбы с контрафактом?
Приемник: В следующем году будет большая реакция на правила Раздела 818. Крайний срок [был] сентябрь, поэтому вскоре после этого я ожидаю увидеть еще один шквал опросов, о которых я упоминал ранее [относительно планов поставщиков по борьбе с контрафактом].Я также наблюдаю активную деятельность, связанную с изменением процедур компаний, ограничением их базы поставок и усилением аутентификации продукта на входящей точке получения – и, если есть какие-либо вопросы, гораздо больше продуктов отправляется в испытательные лаборатории. Этого будет немного больше.
Этого будет немного больше.
Мартин: Я думаю, что правительство США должно ввести более строгие правила и нормы и работать рука об руку с другими странами [для борьбы с проблемой]. Пока этого не произойдет, фальшивомонетчики будут стараться быть на шаг впереди процесса.
Hartzell: В последние несколько лет на отраслевых совещаниях все чаще обсуждались [проблемы] несоответствующей продукции на рынке и необходимость повышения бдительности, способности независимых инспекторов, страховки и т.д. и их ресурсы, которые сосредоточены исключительно на том, чтобы убедиться, что они продают хорошие продукты. [В результате] небольшим компаниям в этой отрасли становится все труднее вести бизнес. И я думаю, вы увидите, что это продолжится.
Правительство принимает меры
Одним из новейших вопросов, вытекающих из требований NDAA, является постановление правительства о том, чтобы все микросхемы, поставляемые и управляемые через Агентство логистики обороны, были помечены технологией ДНК.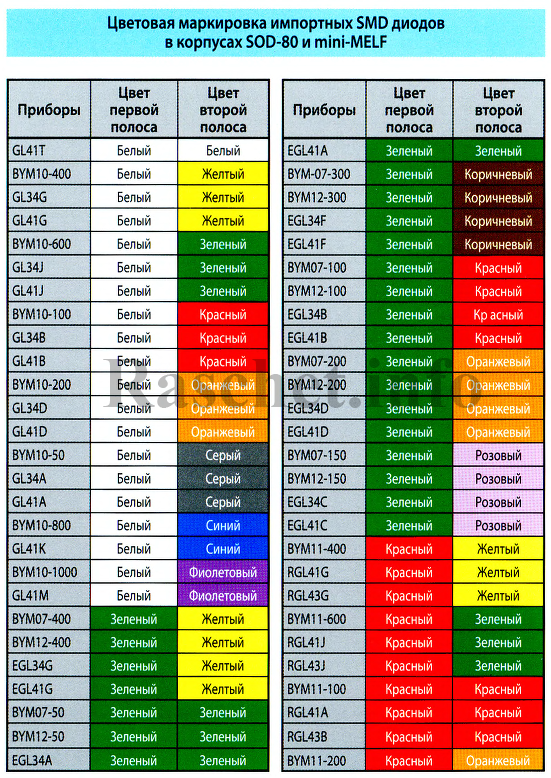 DLA – это агентство боевой логистики, которое поставляет широкий спектр продукции для вооруженных сил США по всему миру. Мы попросили Кристин Мец из DLA и Дженис Мераглию из его авторизованного провайдера маркировки ДНК Applied DNA Sciences объяснить требования и их значение для цепочки поставок.
DLA – это агентство боевой логистики, которое поставляет широкий спектр продукции для вооруженных сил США по всему миру. Мы попросили Кристин Мец из DLA и Дженис Мераглию из его авторизованного провайдера маркировки ДНК Applied DNA Sciences объяснить требования и их значение для цепочки поставок.
ED / SourceESB: Каковы новые требования к маркировке ДНК при продаже в Агентство логистики обороны?
Metz: Новое требование – это маркировка дезоксирибонуклеиновой кислоты (ДНК) для изделий, управляемых DLA, в рамках федерального класса поставок 5962, электронные микросхемы. DLA считает микросхемы очень опасными для подделки. Первоначальное требование относится к подмножеству микросхем, поставляемых в рамках программы Generalized Emulation of Microcircuits (GEM).SRI International является подрядчиком DLA по этим конкретным микросхемам. Однако в течение следующих 90 дней DLA намеревается применить это требование к другим объектам, управляемым DLA в рамках FSC 5962.
В пятницу, 3 августа 2012 г., DLA разместило уведомление на сайтах DLA Internet Bid Board System (DIBBS) и Центра информационных ресурсов для поставщиков (SIRC) о введении нового требования к маркировке. DLA отдельно запросила отзывы о новом требовании от поставщиков DLA FSC 5962.
ED / SourceESB: Как это влияет на поставщиков? Есть ли период поэтапного внедрения?
Metz: Поставщики, которые предоставляют товары DLA FSC 5962, должны будут предоставить товары, отмеченные уникальным ботаническим знаком ДНК SigNature (r), предоставленные Applied DNA Sciences или ее уполномоченными лицензиатами, если таковые имеются.Используемая аутентификационная ДНК должна быть уникальной для поставщика или производителя детали. Маркировка ДНК может быть нанесена с помощью видимой или невидимой метки ДНК на детали, или чернила подрядчика, используемые для маркировки деталей, могут быть смешаны с материалом для маркировки ДНК. Кроме того, поставщики должны будут хранить документацию по отслеживаемости, которая демонстрирует, что предметы, предоставленные по контракту DLA, имеют маркировку ДНК SigNature (r), произведенную Applied DNA Sciences или уполномоченным лицензиатом, и что маркировка ДНК является уникальной для подрядчика. .
.
Есть поэтапный подход. Сначала требования будут применяться к эмулируемым микросхемам (август 2012 г.), а затем ко всем FSC 5962 (ноябрь 2012 г.).
DLA выполняет это требование к маркировке для FSC 5962, чтобы внедрить эффективную маркировку аутентификации, одновременно инициируя исследования и разработки для оценки сопоставимых технологий ДНК и альтернативных аутентификационных технологий для предполагаемого применения ко всей электронике. Предстоящий запрос информации (RFI) запросит вводные данные относительно технологий аутентификации маркировки, которые удовлетворяли бы требованиям DLA, изложенным в RFI.
ED / SourceESB: Какие конечные продукты / оборудование поможет защитить это новое требование?
Metz: Это новое требование поможет защитить большинство систем вооружения Министерства обороны США. Многие предметы, которыми управляет DLA, используются более чем в одной системе оружия. DLA изначально ориентирована на микроэлектронику.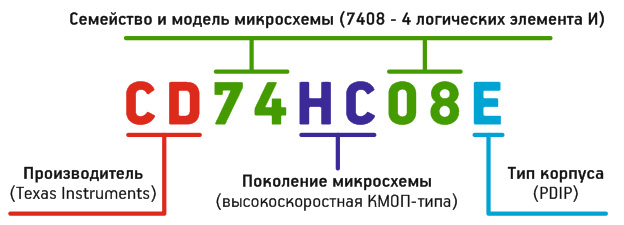 Технология используется с другими товарами и имеет широкое значение для других продуктов и оборудования DLA, не содержащих микросхемы.
Технология используется с другими товарами и имеет широкое значение для других продуктов и оборудования DLA, не содержащих микросхемы.
ED / SourceESB: Как вы думаете, насколько это повлияет на борьбу с контрафактной электроникой?
Metz: Выполнение этого нового требования снизит риск попадания контрафактных деталей в цепочки поставок Министерства обороны. Как правило, DLA намеревается внедрить детали с лучшими атрибутами аутентификации, и новое требование к маркировке является одним из инструментов для достижения этой цели и частью комплексной программы DLA для обнаружения и предотвращения подделок.Новое требование способствует достижению целей недавних законов (NDAA), требующих от Министерства обороны и промышленности принятия мер по обнаружению и предотвращению контрафактных товаров для электронных товаров и комплексной национальной кибербезопасности.
Стратегия DLA включает постоянное применение новых или обновленных инструментов для обнаружения и предотвращения контрафактной продукции по мере непрерывного развития тактики фальсификаторов. DLA на данный момент требует аутентификационной маркировки микросхем с использованием ДНК SigNature (r) на основе успешного проекта НИОКР.Мы открыты для других решений и инструментов, когда узнаем о них.
DLA на данный момент требует аутентификационной маркировки микросхем с использованием ДНК SigNature (r) на основе успешного проекта НИОКР.Мы открыты для других решений и инструментов, когда узнаем о них.
Усилия DLA по популяризации концепции аутентификационной маркировки и поощрению более широкого признания положительно влияют на практику управления рисками и разработку решений участников цепочки поставок Министерства обороны.
DLA разработало запрос информации (RFI) для маркировки аутентификации, который будет открыт для дополнительных технологических решений. DLA планирует выпустить RFI в октябре 2012 года.
ED / SourceESB: Какова роль прикладных наук о ДНК в этом процессе?
Meraglia: Последние пару лет мы разговаривали с представителями отрасли, поэтому люди достигли той точки, когда они понимают технологию [нашей ДНК], и теперь они говорят: «Что это значит для меня? Что мне делать? С чего начать? »Более конкретно, мы потратили последние несколько недель [с момента объявления DLA], отвечая на запросы, помогая людям быстрее понять, что влечет за собой процесс.
ED / SourceESB: Вкратце, как работает технология и что она даст для отрасли?
Meraglia: Когда вы смотрите на микрочип, это обычно металлический или керамический корпус, затем у вас есть матрица и так далее, и у вас есть определенная информация о микросхеме – на ней может быть дата, код партии и производитель имя, напечатанное на нем, например. Если вы печатаете эту информацию на чипе, мы можем нанести ДНК на чернила, которые вы используете.С другой стороны, есть компании, которые могут выгравировать информацию на чипе лазером. [Если это так], тогда есть другие способы применить ДНК.
В любом случае мы считаем, что это чрезвычайно мощный инструмент для решения задач, которые необходимо выполнить в этой отрасли. Мы стараемся работать с производителями, потому что это дает наибольшую выгоду в цепочке поставок. Это обеспечивает максимальную выгоду для всех участников, [потому что] он может быть аутентифицирован в различных точках. Чем дальше отмечена [часть] вверх по течению, тем лучше для всех.
Чем дальше отмечена [часть] вверх по течению, тем лучше для всех.
ED / SourceESB: Одна из самых больших проблем в войне с контрафактными компонентами – это эволюция подделок – фальшивомонетчики продолжают находить способы обойтись методами обнаружения. Насколько эффективна технология ДНК?
Meraglia: У нас были довольно сильные организации, которые пытались скопировать то, что мы делаем, но никто не мог этого сделать. Мы используем ДНК растений, и она настолько же сильна или сильнее, чем ДНК человека.Фальсификаторы не смогут прорваться.
На этапе исследований и разработок с DLA они искали независимую проверку того, что мы делаем. Это важная часть процесса. По мере продвижения вперед мы уходим от демонстрации [технологии] сами в пользу того, чтобы другие продемонстрировали, насколько хорошо она работает.
В конце концов, мы хотим, чтобы плохие парни не появлялись в этой индустрии. Мы хотим, чтобы им было настолько сложно попытаться [подделать], что они просто уходят из бизнеса.
Стандартизация деталей – Lansdale Semiconductor
Стандартизация деталей уходит в прошлое?Переход с MIL-M38510 (QPL) на MIL-PRF-38535 (QML) создал условия, при которых компоненты, стандартизированные для военных целей, не соответствуют «Передовой коммерческой практике» и могут даже нарушать текущие методы управления конфигурацией. Этот документ проведет вас через исторические изменения, которые привели к ослаблению требований в рамках MIL-PRF-38535, и покажет, как DLA адаптировало этот документ для создания этого условия вопреки советам как пользователей, так и производителей продуктов.
A История технических характеристик продукта:
Первоначально существовали SCD (чертежи системы контроля версий): эти документы определяли экологические, электрические, маркировочные, упаковочные и технологические требования для деталей. Проблема с SCD заключалась в том, что несколько NSN создавались по существу для одного и того же номера детали.
Тогда родились части Военного стандарта. MIL-M-38510 (Общие технические условия для микросхем) определял систему качества, обработку пластин, сборку и упаковку, квалификацию, соответствие требованиям к контролю качества для всех частей, предназначенных для «косой черты» и «SMD».В нем также определены группы микросхем, основанные на технологии устройства. Основным преимуществом было то, что для стандартизованных деталей на каждую деталь приходился только один NSN. Детали были внесены в Список квалифицированных деталей (QPL).
M38510 / XXX и 5962-XXXXX стали стандартными номерами деталей. Спецификации отдельных устройств определяли требования к электрическим испытаниям, так что несколько производителей могли квалифицировать один и тот же продукт по одной спецификации (например, TI, National и Motorola 5400 TTL, хотя и произведенные несколько иначе, будут квалифицироваться под одним и тем же номером детали).Номера групп микросхем для всех частей на слэш-листе определены в разделе 3 «Требования».
Система изменилась, когда была создана MIL-PRF-38535 (Общие технические условия для производства интегральных схем) QML (Квалифицированная производственная линия). QML – это спецификация производительности, означающая, что она носит описательный, а не предписывающий характер, при этом в большинстве приложений приводятся руководящие принципы, а не истинные требования. QML дает производителю устройства гибкость, позволяющую исключить тестирование, не добавляющее ценности, на основе статистических данных.Спецификация предназначена для документирования «Лучшей коммерческой практики». Многие определения в MIL-M-38510 были изменены или исключены, многие требования были удалены, и были добавлены новые определения. Требования к электрическим испытаниям по-прежнему покрываются спецификациями M38510 / XXX и 5962 -XXXXX.
Как указано в основной части MIL-PRF38535, многие определения, ранее содержащиеся в MIL-M-38510, были либо изменены, либо удалены. Среди них следует отметить то, что произошло с определениями типа устройства и группы микросхем.![]() Предыдущее определение типа устройства, которое требовало, чтобы части были электрически и функционально взаимозаменяемыми на уровне кристалла или подложки, было опущено в новом определении. Определение группы микросхем, которое требовало, чтобы изделие было одной базовой схемотехники, было полностью удалено. Оба эти определения в Приложении A к MIL-PRF-38535 очень похожи на старые требования MIL-M-38510, однако Приложение A к MIL-PRF-38535 не требуется для производителей QML (хотя они по-прежнему необходимы для класс M, поставщики MIL-STD-883).Таблица A-VI, в которой нумеруются группы микросхем, также все еще находится в Приложении A и определяет, что стандартный TTL-вентиль находится в группе микросхем 1; в нем также указано, что вентиль BiCMOS находится в группе 125.
Предыдущее определение типа устройства, которое требовало, чтобы части были электрически и функционально взаимозаменяемыми на уровне кристалла или подложки, было опущено в новом определении. Определение группы микросхем, которое требовало, чтобы изделие было одной базовой схемотехники, было полностью удалено. Оба эти определения в Приложении A к MIL-PRF-38535 очень похожи на старые требования MIL-M-38510, однако Приложение A к MIL-PRF-38535 не требуется для производителей QML (хотя они по-прежнему необходимы для класс M, поставщики MIL-STD-883).Таблица A-VI, в которой нумеруются группы микросхем, также все еще находится в Приложении A и определяет, что стандартный TTL-вентиль находится в группе микросхем 1; в нем также указано, что вентиль BiCMOS находится в группе 125.
Итак, что эти изменения значат для пользователей и производителей? В настоящее время часть, которая производится в соответствии со стандартом MIL-STD-883, параграф 1.2.1, должна соответствовать исходным требованиям MIL-M-38510, как теперь определено в Приложении A стандарта MIL-PRF-38535. Производитель MIL-PRF-38535 QML не обязан соблюдать требования Приложения A.
Производитель MIL-PRF-38535 QML не обязан соблюдать требования Приложения A.
Так это проблема? Один производитель QML получил от DLA / DSCC специальное разрешение на поставку детали из BiCMOS размером 1,5 микрона с тем же номером детали, что и деталь с биполярным TTL-сигналом с 7 микронами, легированными золотом, даже несмотря на то, что на слэш-листе эта деталь указана как принадлежащая к группе микросхем 1, а не 125, и электрические испытания показывают, что детали реагируют иначе, чем оригинал, в военном температурном диапазоне. (Посетите веб-сайт Lansdale http://www.lansdale.com/gemhtml/wkparts.htm, чтобы просмотреть фактические данные испытаний для 5 различных номеров деталей.) Никакие другие производители не могут это делать, поскольку все изменения, которые они вносят в технологию устройства, требуют изменения номера детали.
Другие новые определения в стандарте MIL-PRF-38535 включают 6.4.24 Форма, 6.4.25 Подгонка и 6.4.26 Функция. Форма, как определено в MIL-PRF-38535, определяет только физические и визуальные параметры и не упоминает технологию устройства. Подгонка относится только к возможности физического взаимодействия, функция относится к действию или действиям, для выполнения которых предназначен элемент, которые были интерпретированы как строгие требования спецификации без каких-либо «недокументированных» параметров.
Подгонка относится только к возможности физического взаимодействия, функция относится к действию или действиям, для выполнения которых предназначен элемент, которые были интерпретированы как строгие требования спецификации без каких-либо «недокументированных» параметров.
Действительно ли MIL-PRF-38535 является наилучшей коммерческой практикой? Документ с требованиями EIA Engineering Aerospace Qualified Electronic Component (AQEC), который станет лучшей коммерческой практикой для компонентов авионики, так не считает. В нем конкретно указано, что части устройств с разными технологиями не должны поставляться под одним и тем же номером детали, поскольку они не имеют одинаковой формы, соответствия и функций. Это означает, что устройства QML, которые могут поставляться через программу GEM как устройства, отличные от Gem, не допускаются как устройства AQEC.
В мае 2003 года в DLA было направлено совместное письмо комитетов JC13.2 и G-12. В этом письме говорилось, что, хотя программа GEM, поддерживающая устаревшие интегральные схемы для материально-технического обеспечения систем вооружения, приносит пользу, они были очень обеспокоены повторным использованием существующих стандартизированных номеров деталей. Один из основных принципов управления конфигурацией гласит, что при изменении формы, соответствия или функции элемента необходимо изменить соответствующий номер элемента. В случае устройств, имитирующих GEM, форма из-за технологии устройства явно меняется, и при этом существует вероятность различий в неуказанных или непроверенных параметрах (функциональные характеристики, электростатический разряд, радиационная стойкость и т. Д.).G-12 единогласно приняла следующее предложение:
Один из основных принципов управления конфигурацией гласит, что при изменении формы, соответствия или функции элемента необходимо изменить соответствующий номер элемента. В случае устройств, имитирующих GEM, форма из-за технологии устройства явно меняется, и при этом существует вероятность различий в неуказанных или непроверенных параметрах (функциональные характеристики, электростатический разряд, радиационная стойкость и т. Д.).G-12 единогласно приняла следующее предложение:
«G-12 не поддерживает продукт более старой технологии, который эмулируется с помощью более новой технологии, имеющей тот же номер детали, потому что он не идентичен по форме, посадке и функциям».
Комитет JC13.2 принял аналогичное предложение в поддержку позиции G-12.
Стандартизация деталей ушла в прошлое?
ВЫВОД:
MIL-PRF-38535 упустил из виду тот факт, что эти детали используются в системах вооружения военного назначения, вместо этого он сосредоточился на производственных системах, используемых для их производства. Спецификация MIL-M-38510 не позволяла деталям, эмулируемым с использованием другой технологии устройства, использовать тот же номер детали, что и оригинальные устройства, однако, похоже, что MIL-PRF-38535 (QML) был написан специально для этого, хотя он нарушает «Лучшие коммерческие практики», как это определено в спецификации EIA для частей AQEC и определениях управления конфигурацией комитетов G-12 и JC13.2.
Спецификация MIL-M-38510 не позволяла деталям, эмулируемым с использованием другой технологии устройства, использовать тот же номер детали, что и оригинальные устройства, однако, похоже, что MIL-PRF-38535 (QML) был написан специально для этого, хотя он нарушает «Лучшие коммерческие практики», как это определено в спецификации EIA для частей AQEC и определениях управления конфигурацией комитетов G-12 и JC13.2.
Если MIL-PRF-38535 не вернется в соответствие с «Лучшими коммерческими методами» и практиками управления конфигурацией, больше производителей прекратят его использовать.
Хотя имитация действительно недоступных для приобретения частей является хорошей вещью для материально-технического обеспечения систем вооружения, эти части не должны иметь одинаковый номер детали, потому что они «просто не одинаковы».
Микросхема агранулярной лобной коры: проверка общности канонической кортикальной микросхемы
Введение
Микросхема агранулярной лобной коры неизвестна. Влиятельная каноническая кортикальная микросхема (CCM) была подробно описана на основе работы, выполняемой в зрительной коре головного мозга кошек и обезьян (Gilbert, 1983; Callaway, 1998; Douglas and Martin, 2004).Отличительные особенности этой хорошо известной модели включают восходящий вход в зернистый слой IV и локальные, зависящие от слоя образцы проекций (рис. 1). Но в отличие от зрительной коры, агранулярные лобные области демонстрируют плохо определяемую ламинарную цитоархитектуру без идентифицируемого слоя IV (Fig. 2). Тем не менее, широко распространено мнение, что CCM, описанный в ранних сенсорных областях, является повсеместным признаком неокортекса. Эта гипотеза направляет влиятельные корковые иерархии (Felleman, Van Essen, 1991; Markov et al., 2011), лежит в основе интерпретации сигнала fMRI BOLD (Logothetis, 2008; Boynton, 2011) и является основополагающим для крупномасштабной реализации функции коры головного мозга, включая амбициозный проект Blue Brain Project (Markram, 2006; Heinzle et al.
Влиятельная каноническая кортикальная микросхема (CCM) была подробно описана на основе работы, выполняемой в зрительной коре головного мозга кошек и обезьян (Gilbert, 1983; Callaway, 1998; Douglas and Martin, 2004).Отличительные особенности этой хорошо известной модели включают восходящий вход в зернистый слой IV и локальные, зависящие от слоя образцы проекций (рис. 1). Но в отличие от зрительной коры, агранулярные лобные области демонстрируют плохо определяемую ламинарную цитоархитектуру без идентифицируемого слоя IV (Fig. 2). Тем не менее, широко распространено мнение, что CCM, описанный в ранних сенсорных областях, является повсеместным признаком неокортекса. Эта гипотеза направляет влиятельные корковые иерархии (Felleman, Van Essen, 1991; Markov et al., 2011), лежит в основе интерпретации сигнала fMRI BOLD (Logothetis, 2008; Boynton, 2011) и является основополагающим для крупномасштабной реализации функции коры головного мозга, включая амбициозный проект Blue Brain Project (Markram, 2006; Heinzle et al. , 2007; Helmstaedter et al., 2007). Если CCM не распространяется на лобную кору, большая часть этой работы может потребовать повторной оценки. Таким образом, исследователи выступают за исследования, чтобы выявить детали микросхем в агранулярных областях (Shipp, 2005).
, 2007; Helmstaedter et al., 2007). Если CCM не распространяется на лобную кору, большая часть этой работы может потребовать повторной оценки. Таким образом, исследователи выступают за исследования, чтобы выявить детали микросхем в агранулярных областях (Shipp, 2005).
Основные характеристики СКК. a , Интерактивные и интерламинарные возбуждающие проекции, выделяющие проекции, которые, как считается, определяют время CSD в определенных пластинках. Для ясности прогнозы пронумерованы в порядке их временного приоритета (адаптировано из Gilbert, 1983). b , Боковые, повторяющиеся возбуждающие и тормозные проекции. Красным цветом изображены ГАМКергические проекции, оказывающие тормозящее влияние. Голубым цветом показаны глутаматергические выступы, оказывающие возбуждающее действие.Пулы пирамидных нейронов поверхностного и глубокого слоев моделируются отдельно, чтобы учесть различия в распределении рецепторов GABA A и GABA B (адаптировано из Douglas and Martin, 1991).
Цитоархитектура агранулярной лобной коры в сравнении со зрительной корой. a , Данные взяты из Paxinos et al. (2000) с разрешения. Срезы подвергали иммуногистохимической реакции для демонстрации белка нейрофиламента SMI32.Сулькальные ориентиры и определенные области помечены для облегчения ориентации (V1, первичная зрительная кора; V2, зрительная область 2). На врезках схем показаны примерные плоскости, с которых были взяты срезы. Области, обведенные синим цветом, увеличиваются справа. V1 может быть четко очерчен пластинками и показывает отчетливый слой IV, разделяющий слои III и V. Напротив, SEF обнаруживает кластеры пирамидных клеток в слое III с менее плотными пирамидными клетками в слое V (Geyer et al., 2000). b , Сравнение ламинарного распределения ацетилхолинэстеразы (AChE), миелиновых волокон и вещества Ниссля в первичной зрительной коре (вверху) и SEF (внизу).Каждая пара представляет собой ткань, взятую у одной и той же обезьяны. Выраженная многослойная структура зрительной коры контрастирует с более однородным внешним видом SEF. Ламинарный паттерн окрашивания AChE в SEF сильно отличается от такового в первичной зрительной коре, будучи наиболее плотным в слое I и плотным также в слоях V и VI. Сходным образом, ламинарный паттерн окрашивания миелиновых волокон в SEF заметно отличается от такового в первичной зрительной коре головного мозга, не имеет ламинации и наиболее плотен только до слоя II.Сечения Ниссля показывают, что SEF полностью отличается от первичной зрительной коры без четкой границы, разделяющей гомогенные слои II и III, которые содержат в основном маленькие пирамиды, за исключением самой нижней части III, в которой присутствуют пирамиды среднего размера. Слой VI содержит в основном веретеновидные клетки и по плотности клеток может быть разделен на поверхностные, относительно редкие и более глубокие, относительно плотные подслои (Matelli et al., 1991).
Выраженная многослойная структура зрительной коры контрастирует с более однородным внешним видом SEF. Ламинарный паттерн окрашивания AChE в SEF сильно отличается от такового в первичной зрительной коре, будучи наиболее плотным в слое I и плотным также в слоях V и VI. Сходным образом, ламинарный паттерн окрашивания миелиновых волокон в SEF заметно отличается от такового в первичной зрительной коре головного мозга, не имеет ламинации и наиболее плотен только до слоя II.Сечения Ниссля показывают, что SEF полностью отличается от первичной зрительной коры без четкой границы, разделяющей гомогенные слои II и III, которые содержат в основном маленькие пирамиды, за исключением самой нижней части III, в которой присутствуют пирамиды среднего размера. Слой VI содержит в основном веретеновидные клетки и по плотности клеток может быть разделен на поверхностные, относительно редкие и более глубокие, относительно плотные подслои (Matelli et al., 1991).
Плотность источника тока (CSD), полученная из потенциала локального поля (LFP), выявляет ламинарную последовательность нейронной активации в корковых слоях (Freeman and Nicholson, 1975; Nicholson and Freeman, 1975; Mitzdorf and Singer, 1978, 1979; Mitzdorf, 1985).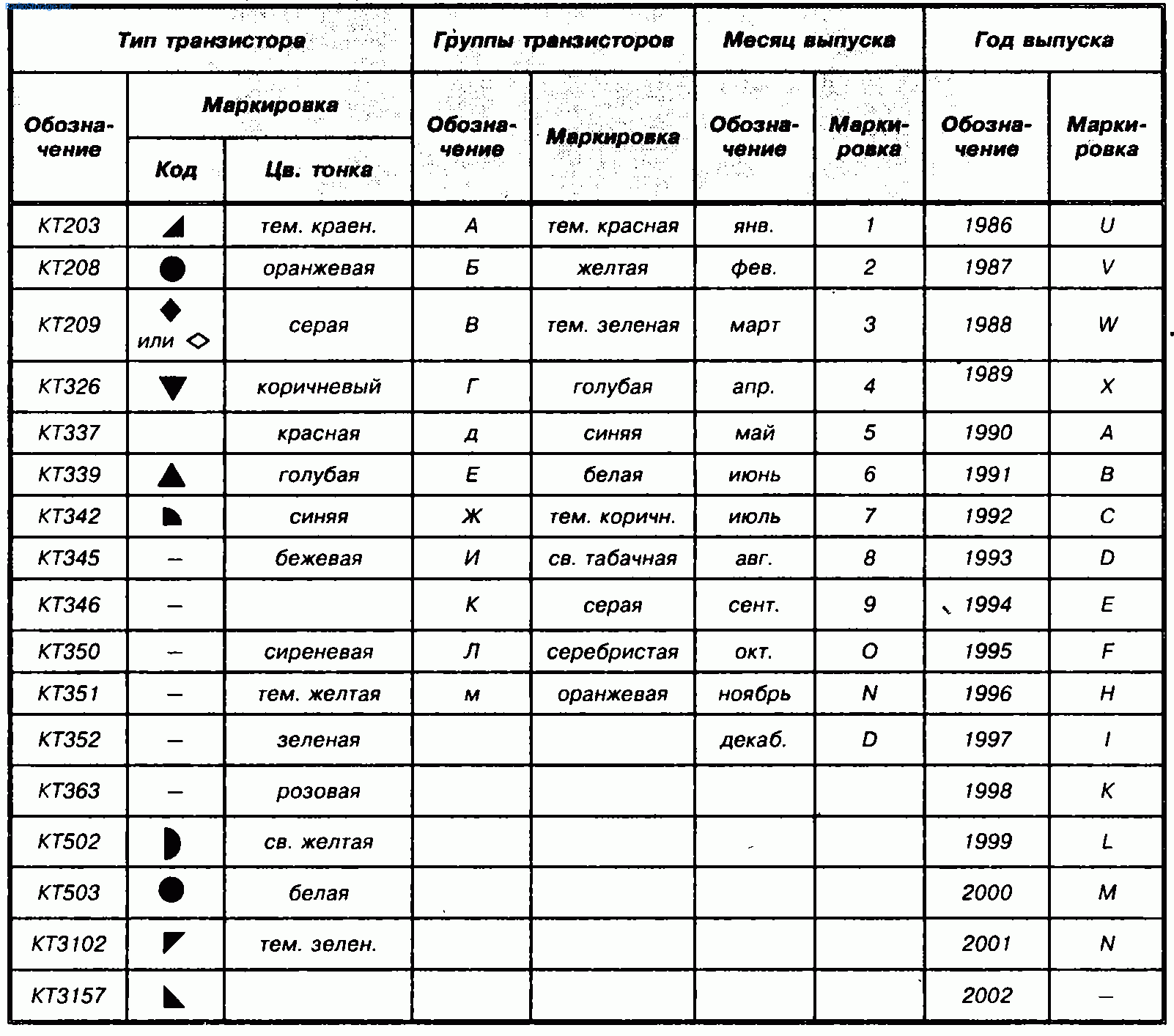 ; Buzsáki et al., 1986). Используя линейные электроды для сравнения LFP на соседних участках, можно удалить потенциалы дальнего поля и ссылочные артефакты и наблюдать локальный ток (Kajikawa and Schroeder, 2011). Этот подход выявляет временную структуру активности корковых слоев по мере того, как ансамбли дендритов последовательно деполяризуются (Di et al., 1990; Schroeder et al., 1998; Lakatos et al., 2007; Lipton et al., 2010). Таким образом, CSD обеспечивает функциональное считывание кортикальных микросхем, охватывая более широкое поле зрения, чем одноэлементный и оптогенетический подходы.В сочетании с измерениями пиковой активности CSD также может выявить ламинарное происхождение одно- и многоединичной активности. Однако те самые свойства, которые делают агранулярную кору полезной для тестирования универсальности CCM, могут сделать записи CSD непрактичными. Подход CSD оказался эффективным в сенсорных областях именно из-за их анатомической структуры; дендриты из нейронных ансамблей ветвятся вместе в четко определенные слои и деполяризуются в унисон, что позволяет наблюдать суммирование тока в мезоскопическом масштабе (Freeman and Nicholson, 1975; Nicholson and Freeman, 1975; Mitzdorf, 1985; Riera et al.
; Buzsáki et al., 1986). Используя линейные электроды для сравнения LFP на соседних участках, можно удалить потенциалы дальнего поля и ссылочные артефакты и наблюдать локальный ток (Kajikawa and Schroeder, 2011). Этот подход выявляет временную структуру активности корковых слоев по мере того, как ансамбли дендритов последовательно деполяризуются (Di et al., 1990; Schroeder et al., 1998; Lakatos et al., 2007; Lipton et al., 2010). Таким образом, CSD обеспечивает функциональное считывание кортикальных микросхем, охватывая более широкое поле зрения, чем одноэлементный и оптогенетический подходы.В сочетании с измерениями пиковой активности CSD также может выявить ламинарное происхождение одно- и многоединичной активности. Однако те самые свойства, которые делают агранулярную кору полезной для тестирования универсальности CCM, могут сделать записи CSD непрактичными. Подход CSD оказался эффективным в сенсорных областях именно из-за их анатомической структуры; дендриты из нейронных ансамблей ветвятся вместе в четко определенные слои и деполяризуются в унисон, что позволяет наблюдать суммирование тока в мезоскопическом масштабе (Freeman and Nicholson, 1975; Nicholson and Freeman, 1975; Mitzdorf, 1985; Riera et al. , 2012). При отсутствии слоя IV и отсутствии четких ламинарных границ агранулярная лобная кора не может производить интерпретируемый CSD.
, 2012). При отсутствии слоя IV и отсутствии четких ламинарных границ агранулярная лобная кора не может производить интерпретируемый CSD.
Чтобы ответить на эти вопросы, мы сообщаем о первом CSD, измеренном в лобной коре. Используя LFP в сочетании с ламинарными одно- и многоэлементными записями, мы охарактеризовали функциональные параметры кортикальной микросхемы в агранулярном дополнительном поле глаза (SEF), хорошо изученной области дорсомедиальной области 6 (Schlag and Schlag-Rey, 1987; Matelli et al. , 1991; Schall, 1991).Хотя многие функции соответствовали СКК, наблюдались также ключевые различия.
Результаты
Используя линейные матрицы микроэлектродов (расстояние между контактами 150 мкм), мы зарегистрировали визуально вызванные и связанные с саккадами LFP и спайки от SEF двух макак. Мы приобрели 12 342 испытания (6448 обезьян E, 5894 обезьян X) за 17 сеансов (7 обезьян E, 10 обезьян X). Количество сеансов аналогично тому, которое использовалось в предыдущих исследованиях, в которых регистрировали CSD из полосатой коры (Maier et al.![]() , 2010; Maier et al., 2011; Spaak et al., 2012), хотя количество испытаний за сеанс несколько больше.
, 2010; Maier et al., 2011; Spaak et al., 2012), хотя количество испытаний за сеанс несколько больше.
SEF был обнаружен с помощью внутрикортикальной электрической микростимуляции для выявления движений глаз (Schlag and Schlag-Rey, 1987; Schall, 1991; Tehovnik et al., 1999; Martinez-Trujillo et al., 2004; рис. 3 a , b). ). Чтобы получить CSD, мы подтвердили, что электродные массивы вошли в кору перпендикулярно поверхности коры с помощью комбинированной МР- и КТ-визуализации (рис. 3 c – j ).После оседания электродной решетки в коре (3–4 ч) мы представили широкопольные (угол обзора 40 × 36 °) световые вспышки (34,80 кд / м 2 ) блоками по 100–200 предъявлений, аналогично стимулы, которые ранее использовались для характеристики ламинарных микросхем в зрительной коре (Schroeder et al., 1998; Maier et al., 2010). Перемежаясь с этими блоками зрительной стимуляции, мы регистрировали активность во время спонтанных движений глаз, производимых, когда обезьяны отдыхали в темноте в течение 5-10 минут.
Расположение и угол проникновения записей. a , b , Карты получены на основе эффектов внутрикортикальной электрической микростимуляции у каждой обезьяны. Слева показано переднее расположение центра камеры. Кружками обозначены отверстия в решетке на расстоянии 1 мм друг от друга. Легенда показывает типы движения, вызванные подаваемым током 50–200 мкА. Перекрестие показывает расположение направляющих трубок на КТ-изображениях справа. c – j , зарегистрированный MR (зеленый), показывающий мягкие ткани, включая серое вещество и белое вещество, с компьютерной томографией (красный), показывающий кость, адаптеры камеры из нержавеющей стали, титановые винты, титановые подголовники, некоторое количество стоматологического акрила, используемого в имплантатах, и направляющие трубы из нержавеющей стали. c , d , Показать корональную и сагиттальную плоскости для обезьяны X.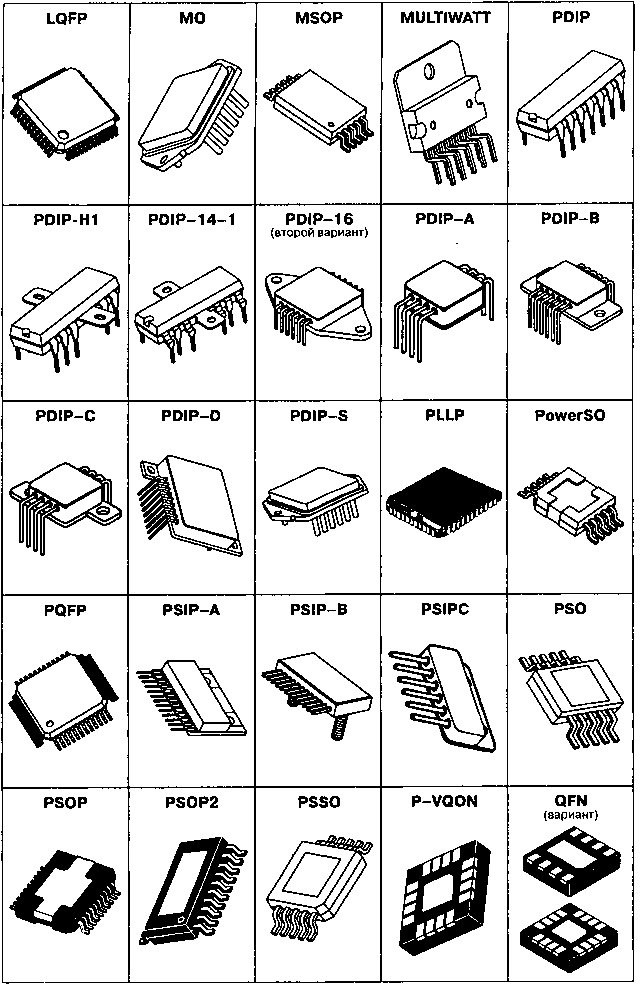 e , f , Показать корональную и сагиттальную плоскости для обезьяны E. Синие квадраты в c – f увеличены в г – дж . Голубые линии в g – j показывают пиальную поверхность и переход от серого вещества к белому. Тонкие желтые линии показывают результат автоматизированного алгоритма, который минимизировал расстояние между пиальной поверхностью и серым веществом для расчета углов, перпендикулярных серому веществу (см. Материалы и методы).Жирными желтыми линиями показана траектория электродных решеток в зависимости от ориентации направляющих трубок. Толстые и тонкие желтые линии практически параллельны в точках въезда. Эта ориентация подтверждает правильность измерения CSD.
e , f , Показать корональную и сагиттальную плоскости для обезьяны E. Синие квадраты в c – f увеличены в г – дж . Голубые линии в g – j показывают пиальную поверхность и переход от серого вещества к белому. Тонкие желтые линии показывают результат автоматизированного алгоритма, который минимизировал расстояние между пиальной поверхностью и серым веществом для расчета углов, перпендикулярных серому веществу (см. Материалы и методы).Жирными желтыми линиями показана траектория электродных решеток в зависимости от ориентации направляющих трубок. Толстые и тонкие желтые линии практически параллельны в точках въезда. Эта ориентация подтверждает правильность измерения CSD.
Однократный сеанс визуально вызванного CSD
На рисунке 4 показаны данные, собранные во время репрезентативного сеанса. Чтобы интерпретировать CSD, необходимо было оценить глубину электродной решетки относительно серого вещества, как описано (см. Материалы и методы). Некоторые физиологические меры предоставили информацию о положении электродов.Во-первых, на поверхностном канале наблюдался артефакт, связанный с сердечным ритмом (далее именуемый артефактом пульса). Этот сигнал указывал, где электрод контактировал либо с твердой мозговой оболочкой, либо с эпидуральным солевым раствором в записывающей камере, который заметно пульсировал вместе с сердцебиением обезьян. Во-вторых, на протяжении всех сеансов мы наблюдали заметное увеличение мощности в диапазоне частот γ (40–80 Гц) на нескольких контактах электродов, которое постепенно уменьшалось в более глубоких местах.Несколько недавних исследований показали повышенную мощность гамма-излучения в поверхностных и средних слоях по сравнению с глубокими слоями (Maier et al., 2010; Xing et al., 2012; Smith and Sommer, 2013), предполагая, что эта мера является еще одним полезным маркером для оценки глубины . Наконец, мы записали хорошо изолированные единичные единицы одновременно с LFP и совместили их положение с маркерами, описанными выше.
Материалы и методы). Некоторые физиологические меры предоставили информацию о положении электродов.Во-первых, на поверхностном канале наблюдался артефакт, связанный с сердечным ритмом (далее именуемый артефактом пульса). Этот сигнал указывал, где электрод контактировал либо с твердой мозговой оболочкой, либо с эпидуральным солевым раствором в записывающей камере, который заметно пульсировал вместе с сердцебиением обезьян. Во-вторых, на протяжении всех сеансов мы наблюдали заметное увеличение мощности в диапазоне частот γ (40–80 Гц) на нескольких контактах электродов, которое постепенно уменьшалось в более глубоких местах.Несколько недавних исследований показали повышенную мощность гамма-излучения в поверхностных и средних слоях по сравнению с глубокими слоями (Maier et al., 2010; Xing et al., 2012; Smith and Sommer, 2013), предполагая, что эта мера является еще одним полезным маркером для оценки глубины . Наконец, мы записали хорошо изолированные единичные единицы одновременно с LFP и совместили их положение с маркерами, описанными выше. Этот набор различных физиологических сигналов предоставил сходные данные для оценки положения электрода с учетом ламинарной глубины, которая была назначена с помощью процедуры автоматического выравнивания (описанной в разделе «Материалы и метод» и ниже).
Этот набор различных физиологических сигналов предоставил сходные данные для оценки положения электрода с учетом ламинарной глубины, которая была назначена с помощью процедуры автоматического выравнивания (описанной в разделе «Материалы и метод» и ниже).
Необработанные и обработанные данные репрезентативного сеанса (1316 испытаний). a , Схематическая диаграмма электродной решетки, нарисованная в масштабе и расположенная на разрезе Ниссля из SEF (адаптировано из Matelli et al., 1991 с разрешения). b , Три секунды необработанного LFP, записанного с каждого из контактов. Красный график (восьмой сверху) показывает артефакт пульса. c , полосовой фильтр LFP от 40 до 80 Гц. Синие кривые показывают активность γ, превышающую среднее значение. d , Нормализованная средняя мощность γ, записанная на каждом контакте электрода во время сеанса (синий), по сравнению со средней мощностью γ, записанной для всех контактов (вертикальная черная линия). Обратите внимание на выраженное увеличение мощности γ на контактах в нейропиле. e , Суммарный рисунок, показывающий глубину артефакта пульса (красная линия), повышенная мощность γ (синяя линия) и количество хорошо изолированных одиночных единиц (черные треугольники), записанных одновременно. Во время этого сеанса мы записали 29 хорошо изолированных одиночных устройств: 2 устройства на 5 каналах и 3 устройства на других 5 каналах. f , Триста миллисекунд LFP, связанного с событием, выровнены со стимулом вспышки (вертикальная черная линия). Обратите внимание на изменение полярности напряжения в канале с артефактом импульса. Выше этого канала сигнал представляет собой объемную ЭЭГ в физиологическом растворе, заполняющем камеру записи. Ниже этого канала сигналы представляют собой электрокортикограмму, записанную с пиальной поверхности, и LFP, записанные с серого вещества. г , Триста миллисекунд CSD, полученных из LFP, интерполяция между контактами с разрешением 10 мкм.
Обратите внимание на выраженное увеличение мощности γ на контактах в нейропиле. e , Суммарный рисунок, показывающий глубину артефакта пульса (красная линия), повышенная мощность γ (синяя линия) и количество хорошо изолированных одиночных единиц (черные треугольники), записанных одновременно. Во время этого сеанса мы записали 29 хорошо изолированных одиночных устройств: 2 устройства на 5 каналах и 3 устройства на других 5 каналах. f , Триста миллисекунд LFP, связанного с событием, выровнены со стимулом вспышки (вертикальная черная линия). Обратите внимание на изменение полярности напряжения в канале с артефактом импульса. Выше этого канала сигнал представляет собой объемную ЭЭГ в физиологическом растворе, заполняющем камеру записи. Ниже этого канала сигналы представляют собой электрокортикограмму, записанную с пиальной поверхности, и LFP, записанные с серого вещества. г , Триста миллисекунд CSD, полученных из LFP, интерполяция между контактами с разрешением 10 мкм. Вертикальная черная линия показывает начало вспышки.
Вертикальная черная линия показывает начало вспышки.
На основе известной толщины отдельных слоев в SEF (Мателли и др., 1991) мы оценили ламинарные границы и отнесли визуально вызванные стоки тока к определенным слоям (см. Материалы и методы). В репрезентативном сеансе записи наибольший сток (минимум = -42 нА / мм 3 ) произошел в слое III, начиная примерно через 50 мс после предъявления стимула. Второй сток (минимум = -25 нА / мм 3 ) начался через несколько миллисекунд в слое V.Более позднее погружение (минимум = -23 нА / мм 3 ) произошло более поверхностно в слоях I / II, а дополнительные более слабые погружения (минимум = -20 нА / мм 3 ) были очевидны в слое VI.
Также очевидно несколько источников тока, в том числе один на уровне артефакта пульса. В соответствии с предыдущими исследованиями мы интерпретируем этот источник как пассивный ток, возвращающийся к нижним стокам, поскольку он был зарегистрирован на том же уровне, что и импульсный артефакт, и, следовательно, не может иметь коркового происхождения (Mitzdorf, 1985). В общем, источники тока могут быть вызваны либо пассивным обратным током, либо дендритной гиперполяризацией, что затрудняет их интерпретацию, чем стоки тока (Nicholson and Freeman, 1975; Mitzdorf, 1985). Поэтому в оставшейся части исследования мы сосредоточимся на текущих стоках.
В общем, источники тока могут быть вызваны либо пассивным обратным током, либо дендритной гиперполяризацией, что затрудняет их интерпретацию, чем стоки тока (Nicholson and Freeman, 1975; Mitzdorf, 1985). Поэтому в оставшейся части исследования мы сосредоточимся на текущих стоках.
Чтобы увеличить SNR, мы усреднили вызванные паттерны CSD по сеансам записи, аналогично созданию больших средних ERP из данных ЭЭГ, как это делалось ранее (Maier et al., 2010; Riera et al., 2012). Чтобы сделать это беспристрастным, управляемым данными способом, мы разработали процедуру автоматического выравнивания глубины, чтобы максимизировать сходство между сеансами записи с использованием всей ламинарной структуры источника и стока и времени.Это математически оптимизированное решение для выравнивания глубины основано на единственном предположении, что есть надежные сходства в CSD, измеренные во время сеансов записи (см. Материалы и методы). На рисунке 5 показаны результаты этой процедуры с данными каждого сеанса записи. Абсолютная величина CSD варьировалась день ото дня и у разных обезьян, но в каждом сеансе мы наблюдали четкое снижение в слое III через ~ 50 мс после визуального стимула (рис. 5 b ). В 15 из 17 сеансов (88%) мы также наблюдали второй сток в слое V, а в 16 из 17 сеансов (94%) мы наблюдали третий сток в слоях I / II.На нескольких сеансах записи мы размещали электродную решетку слишком поверхностно, чтобы взять образец из слоя VI, но мы наблюдали опускание в этом месте в 11 из 13 сеансов (85%). Эти согласованности подтверждают предположение, что CSD надежен для сеансов записи и обезьян. Примечательно, что метод автоматического выравнивания не учитывал физиологические сигналы, описанные выше (то есть импульсный артефакт, мощность LFP γ и местоположения отдельных единиц), потому что он полагался исключительно на данные CSD. Тем не менее, мы наблюдали тесное соответствие между его оценками глубины коры головного мозга и другими физиологическими сигналами, наблюдаемыми в необработанных данных, что еще раз подтверждает точность этого подхода (рис.
Абсолютная величина CSD варьировалась день ото дня и у разных обезьян, но в каждом сеансе мы наблюдали четкое снижение в слое III через ~ 50 мс после визуального стимула (рис. 5 b ). В 15 из 17 сеансов (88%) мы также наблюдали второй сток в слое V, а в 16 из 17 сеансов (94%) мы наблюдали третий сток в слоях I / II.На нескольких сеансах записи мы размещали электродную решетку слишком поверхностно, чтобы взять образец из слоя VI, но мы наблюдали опускание в этом месте в 11 из 13 сеансов (85%). Эти согласованности подтверждают предположение, что CSD надежен для сеансов записи и обезьян. Примечательно, что метод автоматического выравнивания не учитывал физиологические сигналы, описанные выше (то есть импульсный артефакт, мощность LFP γ и местоположения отдельных единиц), потому что он полагался исключительно на данные CSD. Тем не менее, мы наблюдали тесное соответствие между его оценками глубины коры головного мозга и другими физиологическими сигналами, наблюдаемыми в необработанных данных, что еще раз подтверждает точность этого подхода (рис. 5 c ). Дополнительным доказательством точности этой процедуры автоматического выравнивания являются наши выводы о нейронных ответах, которые различаются по слоям в SEF (см. Ниже).
5 c ). Дополнительным доказательством точности этой процедуры автоматического выравнивания являются наши выводы о нейронных ответах, которые различаются по слоям в SEF (см. Ниже).
Результаты автоматизированной процедуры согласования записей U-зонда по сессиям. A , CSD, вызываемая визуально, записывается индивидуально для каждого сеанса. Обезьяна и место каждого проникновения указаны выше. Записи выровнены слева направо в хронологическом порядке для каждой обезьяны.Цифры в верхней части указывают масштабы карт CSD (нА / мм 3 ). Горизонтальные черные полосы указывают предполагаемое среднее местоположение серого вещества на основе физиологических сигналов. b , Визуально вызванные CSD замаскированы, чтобы показать местоположения четырех усредненных визуально вызванных поглотителей, представленных на Рисунке 6. Обратите внимание на близкое соответствие в расположении этих поглотителей во время сеансов записи, демонстрирующее сходство в CSD, записанное в последующие дни, и успех нашей автоматической процедуры центровки. c , Физиологические сигналы, очевидные в необработанных данных по отдельным сеансам. Импульсный артефакт (красные линии), повышенная гамма-активность (синие линии) и отдельные единицы (черные треугольники) хорошо согласуются с нашей оценкой местоположения серого вещества (серая заливка).
c , Физиологические сигналы, очевидные в необработанных данных по отдельным сеансам. Импульсный артефакт (красные линии), повышенная гамма-активность (синие линии) и отдельные единицы (черные треугольники) хорошо согласуются с нашей оценкой местоположения серого вещества (серая заливка).
Гранд-средний, визуально вызываемый CSD
Чтобы гарантировать, что данные от обеих обезьян вносят равный вклад в среднюю CSD, сохраняя при этом ее абсолютную величину, мы нормализовали данные таким образом, чтобы SD средней CSD каждой обезьяны совпадала со SD средней CSD.Мы подтвердили, что такая же качественная картина результатов наблюдалась, когда данные не были нормализованы, но сообщаем результаты этого подхода, поскольку он дает среднее значение, менее подверженное влиянию выбросов. Визуально вызванный сток тока наблюдался на глубине записи 2 мм (Рис. 6 a ). Из-за расположения электродной решетки более глубокие каналы отбирались реже, чем поверхностные каналы ( N = 17 каналов 1–12, N = 15 каналов 13, N = 13 каналов 14–15, N = 11 канал 16). Пиковая величина -25 нА / мм 3 составляет лишь ~ 15% от величины визуально вызванного потока тока, зарегистрированного для первичной зрительной коры с использованием аналогичных параметров стимуляции и методов записи и анализа (Maier et al., 2010). Тем не менее, была очевидна четкая ламинарная последовательность стоков тока. Два начальных опускания наблюдались в большом среднем CSD, предполагая, что зрительные афференты оканчиваются двумя отдельными пластинками. Эти данные согласуются с опубликованными анатомическими данными (Matelli et al., 1991).Хотя в нем отсутствует зернистый слой, SEF содержит два слоя относительно плотных пирамидных клеток, один в глубоком слое III и второй формирующий слой V, которые оба получают зрительные афференты (Maioli et al., 1998; Shipp et al., 1998 см. Обсуждение. ).
Пиковая величина -25 нА / мм 3 составляет лишь ~ 15% от величины визуально вызванного потока тока, зарегистрированного для первичной зрительной коры с использованием аналогичных параметров стимуляции и методов записи и анализа (Maier et al., 2010). Тем не менее, была очевидна четкая ламинарная последовательность стоков тока. Два начальных опускания наблюдались в большом среднем CSD, предполагая, что зрительные афференты оканчиваются двумя отдельными пластинками. Эти данные согласуются с опубликованными анатомическими данными (Matelli et al., 1991).Хотя в нем отсутствует зернистый слой, SEF содержит два слоя относительно плотных пирамидных клеток, один в глубоком слое III и второй формирующий слой V, которые оба получают зрительные афференты (Maioli et al., 1998; Shipp et al., 1998 см. Обсуждение. ).
Гранд-среднее, визуально вызванное (слева) и связанное с саккадами (справа) CSD из SEF. Сечение Ниссля из SEF в центре указывает на ламинарную архитектуру (адаптировано из Matelli et al. , 1991 с разрешения). a , CSD, записанный, когда обезьяны пассивно наблюдали широкопольные вспышки света.Наблюдалось четыре стока тока, которые пронумерованы в порядке появления. b , запись CSD, когда обезьяны совершали спонтанные саккады в темноте. c , d , Данные из a и b воспроизведены без интерполяции, с выделением периодов времени, когда каналы значительно отклоняются от базового уровня (выполняется Wilcoxon, p <0,05 для> 5 последовательных Интервалы времени 10 мс).Показаны шестнадцать глубин канала.
, 1991 с разрешения). a , CSD, записанный, когда обезьяны пассивно наблюдали широкопольные вспышки света.Наблюдалось четыре стока тока, которые пронумерованы в порядке появления. b , запись CSD, когда обезьяны совершали спонтанные саккады в темноте. c , d , Данные из a и b воспроизведены без интерполяции, с выделением периодов времени, когда каналы значительно отклоняются от базового уровня (выполняется Wilcoxon, p <0,05 для> 5 последовательных Интервалы времени 10 мс).Показаны шестнадцать глубин канала.
Мы использовали два метода для измерения задержки начала поглощения на каждом уровне (см. Материалы и методы). Во-первых, мы оценили отличия от исходного уровня, используя текущий подход Вилкоксона. Этот метод довольно чувствителен, но дает только одну меру задержки между сеансами для каждого уровня. Первый сток (мин -25 нА / мм 3 ) появился в слое III через 51 мс после стимула и стал максимальным через 72 мс. Второй сток (мин -22 нА / мм 3 ) развивался в слое V за 55 мс, становясь максимальным через 105 мс.Последующие понижения произошли в слоях I / II (мин. −14 нА / мм 3 ) при 147 мс (пик на 168 мс) и в слое VI (мин. −10 нА / мм 3 ) при 172 мс (пик при 173 мс). РС). Мы также измерили латентность начала ламинарных поглотителей в отдельных сеансах, записав время, когда поглотитель достигал максимума 33%. Хотя этот метод несколько менее чувствителен, так как не может идентифицировать малозаметные начала, он дает полезную оценку вариабельности между сеансами записи. Используя этот подход, мы записали следующие средние задержки ± SEM: слой III 50 ± 3 мс, слой V 73 ± 7 мс, слои I / II 100 ± 12 мс, слой VI 159 ± 25 мс.
Первый сток (мин -25 нА / мм 3 ) появился в слое III через 51 мс после стимула и стал максимальным через 72 мс. Второй сток (мин -22 нА / мм 3 ) развивался в слое V за 55 мс, становясь максимальным через 105 мс.Последующие понижения произошли в слоях I / II (мин. −14 нА / мм 3 ) при 147 мс (пик на 168 мс) и в слое VI (мин. −10 нА / мм 3 ) при 172 мс (пик при 173 мс). РС). Мы также измерили латентность начала ламинарных поглотителей в отдельных сеансах, записав время, когда поглотитель достигал максимума 33%. Хотя этот метод несколько менее чувствителен, так как не может идентифицировать малозаметные начала, он дает полезную оценку вариабельности между сеансами записи. Используя этот подход, мы записали следующие средние задержки ± SEM: слой III 50 ± 3 мс, слой V 73 ± 7 мс, слои I / II 100 ± 12 мс, слой VI 159 ± 25 мс.
Для количественной оценки этих наблюдений мы разделили временной ход после визуального представления на раннюю (51–150 мс) и позднюю (151–250 мс) эпохи и провели иерархический дисперсионный анализ с повторными измерениями с использованием слоев, эпох и обезьян в качестве фиксированных факторов. и номер сеанса как случайный фактор, вложенный в фиксированный фактор обезьяны (Self et al., 2013). Мы наблюдали значительные различия в CSD по слоям ( F (3,129) = 9,87, p = 4,73 × 10 −5 ) и уменьшение CSD по слоям в позднюю эпоху ( F ( 1,129) = 166.28, p = 1,51 × 10 −10 ). Также наблюдался значительный главный эффект между обезьянами, показывающий, что CSD имел тенденцию быть больше по величине для обезьяны E ( F (1,129) = 21,91, p = 2,75 × 10 −4 ). Не было отмечено значительных эффектов для вложенного случайного фактора числа проникновений ( F (15,129) = 0,57, p = 0,86). Важно отметить, что значительное взаимодействие между слоями и временными периодами наблюдалось в CSD большого среднего ( F (3,129) = 21.96, p = 6.55 × 10 −9 ). В качестве дополнительного теста мы провели межсессионные тесты Вилкоксона на каналах, разделенных расстоянием между электродами (150 мкм).
и номер сеанса как случайный фактор, вложенный в фиксированный фактор обезьяны (Self et al., 2013). Мы наблюдали значительные различия в CSD по слоям ( F (3,129) = 9,87, p = 4,73 × 10 −5 ) и уменьшение CSD по слоям в позднюю эпоху ( F ( 1,129) = 166.28, p = 1,51 × 10 −10 ). Также наблюдался значительный главный эффект между обезьянами, показывающий, что CSD имел тенденцию быть больше по величине для обезьяны E ( F (1,129) = 21,91, p = 2,75 × 10 −4 ). Не было отмечено значительных эффектов для вложенного случайного фактора числа проникновений ( F (15,129) = 0,57, p = 0,86). Важно отметить, что значительное взаимодействие между слоями и временными периодами наблюдалось в CSD большого среднего ( F (3,129) = 21.96, p = 6.55 × 10 −9 ). В качестве дополнительного теста мы провели межсессионные тесты Вилкоксона на каналах, разделенных расстоянием между электродами (150 мкм). Все четыре стока тока значительно отличались от базовой линии (рис. 6 c ). Таким образом, даже несмотря на то, что плотность тока была слабее, чем плотность тока, наблюдаемая в ранней сенсорной коре, картина синаптического тока в средних слоях, за которым следовали ток в поверхностных и глубоких слоях, была согласована на всех сессиях и была подобна, хотя и не идентична CSD, полученной в ранней сенсорной кора головного мозга (Mitzdorf, Singer, 1978; Schroeder et al., 1998; Lakatos et al., 2007; Lipton et al., 2010; Riera et al., 2012).
Все четыре стока тока значительно отличались от базовой линии (рис. 6 c ). Таким образом, даже несмотря на то, что плотность тока была слабее, чем плотность тока, наблюдаемая в ранней сенсорной коре, картина синаптического тока в средних слоях, за которым следовали ток в поверхностных и глубоких слоях, была согласована на всех сессиях и была подобна, хотя и не идентична CSD, полученной в ранней сенсорной кора головного мозга (Mitzdorf, Singer, 1978; Schroeder et al., 1998; Lakatos et al., 2007; Lipton et al., 2010; Riera et al., 2012).
CSD, относящаяся к саккаде
Чтобы определить, является ли этот паттерн текущих стоков специфичным для визуального ввода или возникает с другими событиями, во время которых модулируется SEF, мы вывели CSD, связанный с самогенерируемыми саккадическими движениями глаз в темноте. CSD, связанный с саккадами, на отдельных сессиях был очень слабым, но после выравнивания и усреднения мы наблюдали отчетливые опускания (Рис. 6 b , d ).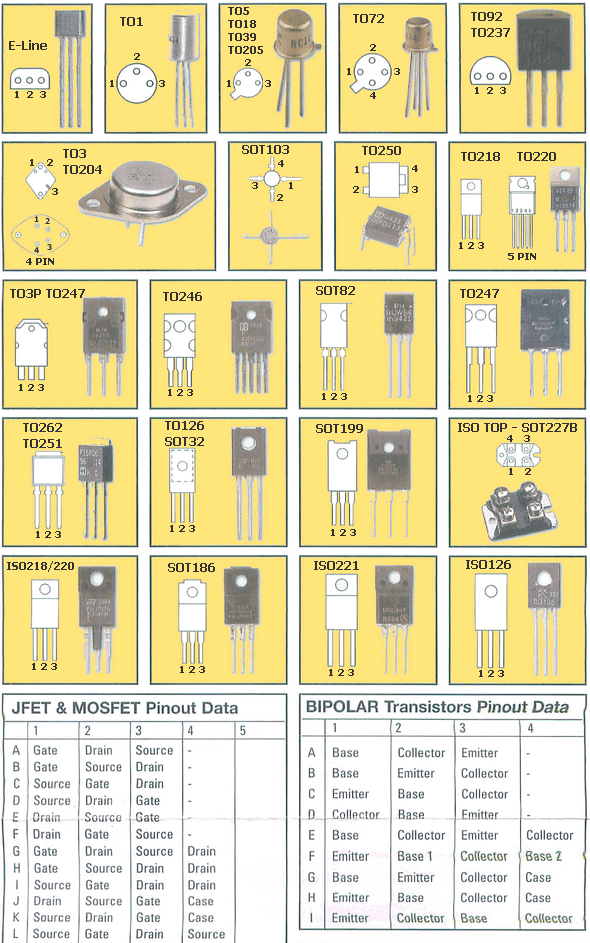

Единичные единицы, зарегистрированные с помощью линейной матрицы микроэлектродов
Мы зарегистрировали пиковую активность 295 отдельных единиц одновременно с LFP (115 обезьян E, 180 обезьян X). Немногие исследователи сообщили о единичных данных, записанных с помощью недавно разработанной электродной решетки, используемой в текущем исследовании (Hansen and Dragoi, 2011; Hansen et al., 2012). Поэтому мы включаем описание метрик всплесков, включая качество изоляции с использованием ложноположительных и ложноотрицательных оценок, и описание форм всплесков по глубине, а также меры ширины всплесков и изменчивости всплесков в зависимости от глубины записи.
На рисунке 7 показано пространство анализа главных компонентов (PCA) и соответствующие формы сигналов из примеров сеансов, взятых от каждой обезьяны. Эти данные показывают, что электродная решетка способна эффективно изолировать отдельные блоки.Мы количественно оценили изоляцию, оценив количество ложноположительных и ложноотрицательных пиков, обнаруженных для каждой предполагаемой единичной единицы (Hill et al. , 2011a). В целом 88% предполагаемых единичных единиц дали ложноположительные результаты ниже 10%, а 95% дали ложноположительные результаты ниже 20%. Кроме того, 53% предполагаемых единичных единиц дали ложноотрицательные показатели ниже 10%, а 81% дали ложноотрицательные показатели ниже 20%. Эти результаты сопоставимы с показателями изоляции, полученными в других исследованиях с использованием тех же методов (Hill et al., 2011b; Jacobs et al., 2013). Тем не менее, частота ложноположительных результатов несколько ниже, что свидетельствует о том, что наши единичные данные содержали очень мало ложноположительных результатов, а частота ложноотрицательных результатов несколько выше, чем те, о которых сообщалось ранее, что позволяет предположить, что мы недооценили нейронный выброс с помощью строгих критериев сортировки. В совокупности эти меры предполагают, что мы приняли консервативный подход при назначении пиков для единичных кластеров. Низкий уровень ложноположительных результатов подтверждает наше утверждение о том, что единичные записи отражают активность отдельных нейронов, но относительно высокий уровень ложноотрицательных результатов говорит о том, что мы недооценили частоту срабатывания некоторых из этих нейронов.
, 2011a). В целом 88% предполагаемых единичных единиц дали ложноположительные результаты ниже 10%, а 95% дали ложноположительные результаты ниже 20%. Кроме того, 53% предполагаемых единичных единиц дали ложноотрицательные показатели ниже 10%, а 81% дали ложноотрицательные показатели ниже 20%. Эти результаты сопоставимы с показателями изоляции, полученными в других исследованиях с использованием тех же методов (Hill et al., 2011b; Jacobs et al., 2013). Тем не менее, частота ложноположительных результатов несколько ниже, что свидетельствует о том, что наши единичные данные содержали очень мало ложноположительных результатов, а частота ложноотрицательных результатов несколько выше, чем те, о которых сообщалось ранее, что позволяет предположить, что мы недооценили нейронный выброс с помощью строгих критериев сортировки. В совокупности эти меры предполагают, что мы приняли консервативный подход при назначении пиков для единичных кластеров. Низкий уровень ложноположительных результатов подтверждает наше утверждение о том, что единичные записи отражают активность отдельных нейронов, но относительно высокий уровень ложноотрицательных результатов говорит о том, что мы недооценили частоту срабатывания некоторых из этих нейронов. В итоге эти результаты показывают, что U-зонд способен изолировать активность отдельных нейронов с характеристиками, сопоставимыми с характеристиками традиционных вольфрамовых электродов.
В итоге эти результаты показывают, что U-зонд способен изолировать активность отдельных нейронов с характеристиками, сопоставимыми с характеристиками традиционных вольфрамовых электродов.
Пример сигналов и прогнозируемое пространство PCA для восьми отсортированных каналов. Серые облака представляют собой многокомпонентную активность (несортированные переходы через порог). Каналы 1–4 взяты из записей с обезьяной X, а каналы 5–8 – из записей с обезьяной E.
Амплитуда пиков не может использоваться в качестве надежного показателя морфологии нейронов, поскольку на этот показатель влияют как размер нейрона, так и его близость к регистрирующему электроду, но другие показатели формы волны могут дать представление.Например, количество пиков в импульсной форме волны может указывать, какой сегмент нейрона участвовал в записи. Двухфазные волновые формы распространены и, как правило, регистрируются от тел клеток или начального сегмента аксонного холма, тогда как трехфазные волновые формы редко встречаются в нейропиле, поскольку они обычно записываются с аксонов (см. Обзор Lemon, 1984). Мы отметили значительную долю трехфазных сигналов при записи с помощью U-зонда (39 из 295, 13%). Эти агрегаты обычно были временными и не изолировались надолго.Это отличительный признак записи аксонов, потому что внеклеточные потенциалы аксонов во много раз меньше, чем генерируемые клеточными сомами, и поэтому электрод должен располагаться очень близко к аксону, чтобы обнаружить его активность (Lemon, 1984). Мы предполагаем, что U-зонд должен подходить для регистрации активности аксонов из-за геометрии контактов электродов. В отличие от традиционных вольфрамовых микроэлектродов с острой записывающей поверхностью, гладкие электродные контакты на стержне U-образного зонда можно разместить в непосредственной близости от аксона, не прокалывая и не повреждая его.Мы количественно оценили количество двухфазных и трехфазных одиночных нейронов, зарегистрированных для каждого слоя коры и белого вещества (см. Материалы и методы). Из 36 отдельных единиц, записанных со слоя II, ни один не отображал трехфазные формы волны.
 Очень небольшая часть (5 из 116, 4%) одиночных единиц, зарегистрированных в слое III, отображала трехфазные формы волны. Большие доли трехфазных сигналов были обнаружены в слоях V (11 из 90, 12%) и VI (11 из 31, 35%). В белом веществе большинство волн были трехфазными (12 из 22, 55%), что позволяет предположить, что многие из этих ответов были записаны от аксонов, а меньшее – от интерстициальных нейронов непосредственно под слоем VI (Suárez-Solá et al., 2009). Поскольку было невозможно определить слой, откуда произошли эти предполагаемые аксоны, мы исключили их из дальнейшего анализа.
Очень небольшая часть (5 из 116, 4%) одиночных единиц, зарегистрированных в слое III, отображала трехфазные формы волны. Большие доли трехфазных сигналов были обнаружены в слоях V (11 из 90, 12%) и VI (11 из 31, 35%). В белом веществе большинство волн были трехфазными (12 из 22, 55%), что позволяет предположить, что многие из этих ответов были записаны от аксонов, а меньшее – от интерстициальных нейронов непосредственно под слоем VI (Suárez-Solá et al., 2009). Поскольку было невозможно определить слой, откуда произошли эти предполагаемые аксоны, мы исключили их из дальнейшего анализа.
Ширина спайка – еще один полезный показатель формы волны, который использовался для классификации нейронов как предполагаемых пирамидных клеток или предполагаемых интернейронов (Constantinidis et al., 2002; Barthó et al., 2004; Mitchell et al., 2007; Cohen et al., 2009; но см. Vigneswaran et al., 2011). Мы измерили ширину пиков, исключая блоки с трехфазной формой волны (Mitchell et al., 2007). Общее распределение ширины шипов было похоже на то, что описано для V4 (Mitchell et al. , 2007) и лобного поля глаза (FEF; Cohen et al., 2009) с использованием аналогичных критериев (рис. 8). Мы оценили изменчивость скорости стрельбы путем расчета коэффициентов вариации, измеренных на интервалах между спайками (Cohen et al., 2009). В соответствии с гипотетической ассоциацией между узкими пиками и интернейронами, мы обнаружили, что единицы с узкими пиками демонстрируют большую изменчивость во времени пиков, чем единицы с широкими пиками [блок с широким пиком N = 203, блок с узким пиком N = 53, Сумма рангов Уилкоксона W = 23971, p = 1.06 × 10 −5 ]. Этот результат был идентичным, когда мы ограничили наш анализ отдельными единицами с ложноположительными показателями <5% [единица с широким пиком N = 170, единица с узким пиком N = 36, сумма рангов Уилкоксона W = 16214, p = 2,15 × 10 -5 ]. Эти результаты предоставляют новые доказательства, подтверждающие биофизическое различие между узкими и широкими шипами единиц в агранулярной коре.
, 2007) и лобного поля глаза (FEF; Cohen et al., 2009) с использованием аналогичных критериев (рис. 8). Мы оценили изменчивость скорости стрельбы путем расчета коэффициентов вариации, измеренных на интервалах между спайками (Cohen et al., 2009). В соответствии с гипотетической ассоциацией между узкими пиками и интернейронами, мы обнаружили, что единицы с узкими пиками демонстрируют большую изменчивость во времени пиков, чем единицы с широкими пиками [блок с широким пиком N = 203, блок с узким пиком N = 53, Сумма рангов Уилкоксона W = 23971, p = 1.06 × 10 −5 ]. Этот результат был идентичным, когда мы ограничили наш анализ отдельными единицами с ложноположительными показателями <5% [единица с широким пиком N = 170, единица с узким пиком N = 36, сумма рангов Уилкоксона W = 16214, p = 2,15 × 10 -5 ]. Эти результаты предоставляют новые доказательства, подтверждающие биофизическое различие между узкими и широкими шипами единиц в агранулярной коре. В объединенной выборке единичных единиц мы обнаружили небольшую, но значимую корреляцию между шириной пика и глубиной записи ( r (254) = 0.15 р = 0,01). Отдельный регрессионный анализ единиц с узкими и широкими пиками показал, что единицы с широкими пиками были основным источником этого эффекта. Единицы с широкими иглами увеличивались по ширине с увеличением глубины записи ( r (201) = 0,42 p = 6,47 × 10 −10 ), в то время как единицы с узкими иглами остались той же ширины по глубине ( r (51) = 0,07 p = 0,60). Эти результаты были идентичны, когда мы ограничили наш анализ единицами с ложноположительными показателями <5% (единица с широкими пиками r (168) = 0.44 p = 1,77 × 10 −9 ; узел с узким шипом r (34) = 0,13 p = 0,46). Обнаружение увеличенной ширины шипа для нейронов с широким шипом в зависимости от глубины согласуется с анатомией; большие пирамидные нейроны более распространены в более глубоких слоях.
В объединенной выборке единичных единиц мы обнаружили небольшую, но значимую корреляцию между шириной пика и глубиной записи ( r (254) = 0.15 р = 0,01). Отдельный регрессионный анализ единиц с узкими и широкими пиками показал, что единицы с широкими пиками были основным источником этого эффекта. Единицы с широкими иглами увеличивались по ширине с увеличением глубины записи ( r (201) = 0,42 p = 6,47 × 10 −10 ), в то время как единицы с узкими иглами остались той же ширины по глубине ( r (51) = 0,07 p = 0,60). Эти результаты были идентичны, когда мы ограничили наш анализ единицами с ложноположительными показателями <5% (единица с широкими пиками r (168) = 0.44 p = 1,77 × 10 −9 ; узел с узким шипом r (34) = 0,13 p = 0,46). Обнаружение увеличенной ширины шипа для нейронов с широким шипом в зависимости от глубины согласуется с анатомией; большие пирамидные нейроны более распространены в более глубоких слоях. Частота появления нейронов с узкими шипами соответствовала плотности парвальбумина, но не нейронов, экспрессирующих кальретинин или кальбиндин, в SEF (рис. 9). Это открытие открывает интересную возможность того, что единицы с узкими шипами могут в первую очередь быть интернейронами, экспрессирующими парвальбумин, так как они обнаруживаются во всех слоях SEF и имеют тенденцию иметь более крупные клеточные тела.
Частота появления нейронов с узкими шипами соответствовала плотности парвальбумина, но не нейронов, экспрессирующих кальретинин или кальбиндин, в SEF (рис. 9). Это открытие открывает интересную возможность того, что единицы с узкими шипами могут в первую очередь быть интернейронами, экспрессирующими парвальбумин, так как они обнаруживаются во всех слоях SEF и имеют тенденцию иметь более крупные клеточные тела.
Биофизические характеристики единичных единиц. a , Все двухфазные сигналы в популяции. Красный и синий обозначают узкие и широкие пики соответственно (разделенные на 250 мкс). Этот цветовой код такой же в остальных частях. b , Кумулятивные распределения вероятностей CV межспайковых интервалов, построенные отдельно для единиц с широкими и узкими иглами. Единицы с узкими пиками показали большую изменчивость во времени пиков. c , Ширина пика как функция глубины записи.График разброса показывает регрессию ширины шипа по глубине, разделенных популяциями с широким и узким шипом. Гистограммы показывают распределение единиц с широкими и узкими пиками по каждому измерению. Ширина широких шипов увеличивалась с глубиной. Для единиц с узкими шипами такой тенденции не наблюдалось. d , Подсчет единиц с широкими и узкими иглами, зарегистрированными в каждом слое.
Гистограммы показывают распределение единиц с широкими и узкими пиками по каждому измерению. Ширина широких шипов увеличивалась с глубиной. Для единиц с узкими шипами такой тенденции не наблюдалось. d , Подсчет единиц с широкими и узкими иглами, зарегистрированными в каждом слое.
Распределение ГАМКергических интернейронов в SEF. Слева: корональные срезы через SEF, иммуногистохимически прореагировавшие на указанные кальций-связывающие белки.Справа: график расположения положительных нейронов, идентифицированных с использованием процедуры полуавтоматической классификации (см. «Материалы и методы»), с соответствующими гистограммами количества клеток в зависимости от глубины. Нейроны кальретинина и кальбиндина наиболее плотны в слое II с уменьшением более глубокой плотности, тогда как нейроны парвальбумина более равномерно распределены по слоям.
Визуально вызванная пиковая активность
Гипотеза CCM делает подробные прогнозы относительно пиковой активности (Дуглас и Мартин, 1991; Дуглас и др. , 1995). Во-первых, предполагается, что возбуждение с последующим подавлением является общей чертой CCM. Во-вторых, возбуждающие и тормозящие нейроны получают синхронизированные входные сигналы; в противном случае повторяющиеся возбуждающие связи привели бы к неограниченному возбуждению. В-третьих, внутриклеточные записи в первичной зрительной коре головного мозга показывают, что пирамидные клетки в поверхностных слоях достигают максимального состояния гиперполяризации позже, чем в глубоких слоях.Эти прогнозы были проверены в SEF путем измерения скорости визуально вызванных разрядов одиночных и множественных единиц, записанных одновременно с LFP. Из зарегистрированных одиночных единиц 103 (35%) продемонстрировали четкую модуляцию после предъявления мигающего стимула (63 обезьяны E, 40 обезьян X). Мы также зарегистрировали пороговую активность нескольких единиц с четкими визуально вызванными ответами в 58 точках (42 обезьяны E, 16 обезьян X). Мы включили эту многокомпонентную активность в наш анализ латентности, но исключили ее в анализах, где ширина спайка использовалась для классификации типов нейронов.
, 1995). Во-первых, предполагается, что возбуждение с последующим подавлением является общей чертой CCM. Во-вторых, возбуждающие и тормозящие нейроны получают синхронизированные входные сигналы; в противном случае повторяющиеся возбуждающие связи привели бы к неограниченному возбуждению. В-третьих, внутриклеточные записи в первичной зрительной коре головного мозга показывают, что пирамидные клетки в поверхностных слоях достигают максимального состояния гиперполяризации позже, чем в глубоких слоях.Эти прогнозы были проверены в SEF путем измерения скорости визуально вызванных разрядов одиночных и множественных единиц, записанных одновременно с LFP. Из зарегистрированных одиночных единиц 103 (35%) продемонстрировали четкую модуляцию после предъявления мигающего стимула (63 обезьяны E, 40 обезьян X). Мы также зарегистрировали пороговую активность нескольких единиц с четкими визуально вызванными ответами в 58 точках (42 обезьяны E, 16 обезьян X). Мы включили эту многокомпонентную активность в наш анализ латентности, но исключили ее в анализах, где ширина спайка использовалась для классификации типов нейронов.
Визуально чувствительные единицы были зарегистрированы во всех слоях SEF (рис. 10, таблица 1). Как сообщалось ранее (Schlag and Schlag-Rey, 1987; Schall, 1991; Chen and Wise, 1995), некоторые устройства показали усиление пиков (80, 50%), а другие – подавление пиков (81, 50%) после появления зрительного стимула. Мы выполнили 4 × 2 ANOVA, чтобы определить, различалась ли задержка этих ответов между слоями (четыре уровня) и типами ответа (два уровня, то есть усиление против подавления). Задержки были значительно короче для единиц с усиленными ответами, чем для единиц с подавленными ответами ( F (1,149) = 16.62, p = 7,6 × 10 −5 ). Таким образом, в совокупности пиковая активность SEF представляет собой визуально вызванную волну возбуждения, за которой следует подавление. Латентность визуально вызванных откликов единиц существенно не различалась по слою ( F (3149) = 1,18, p = 0,32), а также не было очевидного взаимодействия между слоем и типом ответа ( F (3,149) = 0,53, p = 0,66). Таким образом, временная и пространственная картина возбуждения и подавления в SEF согласуется с первым предсказанием модели CCM.
Таким образом, временная и пространственная картина возбуждения и подавления в SEF согласуется с первым предсказанием модели CCM.
Одиночные и многоэлементные задержки визуально вызванной реакции на мигающие стимулы. a , Репрезентативные единицы, записанные из слоя II (красный), слоя III (зеленый), слоя V (синий) и слоя VI (черный), демонстрирующие либо повышенную (слева), либо подавленную скорость разряда (справа) после стимула . Заштрихованные области показывают 95% доверительные интервалы, измеренные по предъявленным стимулам. Вертикальными линиями отмечены задержки ответа. b , Кумулятивные распределения задержек отклика модулей, разделенные по уровням и типам отклика.Расширенные ответы показали более короткие задержки, чем подавленные. См. Также таблицу 1.
Таблица 1.Сводная статистика единиц с визуальными реакциями, записанными с каждого слоя
Семьдесят семь единиц с широкими пиками и 15 единиц с узкими пиками показали четко определяемое время начала (среднее ± стандартное отклонение, 107 ± 43 мс единиц с широкими пиками, 97 ± 43 мс единиц с узкими пиками). Чтобы проверить, получают ли возбуждающие и тормозные нейроны синхронизированные входные сигналы, мы определили, обнаруживают ли предполагаемые пирамидные клетки и интернейроны в SEF схожие латентные периоды.В соответствии с моделью CCM, время начала не различается значимо между единицами с широким и узким пиками [сумма рангов Уилкоксона W = 3664, p = 0,38]. Чтобы гарантировать, что ширина шипов была надлежащим образом оценена по хорошо изолированным единичным единицам, мы повторили этот анализ после отбраковки единиц с ложноположительными показателями> 5%. Этот критерий исключил 14 предполагаемых пирамидных клеток и 4 предполагаемых интернейрона. Идентичные результаты были получены с этим подмножеством [сумма рангов Уилкоксона W = 2410, p = 0.48].
Чтобы проверить, получают ли возбуждающие и тормозные нейроны синхронизированные входные сигналы, мы определили, обнаруживают ли предполагаемые пирамидные клетки и интернейроны в SEF схожие латентные периоды.В соответствии с моделью CCM, время начала не различается значимо между единицами с широким и узким пиками [сумма рангов Уилкоксона W = 3664, p = 0,38]. Чтобы гарантировать, что ширина шипов была надлежащим образом оценена по хорошо изолированным единичным единицам, мы повторили этот анализ после отбраковки единиц с ложноположительными показателями> 5%. Этот критерий исключил 14 предполагаемых пирамидных клеток и 4 предполагаемых интернейрона. Идентичные результаты были получены с этим подмножеством [сумма рангов Уилкоксона W = 2410, p = 0.48].
Чтобы проверить, можно ли различить поверхностный и глубокий слои на основе времени гиперполяризации, мы определили, следовало ли максимальное визуально вызванное подавление спайков более длительному временному ходу в нейронах, записанных с верхних и нижних слоев в SEF.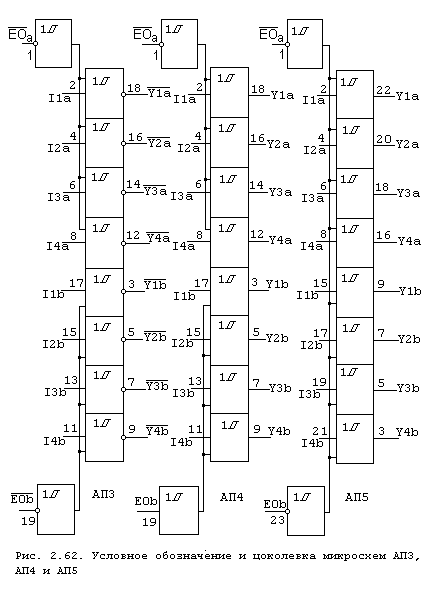 Это аналогично внутриклеточным записям, показывающим, что время максимальной гиперполяризации в нейронах поверхностного слоя происходит позже, чем в нейронах глубокого слоя первичной зрительной коры (Douglas and Martin, 1991). Кроме того, поскольку этот результат основан на ламинарных различиях в относительной пирамидной экспрессии нейронов рецепторов GABA A и GABA B , мы предсказали, что этот эффект будет ограничен только единицами с широким выбросом.Внеклеточные записи в SEF подтвердили эти прогнозы (рис. 11). Мы ограничили этот анализ визуально связанными единицами, которые показали снижение пиков после начала стимула по сравнению с исходным уровнем. Время максимального подавления спайков, зарегистрированное от единиц в слоях II и III (среднее ± SD, 144 ± 61 мс), было значительно позже, чем время максимального подавления спайков от единиц, зарегистрированных в слоях V и VI (среднее ± SD, 115 ± 62 мс) [сумма рангов Уилкоксона W = 2387, p = 4,30 × 10 −3 ].
Это аналогично внутриклеточным записям, показывающим, что время максимальной гиперполяризации в нейронах поверхностного слоя происходит позже, чем в нейронах глубокого слоя первичной зрительной коры (Douglas and Martin, 1991). Кроме того, поскольку этот результат основан на ламинарных различиях в относительной пирамидной экспрессии нейронов рецепторов GABA A и GABA B , мы предсказали, что этот эффект будет ограничен только единицами с широким выбросом.Внеклеточные записи в SEF подтвердили эти прогнозы (рис. 11). Мы ограничили этот анализ визуально связанными единицами, которые показали снижение пиков после начала стимула по сравнению с исходным уровнем. Время максимального подавления спайков, зарегистрированное от единиц в слоях II и III (среднее ± SD, 144 ± 61 мс), было значительно позже, чем время максимального подавления спайков от единиц, зарегистрированных в слоях V и VI (среднее ± SD, 115 ± 62 мс) [сумма рангов Уилкоксона W = 2387, p = 4,30 × 10 −3 ].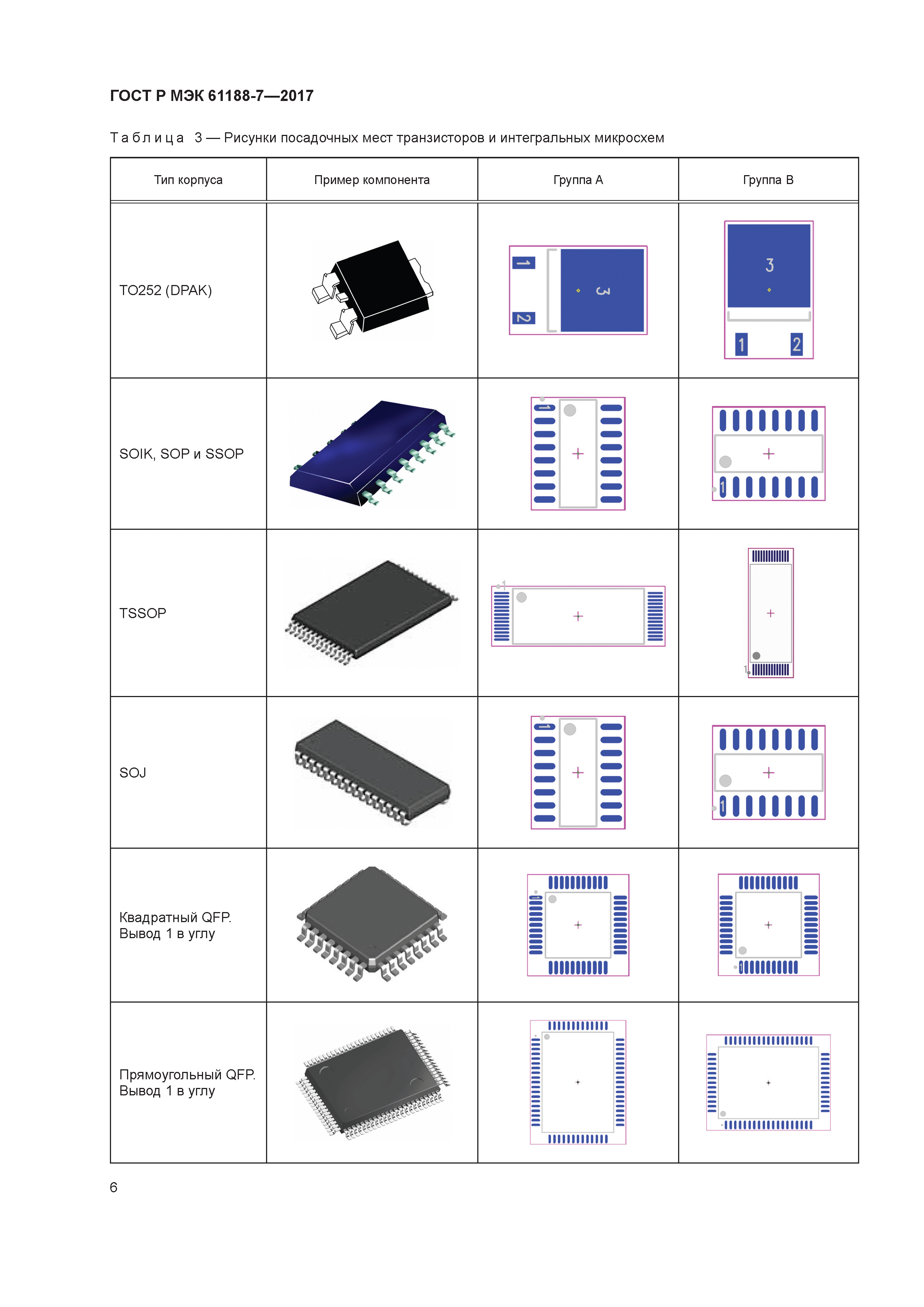 Мы дополнительно исследовали этот эффект, анализируя единицы с широкими и узкими пиками по отдельности. Как и предполагалось, нейроны с широкими шипами, зарегистрированные в поверхностных слоях, показали значительно более позднее максимальное подавление спайков, чем нейроны с широкими шипами, зарегистрированные в глубоких слоях [сумма рангов Уилкоксона W = 1719, p = 3,5 × 10 −3 ], но это не наблюдалось для единиц с узкими пиками [сумма рангов Уилкоксона W = 58, p = 0,80]. Мы повторили этот анализ, используя только хорошо изолированные единичные единицы, показывающие уровень ложноположительных результатов <5%.Этот критерий исключил 11 и 2 единицы широких игл из поверхностных и глубоких слоев, соответственно, и 3 единицы узких игл из поверхностных слоев. Идентичные результаты были получены с этим подмножеством. Единицы с широкими шипами в поверхностных слоях все еще демонстрировали более позднее максимальное подавление шипов [сумма рангов Уилкоксона W = 1042, p = 0,01], и этот результат остался отсутствовать для единиц с узкими шипами [сумма рангов Уилкоксона W = 28, p = 0,95].
Мы дополнительно исследовали этот эффект, анализируя единицы с широкими и узкими пиками по отдельности. Как и предполагалось, нейроны с широкими шипами, зарегистрированные в поверхностных слоях, показали значительно более позднее максимальное подавление спайков, чем нейроны с широкими шипами, зарегистрированные в глубоких слоях [сумма рангов Уилкоксона W = 1719, p = 3,5 × 10 −3 ], но это не наблюдалось для единиц с узкими пиками [сумма рангов Уилкоксона W = 58, p = 0,80]. Мы повторили этот анализ, используя только хорошо изолированные единичные единицы, показывающие уровень ложноположительных результатов <5%.Этот критерий исключил 11 и 2 единицы широких игл из поверхностных и глубоких слоев, соответственно, и 3 единицы узких игл из поверхностных слоев. Идентичные результаты были получены с этим подмножеством. Единицы с широкими шипами в поверхностных слоях все еще демонстрировали более позднее максимальное подавление шипов [сумма рангов Уилкоксона W = 1042, p = 0,01], и этот результат остался отсутствовать для единиц с узкими шипами [сумма рангов Уилкоксона W = 28, p = 0,95]. Добавляя доказательства различий в ингибировании между слоями, общая вероятность регистрации единиц с подавленными ответами была также выше в поверхностных слоях, о чем свидетельствует значительная разница в глубине по типу ответа [сумма рангов Уилкоксона W = 6509, p = 5 .28 × 10 −4 ]. Таким образом, временной ход и распределение подавления визуально вызванных спайков в SEF согласуется с моделью CCM.
Добавляя доказательства различий в ингибировании между слоями, общая вероятность регистрации единиц с подавленными ответами была также выше в поверхностных слоях, о чем свидетельствует значительная разница в глубине по типу ответа [сумма рангов Уилкоксона W = 6509, p = 5 .28 × 10 −4 ]. Таким образом, временной ход и распределение подавления визуально вызванных спайков в SEF согласуется с моделью CCM.
Латентность максимального подавления спайков, дифференцированная по глубине нейрона и ширине спайка. Поверхностные слои (II и III) в сравнении с глубокими слоями (V и VI) и единицы с широкими шипами (синим цветом) по сравнению с элементами с узкими шипами (красным цветом). Планки погрешностей указывают на SEM. Слева: пример блока, иллюстрирующий измерение задержки до максимального подавления пиков.На вставке вверху справа показаны узкие и широкие пики, составляющие образец. Статистически значимая разница наблюдалась в задержке до максимального подавления спайков между широкими единицами, зарегистрированными в поверхностных слоях, и широкими единицами, зарегистрированными в глубоких слоях. Никакие другие сравнения существенно не различались.
Никакие другие сравнения существенно не различались.
Пиковая активность, связанная с саккадом
единиц с модуляцией, связанной с саккадами, также были записаны со всех слоев SEF. Мы зарегистрировали 27 отдельных единиц (9%) с активностью, связанной со спонтанными саккадами, сделанными в темноте (14 обезьян E, 13 обезьян X).Десять из этих нейронов (37%) также проявляли визуально вызванную активность. Мы дополнительно записали связанную с саккадой активность нескольких единиц в 24 местах (12 обезьян E, 12 обезьян X). Семнадцать из этих многокомпонентных записей (71%) также демонстрировали визуально вызываемую активность. Из этих 51 единиц, связанных с саккадами, 23 (45%) показали пресаккадическую модуляцию и 28 (55%) показали постаккадические ответы. Кроме того, 26 единиц (51%) показали повышенную скорость стрельбы до и во время саккад, а остальные 25 (49%) показали подавление.Ни глубина (среднее значение = 0,38 мм относительно текущего стока 1, SD = 0,67 мм, сумма рангов Уилкоксона W = 17323, p = 0,64), ни ширина пиков (среднее значение = 350 мкс, SD = 99 мкс, ранг Вилкоксона сумма W = 5224, p = 0,44), ни коэффициенты вариации в интервалах между спайками (среднее значение CV = 1,42, SD CV = 0,49, сумма рангов Уилкоксона W = 6597, p = 0,39).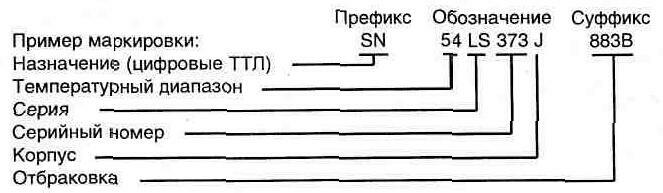 значительно между визуально связанными и связанными с саккадами единицами. ANOVA 4 × 2 показал, что задержки не различались по слоям ( F (3,43) = 1.12, p = 0,35), в отличие от нейронов, реагирующих на зрение, латентность усиленных и подавленных ответов существенно не различалась ( F (1,43) = 1,09, p = 0,30), равно как и взаимодействие между глубина и тип отклика ( F (3,43) = 2,04, p = 0,12). В таблице 2 представлены сводные статистические данные для этой нейронной популяции, разделенные слоями. В целом, эти результаты демонстрируют относительное отсутствие активности, связанной с саккадами, в SEF, когда саккады инициируются без видимых целей.
значительно между визуально связанными и связанными с саккадами единицами. ANOVA 4 × 2 показал, что задержки не различались по слоям ( F (3,43) = 1.12, p = 0,35), в отличие от нейронов, реагирующих на зрение, латентность усиленных и подавленных ответов существенно не различалась ( F (1,43) = 1,09, p = 0,30), равно как и взаимодействие между глубина и тип отклика ( F (3,43) = 2,04, p = 0,12). В таблице 2 представлены сводные статистические данные для этой нейронной популяции, разделенные слоями. В целом, эти результаты демонстрируют относительное отсутствие активности, связанной с саккадами, в SEF, когда саккады инициируются без видимых целей.
Сводная статистика единиц с саккадой, записанная с каждого слоя
Обсуждение
Мы сообщаем о CSD и ламинарной одно- и многокомпонентной активности, зарегистрированной в SEF обезьяны, агранулярной области в лобной доле. Многие результаты согласуются с CCM, сформулированным для объяснения гранулярных сенсорных областей. Однако некоторые аспекты данных несовместимы с СКК. В частности, двойные стоки тока появлялись почти одновременно в слоях III и V, а латентность визуального отклика не изменялась по слоям.
Многие результаты согласуются с CCM, сформулированным для объяснения гранулярных сенсорных областей. Однако некоторые аспекты данных несовместимы с СКК. В частности, двойные стоки тока появлялись почти одновременно в слоях III и V, а латентность визуального отклика не изменялась по слоям.
CCM в SEF?
Ламинарный паттерн CSD в агранулярной SEF во многом соответствует паттерну, предсказанному моделью CCM, разработанной на основе данных, собранных в зернистой первичной зрительной коре (рис. 1 a ; Gilbert, 1983; Callaway, 1998; Douglas and Martin, 2004 г.). После появления стимула спады тока в SEF начинаются в средних слоях и распространяются на поверхностные и глубокие слои. Однако, в отличие от сенсорных областей коры, в SEF после визуальной стимуляции были видны два начальных спада, соответствующие слоям III и V.Хотя этот результат ожидался на основе исследований анатомических индикаторов (рассмотренных ниже), он неожидан для классической модели CCM, в которой активность в одном входном слое предшествует активности в других слоях.
В дополнение к активности с прямой связью, модель CCM также описывает, как локальное рекуррентное возбуждение усиливает восходящий вход, в то время как ингибирование предотвращает неконтролируемое возбуждение (рис. 1 b ). Восходящий вход возбуждает как пирамидные клетки, так и интернейроны, что приводит к характерному паттерну возбуждения с последующим подавлением (Douglas and Martin, 1991; Douglas et al., 1995; см. также Brunel and Wang, 2001; Chance et al., 2002; Haider et al., 2006). В соответствии с этим мы обнаружили повышенную скорость разряда, предшествующую подавлению, на ~ 30 мс во всех слоях SEF. Мы также обнаружили, что предполагаемые пирамидные нейроны (единицы с широкими пиками) и предполагаемые интернейроны (единицы с узкими пиками) с одинаковой вероятностью демонстрируют начальное усиление пиков после зрительной стимуляции, и что латентность начала не различалась между этими популяциями.
Внутриклеточные записи подчеркивают различия во времени максимальной гиперполяризации между поверхностными и глубокими пирамидными клетками, опосредованными рецепторами GABA A и GABA B (Douglas and Martin, 1991). В соответствии с этими результатами мы обнаружили, что единицы, зарегистрированные в поверхностных слоях, с большей вероятностью реагировали на визуальные стимулы подавлением, и мы отметили более длительный временной ход для максимального подавления скорости разряда в поверхностных слоях по сравнению с записью в глубоких слоях. Этот эффект был ограничен предполагаемыми пирамидными клетками, что позволяет предположить, что ламинарные различия в экспрессии GABA A по сравнению с GABA B только в этом типе клеток могут объяснить результаты. Чтобы избежать путаницы, мы отмечаем, что наши меры аналогичны, но не идентичны тем, о которых сообщалось ранее.Хотя Дуглас и Мартин (1991) измерили время максимальной гиперполяризации внутриклеточных потенциалов в первичной зрительной коре анестезированных кошек после электростимуляции таламических афферентов, мы зарегистрировали подавление скорости разряда от внеклеточных потенциалов в SEF бодрствующих обезьян при представлении визуальной стимуляции.
В соответствии с этими результатами мы обнаружили, что единицы, зарегистрированные в поверхностных слоях, с большей вероятностью реагировали на визуальные стимулы подавлением, и мы отметили более длительный временной ход для максимального подавления скорости разряда в поверхностных слоях по сравнению с записью в глубоких слоях. Этот эффект был ограничен предполагаемыми пирамидными клетками, что позволяет предположить, что ламинарные различия в экспрессии GABA A по сравнению с GABA B только в этом типе клеток могут объяснить результаты. Чтобы избежать путаницы, мы отмечаем, что наши меры аналогичны, но не идентичны тем, о которых сообщалось ранее.Хотя Дуглас и Мартин (1991) измерили время максимальной гиперполяризации внутриклеточных потенциалов в первичной зрительной коре анестезированных кошек после электростимуляции таламических афферентов, мы зарегистрировали подавление скорости разряда от внеклеточных потенциалов в SEF бодрствующих обезьян при представлении визуальной стимуляции. Учитывая эти различия в технике, видах и ареалах, сходство результатов исследований позволяет предположить, что это может быть общей особенностью неокортекса млекопитающих.
Учитывая эти различия в технике, видах и ареалах, сходство результатов исследований позволяет предположить, что это может быть общей особенностью неокортекса млекопитающих.
Отношение к предыдущим анатомическим исследованиям
Быстрый визуальный ввод может передаваться в SEF через медиодорсальное ядро (MD) таламуса (Huerta, Kaas, 1990; Shook et al., 1990), которое иннервируется верхним бугристым бугорком (Benevento and Fallon, 1975; Harting et al. ., 1980). Но проекции MD не могут объяснить начальный сток тока, который мы наблюдали в слое V, потому что афференты заканчиваются в нижнем слое III SEF (Giguere and Goldman-Rakic, 1988). Зрительные афференты к SEF также поставляются кортикальными областями, включая латеральную интрапериетальную область, область 7a, FEF, верхнюю височную полисенсорную область, визуальную область 6a (V6a) и медиальную верхнюю височную область (MST; Barbas and Pandya, 1987). ; Huerta and Kaas, 1990; Shipp et al., 1998). Такие области, как MST и V6a, вероятно, обеспечивают быстрый визуальный ввод в SEF (Schmolesky et al., 1998). В соответствии с нашими результатами, инъекции ортоградных индикаторов в MST (Maioli et al., 1998) и в V6a (Shipp et al., 1998) выявляют терминалы в слоях III и V в SEF. Кроме того, в соответствии с нашим наблюдением, что слой VI является последним слоем, показывающим визуально связанную CSD, проекции из областей спинного потока только редко заканчиваются в слое VI SEF (Maioli et al., 1998; Shipp et al., 1998). Таким образом, ламинарное распределение стоков тока в ответ на зрительную стимуляцию хорошо согласуется с известной анатомией.
Изучение CCM может выявить корковую иерархию в агранулярной коре во многом так же, как и в зернистой коре (Felleman and Van Essen, 1991). Shipp (2005) предположил, что отношение проекций слоя III к уровню V можно использовать для размещения агранулярных областей в ареальной иерархии. Наше открытие относительно короткой задержки, управляющей входными данными на уровни III и V SEF, согласуется с этой гипотезой. Эти опускания тока в SEF имели одинаковую задержку и величину после визуальной стимуляции, что позволяет предположить, что оба слоя получают визуальные афференты одинаковой силы.
Связь с предыдущими исследованиями внутричерепных записей
Широкопольные импульсные стимулы, такие как используемые здесь, использовались для сравнения профилей ламинарной активации на последовательных стадиях визуальной иерархии (Schroeder et al., 1989; Givre et al., 1994; Schroeder et al., 1998). Средняя латентность визуально вызванного CSD в SEF (51 мс) согласуется с работой Schroeder et al. (1998), которые измерили CSD по всему зрительному пути и сообщили о самых длительных латентных периодах в нижневисочной коре (49 мс).Наш показатель задержки CSD также сравним с таковыми для визуально вызванных LFP в SEF во время задачи визуального поиска (Purcell et al., 2012). Кроме того, наблюдаемые здесь задержки одно- и многочастичных ответов согласуются с несколькими предыдущими сообщениями (Schall, 1991; Chen and Wise, 1995; Pouget et al., 2005; Purcell et al., 2012).
В областях раннего зрения латентные периоды ответа самые короткие в слое IV и больше в супрагранулярных и инфрагранулярных слоях (Bullier and Henry, 1980; Maunsell and Gibson, 1992; Raiguel et al., 1999). При записи в первичной зрительной коре несколько групп сообщили, что латентные периоды в супрагранулярных слоях больше, чем наблюдаемые в глубоких слоях (Maunsell and Gibson, 1992; Self et al., 2013). В SEF визуальная задержка одного и нескольких единиц не различалась по слоям. Этот результат неожиданный для модели CCM. Возможно, использованные нами стимулы сыграли роль в создании этого загадочного результата. Хотя использованная нами парадигма широкоугольной вспышки является необходимым шагом для сравнения CSD с опубликованными отчетами из ранних визуальных областей, она несколько менее эффективна для управления потенциалами действия в одиночных и множественных единицах в SEF (рис.10 а ). Поскольку изменения в скорости стрельбы были скромными, наши оценки задержки могли иметь повышенную изменчивость. Однако наблюдаемая нами изменчивость была сопоставима с вариабельностью, о которой сообщалось в исследовании, в котором использовались локализованные, соответствующие поведению визуальные стимулы (Pouget et al., 2005). Авторы этого исследования обнаружили разницу в латентности 37 мс в SEF. Однако гранулярный и супрагранулярный слои в первичной зрительной коре обнаруживают различия в латентности только ~ 10-15 мс (Maunsell and Gibson, 1992), поэтому различия в латентности между слоями может быть труднее обнаружить при SEF.Таким образом, этот нулевой результат не следует интерпретировать как окончательный.
Подавление отклика было обычным явлением в наших нейронных записях. Несколько исследователей отметили подавление нейронной активности при SEF (Schlag and Schlag-Rey, 1987; Schall, 1991; Chen and Wise, 1995). Доля единиц, отображающих подавление, варьируется в разных исследованиях, но это может зависеть от параметров стимула и непредвиденных обстоятельств задачи. Наблюдаемое нами подавление может также возникать из-за окружения рецептивного поля. Архитектура рецептивного поля центр / окружение не была охарактеризована в SEF, но об этом сообщалось в FEF (Schall et al., 2004; Cavanaugh et al., 2012).
Наши измерения ширины спайков и изменчивости SEF согласуются с измерениями из других визуальных областей с использованием аналогичных показателей (Mitchell et al., 2007; Cohen et al., 2009). Наше соотношение единиц с узкими и широкими пиками (79%) сравнимо с тем, что указано в V4 (73%; Mitchell et al., 2007). Кроме того, единицы с узкими пиками в SEF обнаруживают повышенную вариабельность времени пика, обеспечивая новые доказательства того, что эти клетки представляют собой интернейроны.
Резюме и выводы
Эти данные подчеркивают обширную мультимодальную информацию, которую можно получить, поскольку достижения в технологии электродных матриц позволяют исследователям регистрировать хорошо изолированные единичные единицы с LFP через корковые слои.Мы обнаружили, что некоторые особенности визуально вызванных ламинарных ответов в агранулярных SEF соответствуют тем, которые наблюдаются в зернистых сенсорных областях. Крайне важно, что в средних слоях SEF были очевидны выраженные спады тока. Глубина этих погружений обеспечивает маркер, по которому можно измерить глубину каждого зарегистрированного нейрона in vivo . Этот прогресс, несомненно, окажется решающим в описании нейронных взаимодействий внутри целых, ведущих себя обезьян.
APDN продлил контракт с U.Агентство логистики S. Defense (DLA)
Для маркировки ДНК с помощью SigNature® DNAИнформация о выпуске
4 ноября, 2015
Applied DNA Sciences, Inc. объявила, что Агентство оборонной логистики США (DLA) предоставило компании продление на один год ее текущего контракта до 12 ноября 2016 года. Согласно условиям текущего контракта, Applied DNA предоставляет DLA SigNature. ® Знаки ДНК, соответствующее оборудование, услуги и обучение в Центре тестирования электронных продуктов DLA в Колумбусе, штат Огайо, с упором на микросхемы для поддержки существующих систем.Доктор Джеймс Хейворд, президент и главный исполнительный директор компании Applied DNA Sciences, заявил: «Сектор обороны правительства США представляет существенные возможности для прикладных наук о ДНК и наших клиентов, которые в настоящее время входят в пять различных федеральных агентств и являются важными первыми пользователями нашей технологии. Наша технология на основе ДНК предлагает DLA средство, с помощью которого можно обеспечить надежность и безопасность цепочки поставок ».
Applied DNA продолжает выполнение дополнительных федеральных контрактов:
- Фаза II Исследования инноваций малого бизнеса (SBIR) контракт с U.Агентство противоракетной обороны получило награду в июле 2014 года за разработку более эффективных методов применения ДНК SigNature в больших объемах, охват до 100 производителей оригинальных компонентов и разработку оптического считывающего устройства для быстрого обнаружения чернил ДНК SigNature, нанесенных на компонент в полевых условиях. .
- Фонд быстрых инноваций (RIF), заключенный с канцелярией министра обороны , заключен в августе 2014 года на разработку единой платформы аутентификации – ДНК SigNature и дополнительных технологий Applied DNA – для идентификации подлинных продуктов и предотвращения проникновения подделок в шести странах. Министерство обороны (DOD) Группы федерального снабжения (FSG).Эти FSG включают 66 классов товаров, включая компоненты электронного и автомобильного оборудования, подшипники, аксессуары для двигателей, трубы, трубки, шланги и фитинги, оборудование и абразивные материалы. Согласно этому контракту, Applied DNA Sciences также будет выполнять такие услуги, как разработка, тестирование и оценка, полевые испытания и переход к государственным операциям.
- Оба контракта являются единовременными. Однако они представляют возможности для получения дохода в будущем
- Два не связанных с обороной федеральных агентства поддерживают отношения с Applied DNA .Объем услуг, которые предоставляет компания, и финансовые условия являются конфиденциальными.
- Компания изучает дополнительные отношения с правительством и возможные будущие контракты.
Дженис Мераглиа, вице-президент по правительственным и военным программам компании Applied DNA Sciences, прокомментировала: «Три контракта Министерства обороны США впечатляют сами по себе и в совокупности подчеркивают присущую нашей платформе гибкость и ее уникальную функциональность в масштабе, в котором нуждаются государственные учреждения. .Текущий контракт DLA ориентирован на обслуживание и ремонт компонентов, уже изготовленных для поддержки существующих систем в театре; SBIR больше ориентирован на новые производимые детали, а RIF призывает к единой платформе аутентификации для широкого спектра товаров. Вместе они иллюстрируют широту и глубину применения наших технологий для наших оборонных и федеральных заказчиков ».
О прикладных науках о ДНК
Прикладные науки о ДНК делают жизнь реальной и безопасной, предоставляя решения на основе биотехнологий, которые помогают защитить продукты, бренды, целые цепочки поставок и интеллектуальную собственность компаний, правительств и потребителей от краж, подделок, мошенничества и утечки.Запатентованные растворы ботанической ДНК могут использоваться для идентификации, маркировки, отслеживания и отслеживания продуктов, чтобы помочь гарантировать подлинность, отслеживаемость и качество продуктов. SigNature DNA является сердцем семейства некопируемых решений для обеспечения безопасности и аутентификации, таких как SigNature® T и fiberTyping®, предназначенных для текстильных изделий и одежды, DNAnet® для защиты от краж и предотвращения потерь и digitalDNA®, обеспечивающих эффективное отслеживание и след. Все они обеспечивают судебно-медицинскую цепочку доказательств и могут использоваться для судебного преследования преступников.
Посетите adnas.com, чтобы узнать больше, узнать о мероприятиях и узнать больше о том, как прикладные науки о ДНК делают жизнь реальной и безопасной. Обыкновенные акции котируются на NASDAQ под символом APDN, а варранты – под символом APDNW.
Заявления о перспективах
Заявления, сделанные APDN в этом пресс-релизе, могут иметь «прогнозный» характер по смыслу Закона о судебных разбирательствах по частным ценным бумагам 1995 года. Заявления прогнозного характера описывают будущие планы, прогнозы, стратегии и ожидания APDN и основаны на предположений и связаны с рядом рисков и неопределенностей, многие из которых находятся вне контроля APDN.Фактические результаты могут существенно отличаться от прогнозируемых из-за нашей короткой операционной истории, ограниченных финансовых ресурсов, ограниченного признания рынка, рыночной конкуренции и различных других факторов, которые время от времени подробно описываются в отчетах и документах APDN SEC, включая наш Годовой отчет по форме 10-K подана 15 декабря 2014 г. с поправками от 6 марта 2015 г., а также наши последующие квартальные отчеты по форме 10-Q, поданные 9 февраля 2015 г., 11 мая 2015 г. и 10 августа 2015 г., которые доступны на сайте www.sec. губ.APDN не берет на себя никаких обязательств по публичному обновлению каких-либо прогнозных заявлений с целью отражения новой информации, событий или обстоятельств после указанной даты, чтобы отразить наступление непредвиденных событий, если иное не требуется по закону.
Варианты подключения, которые позволяют и ограничивают нейронные вычисления в сенсорной микросхеме
Ниже мы прилагаем полные обзоры всех трех рецензентов. Мы надеемся, что вы сможете ответить на каждую критику в своей редакции. Мы хотели бы прояснить один пункт, касающийся «размера синапсов» (первая критика рецензента 3).После дальнейшего обсуждения рецензентами рецензент 3 сделал следующие дополнительные комментарии: «Взаимосвязь между размером синапса, а также количеством синапсов и силой соединения является частью общей предпосылки исследований дрозофилы. большинство синапсов имеют одинаковый размер, так что сила пути действительно приближается к числу синапсов без ссылки на их размер, но даже в этом случае размер синапса редко сообщается и сравнивается между областями нейропиля.Чтобы продемонстрировать модульность размера синапса, необходимо было бы измерить только ~ 40 синаптических профилей в правильном поперечном сечении и 20 в продольном сечении, чтобы определить вариации в размерах контактов синапсов и сравнить с другими опубликованными отчетами. Это не только затронет важный компонент силы проводящих путей, но и признает важную проблему, которая может быть неочевидной для читателей, работающих с различными нервными системами ».
Мы согласны с тем, что размер синапсов является важным вопросом.Здесь есть два вопроса, на которые мы можем ответить с помощью нашего набора данных. Во-первых, насколько различается размер синапсов? Во-вторых, как вариации размера синапсов коррелируют с другими анатомическими вариациями?
Чтобы ответить на эти вопросы, мы случайным образом выбрали 10 ORN (5 слева и 5 от правой антенны) и вручную сегментировали все их синапсы на все 5 наших реконструированных PN (n T-образных стержней = 683, n профили пн = 1106). В каждом синапсе были измерены две характеристики: пресинаптический объем Т-образного стержня ORN и площадь постсинаптического контакта PN (PSCA) для всех присутствующих постсинаптических профилей PN.Чтобы повысить точность этих измерений, каждый объем Т-образного стержня и PSCA были независимо измерены тремя разными аннотаторами, а затем измерения разных аннотаторов были объединены после взвешивания измерений каждого аннотатора в соответствии с их общей погрешностью измерения по всему набору данных (относительная к другим аннотаторам).
Результаты нашего анализа размера синапсов теперь показаны в новом дополнении (Рисунок 1 – Приложение 6). В целом, мы обнаружили, что два показателя размера синапсов (объем Т-образного стержня и площадь постсинаптического контакта) были значительно (хотя и слабо) коррелированы.Они также значительно коррелировали с количеством постсинаптических профилей на Т-образный стержень. Это означает, что должны существовать механизмы, которые производят скоординированные изменения всех трех переменных на основе синапса за синапсом (объем Т-образного стержня, площадь постсинаптического контакта и количество постсинаптических профилей).
Тем не менее, когда мы сравнивали разные соединения ORN → PN (путем усреднения размера всех синапсов, принадлежащих одному и тому же соединению), мы обнаружили, что размер синапса был лишь незначительным источником вариации между соединениями (рисунок 1 – приложение к рисунку 6) .Основным источником различий между соединениями по-прежнему было количество синапсов на соединение (рис. 5). Другими словами, то, что отличает одно соединение от другого, – это прежде всего количество синапсов, которые оно содержит, а не размер этих синапсов.
Рецензент № 1:
Это отличная статья, объединяющая новые данные ЭМ-реконструкции с моделированием и предыдущими электрофизиологическими исследованиями, чтобы дать общее представление о взаимосвязи между структурой, физиологическими свойствами и функциями микросхем.Авт. Сосредотачиваются на реконструкции нейронов обонятельных рецепторов (ORNs) и проекционных нейронов (PNs), которые иннервируют одну из пятидесяти пар клубочков в антеннальной доле Drosophila. Помимо получения ценных количественных ультраструктурных данных о синаптической связности, авторы создали компартментальную модель, которая имитирует, как входы ORN интегрируются в PN, и использовали ее для анализа трех интересных аспектов своих данных: как PN компенсируют разницу в их количестве. (особенно интересно), почему ипси-латеральные ORN обнаруживают более сильные связи с PN, чем контрлатеральные ORN, и каково функциональное значение вариаций в количестве синапсов между отдельными парами ORN → PN.Авторы также явно обсудили ограничения своего исследования (например, их модель не принимает во внимание тормозящий вход или вариации силы отдельных синапсов). Среди недавнего всплеска исследований по электромагнитной реконструкции этот считается одним из лучших для интеграции структуры и физиологических свойств хорошо изученной модельной схемы и как таковой должен представлять широкий интерес для сообщества нейробиологов.
Благодарим за положительный отзыв.
Мы отмечаем, что теперь мы явно изучаем анатомические параметры, которые могут указывать на «сильные стороны отдельных синапсов».Чтобы быть точным, мы описываем вариации в параметрах размера синапса (объем Т-образного стержня, площадь постсинаптического контакта). Мы обнаружили, что, хотя эти параметры существенно различаются в разных синапсах, они очень незначительно различаются в разных синаптических связях, как отмечалось выше.
Рецензент № 3:
Ссылки даются на текстовые страницы, а не на pdf.
Варианты подключения, которые позволяют и ограничивают нейронные вычисления в сенсорной микросхеме.
Прежде всего, я снимаю шляпу перед авторами за их самоотверженное исследование схем AL с использованием таких трудоемких методов.Для этого представления анатомия является основой, а аспекты моделирования более предположительны. В частности, я бы сказал, что самое слабое звено – это основная сюжетная линия, связь между синаптической структурой и синаптической силой. Даже в этом случае должна быть возможность связать все параметры коннектома с параметрами моделирования при условии, что у нас есть все необходимые меры. С этой целью авт. Предоставляют важные данные о размере дендритов, которые, как правило, не рассматривались широко в литературе по нейронам дрозофилы.Хотя подсчитать количество синапсов и графики распределения легко, однако в рассказах авторов пока отсутствует размер синапсов. Различается ли это каким-либо согласованным образом между (пре) синаптическими сайтами, и изменяются ли постсинаптические плотности, которые, как я полагаю, также видимы, с размером пресинаптического контакта? Размер дендрита может зависеть от них, но сам по себе не является показателем размера синапса. Если авторы заявляют о взаимосвязи между количеством синапсов и силой синаптической передачи, я думаю, что эти дополнительные параметры должны быть включены, и надеюсь, что их можно относительно легко добыть из существующих данных изображения, даже если я предполагаю, что это все фиксированные модули. .
По просьбе рецензента мы теперь измерили параметры размера синапсов. Мы предоставляем обзор нашего анализа и наших выводов в начале этого ответного документа, где мы отвечаем на Резюме редактора (в котором выделен этот конкретный запрос). Анализ размера синапсов теперь включен в рисунок 1 – приложение к рисунку 6.
Введение, четвертый абзац. Дополнительное важное обсуждение точности разводки в мозге можно найти в Takemura et al. (PNAS 112 (Rall, 1964): 13711-13716, 2015), на которые авторы ссылаются, но только позже в подтверждение отсутствия клеток в столбцах продолговатого мозга.Статья PNAS включает ссылки на сравнения между левым / правым гомологами у C. elegans и сравнения между числами синапсов в соседних столбцах мозгового вещества у Drosophila, оба из которых имеют отношение к данным авторов.
Выводы авторов относительно систематических и несистематических вариаций проводников (Введение, последний абзац и далее) отличаются от выводов, полученных из зрительной системы мух (Takemura et al., 2015). Фактически, анализ мозгового вещества указывает на высокий уровень синаптической и проводной точности, с которой дрозофила также способна строить синаптические цепи в своем мозгу, в данном случае в зрительной системе.Точность или инвариантность в последнем случае, по-видимому, обусловлена необходимостью надежной передачи на высоких временных частотах, требуемых зрением, – требование, которое может быть менее требовательным для обонятельного пути.
Мы согласны с тем, что Takemura et al. (2015) следует цитировать более заметно. Мы посвятили этой статье место в Обсуждении, но не во Введении. Теперь мы процитируем эту статью во Введении как пример изменчивости нейронных связей.
Рецензент предполагает, что требования визуальной обработки требуют большей точности нейронной архитектуры, чем требования обонятельной обработки. Мы не думаем, что относительные требования к визуальным и обонятельным вычислениям полностью ясны на этом этапе, и мы не согласны с идеей, что обонятельная обработка нетребовательна. Запахи могут колебаться с высокой временной частотой в естественных турбулентных условиях, с продолжительностью попадания шлейфа менее 20 мс. Эти временные функции полезны для навигации.Обонятельная трансдукция у Drosophila достаточно быстрая, чтобы отслеживать колебания до 10 Гц. Таким образом, обоняние не намного медленнее, чем зрение. Обоняние также имеет очень большие размеры, потому что для описания различий между различными химическими веществами необходимо большое количество физиолохимических измерений. Это еще одна причина, по которой обонятельная система может приспособиться к высокой скорости передачи информации на периферии. Поэтому особенно интересно то, что обонятельная система проводки должна быть неточной.
Мы не думаем, что уровень вариации, который мы здесь описываем, заметно больше, чем неточность, описанная Takemura et al. (2015). Takemura et al. описал два вида вариации:
1) Вариация двоичного соединения (отсутствующие соединения или соединения, которые обычно отсутствуют, но иногда присутствуют). В зрительной доле Takemura et al. обнаружили, что скорость двоичной вариации составляет около 1%. В синапсах ORN-PN мы обнаружили, что частота составляет около 2%. (Один ORN слева не смог установить соединение с двумя правыми PN, а один ORN справа не смог установить соединение с тремя левыми PN.Таким образом, частота пропущенных соединений составляет 2 + 3, разделенные на ((26 ORN + 27 ORN) × 5 PN), или 2%. Таким образом, вариация двоичного соединения не сильно отличается в двух наборах данных.
2) Вариация аналогового соединения (изменение количества синапсов на одно соединение во всех экземплярах определенного типа соединения). В зрительной доле Takemura et al. обнаружили, что CV числа синапсов на одно соединение колеблется от 0,08 до 0,87, в зависимости от типа соединения. В синапсах ORN-PN мы обнаруживаем, что CV равен 0.31. Таким образом, вариации аналогового подключения не сильно отличаются в двух наборах данных.
Учитывая изменчивость, описанную Takemura et al., Неудивительно, что мы также находим изменчивость в нашей реконструкции. Вклад нашего исследования не просто в описание изменчивости как таковой. Скорее, вклад этого исследования состоит в том, чтобы предоставить доказательства как систематических, так и несистематических видов анатомической изменчивости. Нам не нужно предполагать, что вся изменчивость – это «шум» (или «ошибка»).Мы показываем, что некоторые анатомические параметры изменяются систематическим образом, который должен быть адаптивным (учитывая структуру нашей модели), тогда как другие параметры изменяются бессистемным образом, что должно быть неадаптивным (опять же, учитывая структуру нашей модели). .
Прямая связь. Я нахожу «прямую связь» в следующей конструкции ненужной и сбивающей с толку: «вместе со всеми возбуждающими синапсами прямой связи на эти клетки». (Введение, четвертый абзац). «Прямая связь» и «вперед» кажутся мне противоречивыми, поскольку прямая связь может относиться к синаптическим выходам рассматриваемой клетки.Я думаю, что авторы хотят сказать «вместе со всеми возбуждающими входами (синапсами) в эти клетки».
Мы благодарим рецензента за указание на это. Нашей целью было просто отличить синапсы ORN → PN от других типов возбуждающих синапсов, которые не являются «прямой связью» (например, PN → PN синапсы, PN → ORN синапсы). В отредактированной рукописи эту фразу изменили, чтобы она читалась следующим образом: «вместе со всеми возбуждающими синапсами прямой связи, которые получают эти клетки».
«Три эксперта независимо идентифицировали клубочки DM6 в наборе данных ЭМ путем визуального осмотра на основе опубликованных карт уровня освещенности».Отчет примечателен больше тем, что не говорит, чем тем, что он говорит. Я считаю, что документации о том, как авторы выбрали нейроны DM6 для реконструкции, недостаточно. Хотя конфокальные данные ясно показывают клубочковую карту антеннальной доли, особенно на основе синаптической и генетической маркировки, такие данные вводят в заблуждение для электромагнитных исследований, которые выявляют все профили, а не их селективные особенности. В частности, нет четких глиальных границ между клубочками и клубочками, о которых авторы не упоминают, и которые могут быть однозначно идентифицированы только со ссылкой на карту окружающих клубочков или, возможно, с помощью декартовых координат, и я думаю, что авторы должны это признать. .Вполне возможно, что авторы приобрели достаточный опыт из физиологических записей, чтобы идентифицировать специфические клубочки, но это находится в интактной живой доле усиков. Клубочки также частично перекрывают друг друга в глубине нейропиле доли. Более того, нам не говорят, почему сам DM6 был выбран по сравнению с другими клубочками, и в качестве обоснования авторы могут захотеть процитировать свои собственные физиологические исследования. Эти вопросы являются центральными для анализа авторов и должны быть разъяснены в тексте, потому что, хотя я уверен, что были предприняты большие усилия, и мы можем быть уверены, что три эксперта сделали одну и ту же идентификацию, на самом деле это нетривиальная проблема. все на уровне EM.Большинство читателей не знают или не понимают этого, потому что конфокальные карты настолько убедительны, потому что большинство читателей никогда не видят ЭМ и потому что большинство может ошибочно предположить, что существует четкая граница глии, окружающая каждый клубочек. Также вполне возможно нацелить DM6 на живой препарат, что неоднократно проводилось в исследованиях лаборатории Уилсона, но контуры поверхности антенны также отличаются от 2D-изображений из ЭМ-срезов. Эти вопросы «почему» и «как» необходимо четко решить.Текст Методики должен позволять другому работнику повторить наблюдения авторов. В частности, как авторы могут предоставить читателю четкую уверенность в том, что они на самом деле нацелены на одни и те же клубочки на левом и правом полушариях мозга, и что левая / правая асимметрия, которую они обнаруживают, не являются результатом выбора разных клубочков на этих двух? стороны. Я полностью согласен, напротив, с тем, что авторы скрупулезно идентифицируют в наборе данных EM ORN и PN из их соответствующих ветвей и траекторий аксонов.
Границы клубочков антенных долей очерчены специальным классом глии (называемыми «обволакивающими глиями»; Doherty et al. 2009 J Neurosci29: 4768-81). Мы согласны с рецензентом в том, что эти глиальные оболочки могут быть более или менее ясными в материале ssEM, в зависимости от того, как был подготовлен образец и отображен, а также в зависимости от идентичности рассматриваемых клубочков. В нашем материале границы многих клубочков достаточно четкие при низком разрешении (см. Видео 1-2).Мы решили сосредоточиться на клубочке DM6, отчасти потому, что это выступающий клубок, с легко определяемым положением относительно клубочков других долей усиков. На опубликованных картах антеннальной доли DM6 имеет особенно последовательное расположение (Laissue et al., 1999 J Comp Neurol405, 543-552; Couto et al. 2005 Curr Biol15, 1535-1547; Grabe et al.2015 J Comp Neurol523, 530 -544). Мы также выбрали DM6, потому что есть много физиологической информации об этом клубочке. Наконец, DM6 был расположен в области серии EM, где имеется относительно мало недостающих секций (подробно описано в исходных материалах и методах).Мы пересмотрели методы, чтобы более четко объяснить наше обоснование выбора DM6. Тот факт, что мы правильно идентифицировали пару гомологичных клубочков на двух сторонах мозга, был в конечном итоге подтвержден, когда была проведена реконструкция: мы обнаружили, что аксоны ORN DM6 пересекали переднюю комиссуру, чтобы нацеливаться на другой клубок, который мы идентифицировали как DM6.
Рисунок 1A. Связанный: хотя и полезен как обзор EM, рисунок 1A не дает достаточной информации для идентификации DM6, особенно по оси z.Был ли реконструирован весь клубочек и как авторы решили, где лежат его границы?
Благодарим рецензента за возможность прояснить этот момент. Некоторые границы СД6 (а именно передняя, медиальная и дорсальная границы) были четкими при визуальном осмотре (см. Видео 1-2). Задняя, латеральная и вентральная границы были менее четкими и, таким образом, в конечном итоге определялись дендритами реконструированных PN. Все дендриты реконструированных нами PN заполнены полностью одинаковым объемом (что типично для большинства PN и большинства клубочков).Как описано в Методах, мы реконструировали каждый профиль пресинаптического профиля для каждого дендрита PN (по крайней мере, до границы DM6), и поэтому мы с высокой степенью уверенности идентифицировали каждый ORN DM6. Цель рисунка 1A – предоставить обзор ЭМ-разреза на уровне DM6, но идентификация DM6, конечно же, была сделана с помощью 3D-данных с высоким разрешением. Мы ожидаем, что специализированные читатели, заинтересованные в этом выпуске, изучат набор трехмерных данных на сайте neurodata.io.
Сироты: К чему относится процент сирот? Количество независимых фрагментов, которые были пресинаптическими по отношению к PN, но не могли быть связаны с идентифицированной клеткой? Сколько синапсов они внесли? Были ли осиротевшие с обоих концов, не имевшие контакта ни с идентифицированными ORN, ни с PN? Если да, то в какой пропорции?
Мы определяем сирот как фрагменты реконструированных нейритов, которые не могут быть связаны ни с одним нейроном.Мы изменили текст, чтобы уточнить это определение. Автор обзора прав, что сироты были независимыми фрагментами, которые были пресинаптическими по отношению к PN, но не могли быть связаны с идентифицированной клеткой. С нашей стратегией реконструкции (реконструировать все входы в PN) один конец всегда привязан к входному синапсу PN, поэтому мы не генерируем фрагменты, потерянные на обоих концах. Следовательно, по конструкции количество синапсов, внесенных сиротами в PNs, по существу является количеством самих сиротских фрагментов.
Утверждение (подраздел «Клетки, составляющие клубочковую микросхему», последний абзац) о том, что ложные продолжения между фрагментами легко обнаруживаются и исправляются во время независимого обзора, скорее всего, является атрибутом ветви, чем стратегии реконструкции, поскольку личинка нейроны у Drosophila, как правило, имеют относительно более прочные нейриты с дистальными конечностями, несущими большинство синапсов. Таким образом, вывод о том, что они очень редки при окончательной реконструкции, может быть менее обоснованным, чем утверждается.
Хотя ложные продолжения возможны, наша стратегия реконструкции способствует минимизации ложных продолжений с риском ложных завершений, чтобы максимизировать уверенность в восстанавливаемых нами соединениях. Аннотаторы перестают отслеживать
, если продолжение становится неоднозначным. Более того, все реконструированные нейроны были просмотрены несколькими независимыми аннотаторами.
Пресинаптические тройники. Прокомментируйте, пожалуйста, диапазон структурных фенотипов, демонстрируемых пресинаптическими Т-образными полосками на Рисунке 1 – приложение к рисунку 3, или из анализа авторов синапсов в последовательных срезах.
Мы включили количественный анализ объемов Т-образных стержней в исправленную рукопись (рис. 1 – приложение к рисунку 6), как описано выше. Мы считаем, что подробное описание форм Т-образных стержней выходит за рамки данного исследования, но мы планируем изучить этот вопрос в будущем.
Таблица 1. Подраздел «Относительное количество типов подключения», первый абзац. Я не нахожу Таблицу 1, которая необходима для интерпретации Рисунка 1 – дополнения к рисунку 4.
Рецензент, возможно, не смог найти Таблицу 1, поскольку она представлена в виде отдельного машиночитаемого дополнительного файла значений, разделенных запятыми (CSV).
Синаптические числа, подраздел «Структура и сила возбуждающих связей». Документирование номеров синапсов хорошее, и интерпретация, как правило, уместна, но авторы должны внимательно просмотреть свой текст, чтобы проверить тенденцию приравнивать количество синапсов к силе синапсов и строго различать семантические ссылки на эти два.
Мы явно отмечаем во введении, что сила синаптической связи может зависеть от многих анатомических особенностей.Количество синапсов на одно соединение – лишь одна из этих функций. Одна из целей нашего исследования состояла в том, чтобы сделать вывод, какие особенности на самом деле вносят наибольший вклад в наблюдаемые физиологические вариации силы связи. Мы начнем с того факта, что ипсилатеральные связи ORN → PN, как известно, на 30-40% сильнее, чем контралатеральные связи (Gaudry et al. 2013), и мы спрашиваем, какие анатомические особенности могут объяснить это физиологическое различие. Мы обнаружили, что количество синапсов на одно соединение примерно на 27% выше на ипсилатеральной стороне (рис. 4D), в то время как нет систематических различий в дендритных свойствах ипсилатеральных и контралатеральных синапсов (судя по амплитудам mEPSP, которые отражают расположение синапсов на дендрите). и электрические фильтрующие свойства самого дендрита, рис. 4E).Этот результат свидетельствует о том, что количество синапсов является основным фактором изменения прочности соединения. Более того, мы не обнаружили существенной разницы между ипси- и контралатеральной зоной постсинаптического контакта PN. Контралатеральные Т-образные стержни оказываются на 8% больше, чем ипсилатеральные Т-образные стержни, но эта разница относительно небольшая, и это объясняется тем фактом, что Т-образные стержни растут с увеличением количества постсинаптических элементов, а контрлатеральные проекции ORN нацелены на большее количество элементов. профили не-PN, чем профили ипсилатеральных ORN (Рисунок 4 – приложение к рисунку 1).Наконец, когда мы просто сосредотачиваемся на ипсилатеральных связях, мы снова обнаруживаем, что количество синапсов на одно соединение снова является наиболее изменчивой анатомической характеристикой синаптических связей (CV = 0,31, рис. 5C) при сравнительно меньших вариациях общих дендритных свойств соединения (CV амплитуда mEPSP = 0,014, Рисунок 5E; Эффективность суммирования CV = 0,041, Рисунок 5G) или средний размер синапса для каждого соединения ( объем CVT-стержня = 0,13, площадь постсинаптического контакта CV = 0.13, рис. 1 – приложение к рисунку 6г). Взятые вместе, эти результаты свидетельствуют о том, что число синапсов является основным анатомическим признаком, влияющим на колебания прочности соединения, хотя другие анатомические особенности также вносят свой вклад. Это важный результат для данной области, потому что он устанавливает номер синапса как разумный показатель силы соединения – с оговоркой, что эта функция определенно не улавливает все вариации силы соединения.
В контексте рукописи мы используем фразу «сила связи» только тогда, когда намеренно говорим о силе физиологической связи – либо в контексте опубликованных исследований, в которых непосредственно измерялись синаптические реакции, либо в контексте нашей компартментальной модели, который использует параметры, измеренные в опубликованных электрофизиологических исследованиях, для экстраполяции наших анатомических измерений на физиологические прогнозы.Мы подчеркиваем, что число синапсов является разумным показателем силы соединения, но мы также проясняем, что это всего лишь приблизительный показатель, потому что число синапсов – не единственная анатомическая характеристика, которая способствует изменчивости синаптических соединений.
Также важно помнить о глубине обработки, вносимой синапсами обратной связи, для которых усиление передачи отражает не только количество синапсов прямого распространения, но и синапсов восходящего направления. Таким образом, помимо усиления отдельных синапсов, схемы обратной связи наследуют усиление сигнала от своих синаптических входов.
Мы согласны с рецензентом в том, что соединения с прямой связью, на которых сосредоточено внимание в этом исследовании (соединения ORN → PN), не являются единственными важными элементами этой схемы. Мы действительно сообщаем данные о PN → PN синапсах, ORN → ORN синапсах и PN → ORN синапсах, хотя мы не фокусируемся ни на одном из этих типов соединений в последней части исследования. Мы также четко заявляем о том, что в цепи есть много клеток, которые мы не полностью реконструировали (клетки, которые мы называем мульти-клубочковыми клетками, которые в основном являются локальными нейронами).Конечно, некоторые из этих дополнительных клеток, вероятно, опосредуют обратную связь. Мы ожидаем, что элементы обратной связи этой схемы (например, локальные нейроны, которые опосредуют подавление обратной связи) будут описаны в будущих исследованиях ssEM.
Матрица смежности на рис. 1 – дополнение к рисунку 4 напоминает ряд таких матриц, которые уже опубликованы, с верхними числами около 50 синапсов, что соответствует таковым в других областях мозга, которые авторы могли указать. Каждый перехват представляет собой место предполагаемой передачи между одним пре- и постсинаптическим нейроном, но в тексте следует признать, что, поскольку каждый пресинаптический сайт имеет несколько постсинаптических элементов, ряд перехватов в матрице обязательно скоординированно связаны.Фактически, я не нахожу простой метрики синаптической дивергенции, показанной на этом рисунке или где-либо еще в тексте. Сколько диад, триад и т.д. включено в матрицу. Как вариант, сколько монад?
Мы согласны с рецензентом, что эта информация должна быть сообщена. Мы добавили рисунок 1 – приложение 5, показывающее распределение полиад в выходных синапсах ORN в DM6.
Относится к предыдущему пункту: Рисунок 1 – дополнение к рисунку 3. Это отдельные изображения синапсов, которые не могут выявить профили выше и ниже плоскости сечения.Тем не менее, очевидно, что на сайтах единственного выпуска есть несколько дендритов, что действительно можно увидеть в некоторых разделах. Эта информация представлена на Рисунке 4 – приложение к рисунку 1E, но только в виде средних значений для сравнения ORN Ipsi и Contra.
См. Ответ на предыдущий пункт рецензента. Теперь мы более четко проясним существование диад, триад и т. Д. На рисунке 1 – добавлении к рисунку 5.
Авторы заявляют, что «Все события синаптической проводимости имели одинаковый размер и форму.«Это предположительно предполагает, что эти события проводимости были общими в одном и том же пресинаптическом участке? Представляют ли авторы, что каждый постсинаптический дендрит видит одно и то же облако высвобождения нейротрансмиттера, или это облако разделяется между несколькими дендритами. Если это число варьируется, означает ли это эта передача изменяется в зависимости от количества дендритов или дендритный сигнал не изменяется; как результат влияет на моделирование авторов? Я не думаю, что есть простой ответ, но эти проблемы следует, по крайней мере, признать, и насколько возможно обсуждается.
Когда мы говорим: «Все события синаптической проводимости имели одинаковый размер и форму», мы имеем в виду все события проводимости на всех постсинаптических участках периода. В сущности, наша модель предполагает, что появление спайка на Т-образной полосе всегда открывает одинаковое количество постсинаптических ионных каналов во всех клетках, постсинаптических к этой Т-образной полосе. Мы изменили текст рукописи, чтобы сделать это предположение как можно более четким. Это предположение, очевидно, является упрощением, но построение модели всегда связано с некоторыми упрощениями, и мы думаем, что это не является необоснованным упрощением.Конечно, важно, чтобы одни Т-образные стержни были привязаны к одному постсинаптическому профилю, а другие – к двум профилям (диадам) или трем профилям (триадам), или даже больше. Однако мы не думаем, что синаптическая проводимость обязательно слабее в случаях, когда большее количество дендритов «разделяют» одну и ту же Т-образную перемычку, потому что нет причин, по которым концентрации нейротрансмиттеров обязательно ниже в диадических или триадных связях по сравнению с монадическими связями. Примечательно, что мы находим, что объем Т-образного стержня растет с количеством профилей, постсинаптических по отношению к Т-образному стержню (Рис. 1 – рисунок в приложении 6А), что может указывать на механизм, посредством которого квантовое содержание увеличивается, чтобы соответствовать объему синаптической щели.
Рисунок 1 – дополнение к рисунку 4. Кроме того, я считаю, что использование черного цвета для 0 синапсов сбивает с толку, не в последнюю очередь потому, что мне трудно различать различия в тоне для темных перехватов, имеющих мало синапсов, и рекомендую изменить график этой матрицы, используя белый цвет перехватывает 0 синапсов. Как вариант, переверните весь спектр тонов, чтобы 50+ был черным, а 0 – белым.
Благодарим рецензента за это предложение. Пожалуйста, найдите Рисунок 1 – Приложение 4 к рисунку, обновленное в новой редакции.
Методы. Из вырезанных 1917 секций, как нам сообщается в подразделе «Крупномасштабная визуализация и выравнивание ПЭМ», сколько секций было потеряно, по всей видимости, 145, от 1 до 6, около 7% от общей серии – так что примерно равно количество сирот, но не количество срезов, необходимых для охвата клубочков DM6, и, следовательно, не то, какая часть из них была каплями. Потеря 6 секций (0,3 мкм) наносит большой ущерб непрерывности любой серии, но, по крайней мере, авторы честно сообщают о них.Интересно, сколько из этих капель привело к появлению сирот, и были ли они одинаковыми на левой и правой сторонах DN6? Авторы, я думаю, должны попытаться решить эти вопросы.
В пересмотренных Методиках мы приводим следующие подробности относительно утраченных секций: «В пределах части объема, охваченной клубочками DM6, было 9 односекционных потерь, 2 случая 2-секционных потерь, 1 случай 3-секционных потерь. , и 2 случая 4-х секционных потерь (потери относятся к последовательным секциям).Конечно, потерянные разделы прискорбны, но они также не редкость в такого рода наборах данных. Мы выбрали DM6 специально, потому что ни один из 5-секционных или 6-секционных потерь не пересекал этот клубок. Некоторые из бесхозных фрагментов, которые мы реконструировали, являются следствием потери сечения, но другие бесхозные фрагменты не примыкают к потерям сечения, а вместо этого возникают из-за неоднозначности, связанной с тем, как плоскость среза пересекает нейрит.
https://doi.