
Ремонт стабилизаторов напряжения Ресанта – особенности ремонта
Стабилизаторы напряжения «Ресанта» используются во многих домах для обеспечения стабильной работы и защиты «здоровья» электрических приборов. В результате домашняя техника работает в течение длительного времени и почти не подвергается ремонту.
Надо сказать, что самому стабилизатору напряжения тоже необходимо соблюдение условий эксплуатации и периодический уход. Иначе аппарат может выйти из строя и ему потребуется ремонт. Помимо этого, отслужив достаточно большой срок, прибор может поломаться просто по причине износа деталей.
Эта статья посвящена тонким местам стабилизаторов бренда «Ресанта». Рассмотрим, как ремонтируются вышедшие из строя детали, а также восстанавливается полная работоспособность прибора.
Степень сложности ремонта стабилизаторов напряжения
Все приборы стабилизации оснащены защитными функциями, с помощью которых контролируются технические показатели на соответствие заявленным данным и условиям эксплуатации.
Прежде всего, требуется:
- проверка на наличие КЗ, входного и выходного напряжения, температурного режима компонентов;
- изучение высвеченного на дисплее кода ошибки.
Наиболее трудно определить неисправность симисторных ключей прибора, так как их управление связано со знанием электроники. При ремонте не обойтись без принципиальной схемы, измерительных средств, в том числе осциллографа. По контрольным точкам снятых осциллограмм определяются повреждения в структурном модуле устройства. Затем предстоит проверка каждой радиодетали и узла на предмет дефекта.
В стабилизаторах релейного типа нередко причиной неполадок становится реле, предназначенное для переключения обмоток трансформатора. Частые переключения контактов реле приводят к их выгоранию, заклиниванию, или перегоранию самой катушки. Если пропадает напряжение либо выходит сообщение об ошибке – стоит проверить все реле.
Наиболее прост ремонт электромеханического стабилизатора, у которого работа и реакция на изменение параметров сети становятся очевидными сразу после снятия корпуса. Недаром простая конструкция и высокая точность стабилизации делают эти модели весьма распространенными.
Виды неисправностей стабилизаторов напряжения
Ремонт электромеханического типа
Распространенной проблемой таких приборов является перегрев. Поэтому раз в 2 месяца следует предавать устройство техническому обслуживанию. Важной частью ремонта считается именно чистка элементов.
Примером могут служить характерные поломки распространённого стабилизатора АСН-10000/1-ЭМ. Устройство состоит из трёх одинаковых частей — из трёх 1-фазных стабилизаторов, предназначенных для стабилизации только своей фазы. Сердцем аппарата является повышающий автотрансформатор. Он же вместе с контактором и вводным автоматом относится к силовой части.
Принципиальная схема АСН-10000/1-ЭМ приведена на рисунке ниже.
В основе принципа действия электромеханических выравнивателей лежит плавное регулирование выходных параметров. Напряжение изменяется благодаря скольжению электрического контакта по обмотке автотрансформатора посредством электрического привода. На оси электродвигателя крепится ползунок, который перемещаясь, нормализует выходные параметры.
Заслуживает особого внимания следующая характерная неисправность, возникающая в процессе эксплуатации электромеханических стабилизаторов и методы ее устранения – отсутствие стабилизации выходного напряжения.
Первый признак такой неполадки – может ощущаться запах тлеющих деталей. Реверсивный двигатель недаром зовут «ахиллесовой пятой» электромеханических приборов. Контроллером стабилизатора напряжения постоянно отслеживается значение выходных параметров. Ротор постоянно вращается и это постепенно изнашивает сам двигатель.
Одна неисправность может повлечь за собой другие, например, выход из строя целого каскада управления электродвигателем, собранного на паре транзисторов. Помимо этих элементов от перегрева плавятся резисторы, стоящие в их коллекторной цепи.
Помимо этих элементов от перегрева плавятся резисторы, стоящие в их коллекторной цепи.
Конечно, изношенный электродвигатель лучше заменить, но бывает умелая попытка привести его в действие, венчается успехом. Это и есть самый простой способ реанимации двигателя:
- отключение двигателя от схемы;
- подача на его выводы 5 В от мощного источника питания, к примеру, от компьютерного БП ATX.
При этом получается отжиг мелкого «мусора» на щётках двигателя. Нормальный ток электропотребления движка должен не выходить за пределы 90–160 мА. Поскольку двигатель реверсивного типа, то напряжение необходимо подавать не менее двух раз со сменой полярности. После этих воздействий работоспособность агрегата временно восстанавливается.
Другой вариант решения проблемы – небольшая замена схемы с сужением диапазона регулировки. Просто щетка будет ездить по-другому, в обход выгоревших участков дорожки трансформатора.
Ремонт релейных стабилизаторов
В качестве примеров рассмотрим ремонт:
Ресанта АСН-500/1-ц.
Наиболее частыми ошибками являются сообщения «L» и «H», что означает начальные буквы английских слов «низкий» и «высокий». То есть показатели выходят за пределы допустимых параметров. На прежних релейных стабилизаторах Ресанта со стрелочными индикаторами можно было видеть изменение выходного напряжения в пределах 204–235 В при переключении ступеней. На нынешней аппаратуре по записи видно 220 В, а по факту те же +- 6%, согласно паспортным данным.
Случается проблема реле медленно переключается, что влияет на защитное отключение компрессора кондиционера. Дело в том, что производителем используются дешёвые конденсаторы весьма низкого качества. Если заменить электролиты – проблема будет решена.
Главное, не стоит забывать о мощности. То, что написано на шильдике корпуса, справедливо для входного напряжения 200 В, в реальности для заниженного (170–180 В) мощность должна быть в 2 раза меньше.
Ресанта СПН-9000.
В основе принципа действия этого релейного стабилизатора лежит ступенчатое регулирование выходного напряжения. Стабилизация обеспечивается посредством микропроцессора. Коммутация отводов автотрансформатора выполняется пятью мощными реле, которые управляются транзисторными ключами. Стабильность выходного напряжения зависит от дискретности переключения (5–20 В).
Стабилизация обеспечивается посредством микропроцессора. Коммутация отводов автотрансформатора выполняется пятью мощными реле, которые управляются транзисторными ключами. Стабильность выходного напряжения зависит от дискретности переключения (5–20 В).
Основная болезнь СПН-9000 – обгоревшие либо залипшие контакты в реле. Эти неполадки довольно часто возникают в процессе эксплуатации релейного стабилизатора. А также при несоответствии входного напряжения диапазону пороговых значений стабилизация не станет работать. Бывает, сразу при включении прибора выбивает предохранители, так срабатывает защита от КЗ.
По причине неисправности реле «летят» транзисторные ключи. Реле подлежат замене или реставрации. Для этого необходимо убрать крышки с реле, после снять подвижный контакт, освободить его от пружины и наждачной бумагой аккуратно очистить все контакты реле. В завершение очистить все контакты специальным бензином и собрать реле в обратном порядке. Затем впаять все транзисторы, и проверить на целостность переходов. Если понадобится, заменить транзисторы на новые.
Если понадобится, заменить транзисторы на новые.
Заключение
Если вам нужно подключить к стабильнику предположим электрическую печь (9 кВт), то лучшего прибора, чем стабилизатор напряжения Ресанта для этого не найти. А если при этом возникнут мелкие недочеты, то сервисные мастерские быстро и профессионально устранят их на основании гарантийных обязательств. Своевременно сделанный ремонт – залог долговечности и надёжности прибора и после гарантийного срока.
Поломки бывают различные, и иногда сложно понять, то ли просто не соблюдены условия эксплуатации по инструкции, то ли аппарат неисправен. Однако, неполадки могут существовать, и в итоге в самый неподходящий момент может возникнуть проблема. Правильно установить «диагноз» и эффективно устранить их всегда поможет ремонтная компания.
На видео: простой ремонт стабилизатора РЕСАНТА 15 квт 3 фазы.
Стабилизатор Ресанта АСН – 5000 / 1-Ц
Технические свойства:
- Тип сети – 1-фазная.
- Интервал напряжений входа от 140 — 260 вольт.
- Мощность 5 киловатт.
- Напряжение выходное 220 вольт.
- Вид подключения – колодки клеммные.
- Байпас – имеется.
- КПД более 97%.
- Температура работы от 0 — 45 градусов.
- Влажность до 80%.
- Охлаждение – без вентилятора.
- Размеры – 220 х 230 х 340 мм.
- Вес 13 кг.
- Степень защиты – IР 20.
Стабилизатор Ресанта АСН-5000/1ц является релейным видом стабилизирующего прибора, используемого в 1-фазных сетях переменного тока для равномерного питания электрических устройств с общей мощностью не выше 5 киловатт.
Принцип работы
Разберемся в принципе действия стабилизатора. Выходное напряжение выравнивается в этом приборе, как и во всей линейке Ресанты, по принципу дискретной регулировки. Значит, можно сказать, что за выходное стабильное напряжение отвечает автоматический трансформатор, который имеет множество понижающих и повышающих обмоток, микроконтроллер и реле.
Напряжение бытовой сети определяется и сравнивается контроллером с напряжением выхода и подается сигнал на реле для коммутации обмотки, которая может создать необходимое напряжение на выходе прибора.
Ступенчатая коррекция напряжения выхода прибора, выполняемая с помощью коммутации разных обмоток, имеет достаточно большую точность, и составляет +8%, чего хватает для нормальной работы всех бытовых устройств. Намного важнее, что быстродействие регулирования стабилизатора, действующего по этому принципу, очень высокое, и равно 35 мс. Это очень неплохой результат.
Напряжение сети, при котором прибор способен работать и исполнять свои задачи, колеблется от 140 до 260 вольт. В приборе имеется несколько защит.
- Защита от низкого напряжения входа.
- Защита от чрезмерной нагрузки по току.
- Защита от КЗ (короткого замыкания).
При входном напряжении ниже 190 вольт выходная мощность начинает снижаться, а при входном напряжении 140 вольт мощность падает наполовину. Если верхний предел интервала напряжения входа длится очень долго, то срабатывает защитная функция, отключающая выходное напряжение, а прибор переходит в аварийный режим.
Внешний вид
Модель стабилизатора АСН 5000 1 ц изготовлена в компактном металлическом корпусе, обеспечивающем защиту по степени IР20, что означает о возможности применения стабилизатора при влажности не выше 80% и температуре до 45 градусов.
На лицевой части находится цифровой дисплей, которые входят в мощные модели приборов, а также выключатель с двумя секциями. Левая секция предназначена для выключения и включения опции «Байпас», позволяющей коммутацию потребителей напрямую в обход прибора. Правая секция отвечает за подключение к сети.
LCD-дисплей
На дисплее имеются необходимые данные для контроля работы прибора. Вверху расположены различные индикаторы:
- Задержка.
- Работа.
- Защита.
Каждый из них включается в соответствующей ситуации. Индикатор «работа» включен постоянно при работающем приборе.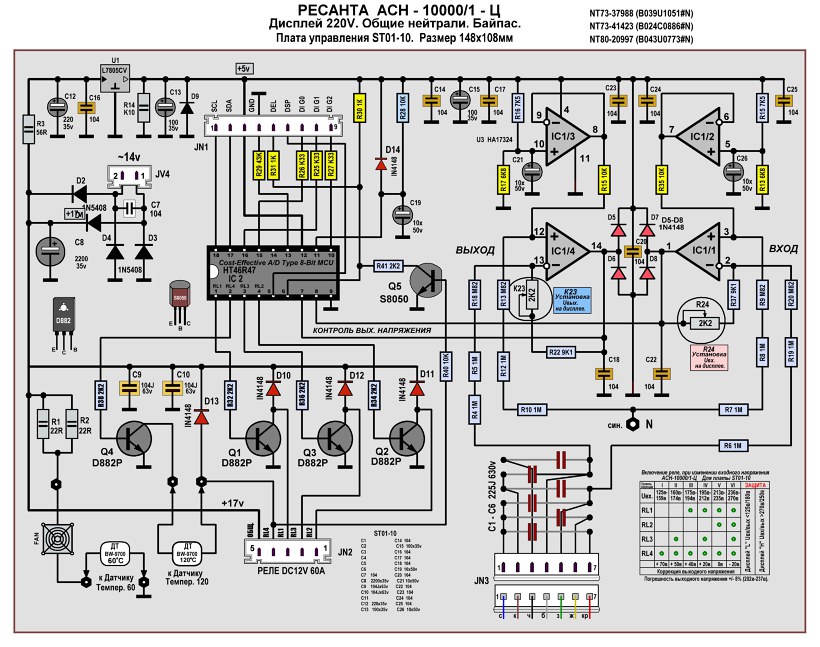 «Защита» светится при срабатывании какой-либо защитной функции. «Задержка» работает в момент включения стабилизатора и сработки защиты.
«Защита» светится при срабатывании какой-либо защитной функции. «Задержка» работает в момент включения стабилизатора и сработки защиты.
Далее расположены значения напряжения выхода и входа, нагрузка, ее изменение. Внизу находятся индикаторы снижения выхода напряжения, либо его возрастания более 245 вольт.
Задняя панель
Сзади находится клеммная колодка, к которой подсоединяется сеть питания и потребители нагрузки к стабилизатору.
Этот вид прибора, как и вся линейка Ресанты, имеет малое потребление электроэнергии, что позволяет достичь повышенного КПД до 97%, а также низкий уровень шума, создавая тем самым должный уровень комфорта.
Однофазный стабилизатор напряжения электронного типа Ресанта АСН-5000/1-Ц 63/6/6
Однофазный стабилизатор напряжения электронного типа Ресанта АСН-5000/1-Ц обеспечивает эффективное электропитание любой техники, защищая от возможных повреждений и сбоев. Данная модель разработана для защиты устройств от аварийных скачков электроэнергии в пределах небольших жилых помещений и производственных комплексов. Прибор реализует уверенную работу различных устройств в условиях нестабильного по значению напряжения.
Прибор реализует уверенную работу различных устройств в условиях нестабильного по значению напряжения.
Особенности модели:
- Сохранение работоспособности однофазных электроприборов низкой и средней мощности при установке в отапливаемом помещении;
- Наличие цифрового дисплея для индикации показаний работы стабилизатора;
- Регулировка выходного напряжения в широком диапазоне без искажения формы сигнала;
- Высокое быстродействие и автоматическое отключение нагрузки при превышении предельного значения входного тока;
- Автоматическое отключение нагрузки при превышении предельного значения выходного напряжения.
Принцип работы
Регулировка напряжения происходит за счет переключения обмоток на трансформаторе при помощи реле. Поэтому данный вид стабилизаторов называется “релейный”. Осуществляется ступенчатая регулировка. При ступенчатой регулировке точность выходного напряжения возрастает до 8%, это 17,6 В, что вполне безопасно для всех бытовых приборов, по ГОСТ допустимо 10%. Но за счет этого сокращается время регулировки, оно минимально и составляет менее 15 миллисекунд, то есть менее 1 секунды! Такой стабилизатор стоит устанавливать в места где входное напряжение постоянно изменяется.
Но за счет этого сокращается время регулировки, оно минимально и составляет менее 15 миллисекунд, то есть менее 1 секунды! Такой стабилизатор стоит устанавливать в места где входное напряжение постоянно изменяется.
Общие сервисные функции стабилизатора
- Регулировка выходного напряжения в широком диапазоне, дискретным способом без искажения формы сигнала.
- Широкий диапазон входных напряжений 140-260 В.
- Высокое быстродействие.
- Контроль над выходным напряжением с помощью встроенного в корпус вольтметра.
- Автоматическое отключение нагрузки при превышении предельных значений выходного напряжения (максимального и минимального).
- Автоматическое отключение нагрузки при коротком замыкании.
- Автоматическое подключение нагрузки при восстановлении выходного напряжения в пределах рабочего диапазона.
- Индикация режимов работы.
Стабилизатор Ресанта ACH-5000/1-Ц имеет мощность 5 кВт, данной мощности хватает, чтобы питать отдельные потребители, или несколько потребителей, но суммарное потребление не должно превышать установленный мощностной номинал. Диапазон входных напряжений стабилизатора 140-260 Вольт, но при понижении входного напряжения ниже 190 Вольт начинается потеря выходной мощности, при минимальном входном напряжении 140 Вольт выходная мощность сократиться на 50% и составит 2,5 кВт.
Диапазон входных напряжений стабилизатора 140-260 Вольт, но при понижении входного напряжения ниже 190 Вольт начинается потеря выходной мощности, при минимальном входном напряжении 140 Вольт выходная мощность сократиться на 50% и составит 2,5 кВт.
Рекомендуем выбирать модель стабилизатора напряжения с небольшим запасом по мощности, который позволит создать резерв для подключения нового оборудования.
При длительных превышениях допустимых значений входного напряжения система защиты отключит выходное напряжение, а сам стабилизатор уйдет в режим защиты. При перегреве стабилизатора так же произойдёт аварийное отключение выходного напряжения. Максимальное температурное значение обмотки трансформатора может достигать 70 °С, нагрев трансформатора напрямую зависит от температуры окружающей среды. Стабилизатор так же защищён от короткого замыкания при помощи предохранителя.
Описание индикаторов дисплея
Стабилизаторы напряжения, оборудованы LCD-дисплеями. Ниже представлено схематичное изображение дисплея с указанием всех индикаторов.
Ниже представлено схематичное изображение дисплея с указанием всех индикаторов.
- Задержка – индикатор активен при включении стабилизатора и при срабатывании одной из защит, (низкое/высокое напряжение, перегрев, перегрузка). Дополнительно на дисплее отображается обратный отсчет времени задержки.
- Работа – индикатор активен постоянно при включенном устройстве.
- Защита – индикатор активен при срабатывании одной из защит.
- Индикатор нагрузки – изменяется пропорционально току нагрузки.
- Гиря – часть индикатора нагрузки – индикатор активен постоянно при включенном устройстве.
- Ресанта – индикатор появляется при включении (буква за буквой), и активен постоянно при включенном устройстве.
- Перегрев – индикатор активен при срабатывании защиты от перегрева.
- Перегрузка – индикатор активен при срабатывании защиты от перегрузки.
- Пониженное напряжение – индикатор активен при выходном напряжении
- Строка состояния – представляет собой 8 точек.
 При включении каждая точка соответствует 1 секунде задержки при включении.
При включении каждая точка соответствует 1 секунде задержки при включении. - Повышенное напряжение – индикатор активен при выходном напряжении >245 В.
- Входное напряжение – отображает входное напряжение.
- Выходное напряжение – отображает выходное напряжение.
Заказывайте Стабилизатор напряжения Ресанта АСН 5000/1 ЭМ по доступным ценам, 49677520
Качественный стабилизатор Ресанта АСН 5000/1 ЭМ
Ни для кого не секрет, что скачки напряжения в электрической сети чрезвычайно вредны для современных бытовых приборов. Для того, чтобы выравнивать подобные колебания были разработаны стабилизаторы. Они позволяют эффективно справляться как с кратковременными колебаниями, так и с длительными скачками напряжения. Для частных домов, квартир, кафе и других небольших заведений целесообразно использовать стабилизатор напряжения Ресанта АСН 5000/1 ЭМ! Он отлично справится с поставленной перед ним задачей!
Что о нем следует знать?
Преимущества модели
1 | Доступная стоимость прибора. |
2 | Максимальная надежность в работе — год гарантии. |
3 | Выпускается известной на отечественном рынке фирмой. |
4 | Заявленная мощность устройства полностью соответствует реальным показателям. |
Заказывайте стабилизатор напряжения Ресанта АСН 5000/1 ЭМ у нас и останетесь довольны его работой!
Схема заказа Стабилизатора напряжения Ресанта АСН 5000/1 ЭМ
Оформить заказ на сайте или по телефону
Консультация менеджера и согласование всех нюансов
Оплата удобным способом
Доставка выбранным способом
| Дополнительные характеристики | |
|---|---|
| Товар сертифицирован | Да |
| Гарантия | 1 год |
| Вес в упаковке | 16.41 кг |
| Вес без упаковки | 14.81 кг |
| Страна производства | Китай |
| Габариты, см | 18,3 х 32,5 х 39,5 |
| Страна бренда | Латвия |
| Комплектация | Стабилизатор напряжения РЕСАНТА Инструкция Упаковка |
| Технические характеристики | |
|---|---|
| Тип | Электромеханический |
| Мощность | 5000 Вт |
| Напряжение сети | 220 В |
| Тип входного напряжения | Однофазный (220 В) |
| Охлаждение | Естественное |
| Размещение | Напольное |
| Класс защиты | IP20 |
| Питание | От электросети |
| Выходное напряжение | 216 – 224 В |
| Габариты, см | 18,3 х 32,5 х 39,5 |
| Высоковольтная защита | 240 – 250 В |
| Вес без упаковки | 14. 81 кг 81 кг |
| Страна бренда | Латвия |
| Точность стабилизации | 2 % |
| Скорость стабилизации | 40 В/с |
| Частота | 50 Гц |
| Рабочая температура | 0 – 45 °C |
| Возможности | цифровая индикация (вольтметр отображает входное и выходное напряжения) |
| КПД | 97 % |
| Страна производства | Китай |
| Влажность воздуха | 80 % |
| Штрихкод EAN-13 | 4606059015260 |
| Входное напряжение | 140 – 260 В |
| Максимальный потребляемый ток | 26.3 А |
| Серия | АСН |
| Комплектация | Стабилизатор напряжения РЕСАНТА Инструкция Упаковка |
| Штрихкод GTIN | 04606059015260 |
| Время отклика | 10 мс |
| Защита | от короткого замыкания, от перегрева, от повышенного напряжения |
| Вес в упаковке | 16. 41 кг 41 кг |
Купить
Стабилизатор не включается или выбивает автоматы. Основные неисправности и ремонт стабилизаторов
Как и любое сложное электронное устройство, стабилизатор напряжения иногда выходит из строя, сам выключается или выбивает автоматы или по крайней мере не корректно работает, гудит или пищит.
Причин может быть несколько, в зависимости от конкретной ситуации, и это может зависеть от неправильности использования или же зависеть непосредственно от типа и электронной начинки самого аппарата.
Попытки хозяев отремонтировать самому такое сложное устройство могут быть оправданы только в случае поверхностных причин поломки и небольшого понимания в принципе работы устройства.
Но не всегда это приводит к желаемому результату, а зачастую и вовсе может привести к полной поломке платы управления а также силовых ключей, что в итоге повысит стоимость ремонта в разы.
По этому лучше доверить ремонт специалистам, тем более в случае если стабилизатор на гарантии.
Но мы все же рассмотрим основные причины неисправностей, и методы их устранения.
Стабилизатор любого типа – это сложное электронное устройство и зачастую для выявления неисправности будут необходимы измерительные приборы и хотя бы некоторые познания в радиотехнике.
Как правило во всех стабилизаторах напряжения стоит целая система защиты целью которой есть защита силовых элементов от сгорания, защита по превышению мощности, перегреву устройства, а также защита выходного напряжения от аномальных скачков напряжения.
В основном вся защита стабилизатора реализована на плате управления, сложность схемы которой, зависит от типа стабилизатора.
Сложнее всего выявить неисправность в стабилизаторе на симисторных ключах, сложная схема управления требует проверки с помощью осциллографа или в крайнем случае можно применить метод последовательной проверки каждого элемента схемы.
В релейных стабилизаторах напряжения частой причиной поломки является реле которое переключает обмотки трансформатора. При частом нестабильном напряжению в сети реле выполняют множество переключений на протяжение дня, со временем контакты реле подгорают, еще могут залипнуть, а бывает и сама катушка реле перегорает. В таких случаях может появится сообщение об ошибке, стабилизатор может просто выключится, а может быть и куда хуже вплоть до внутреннего замыкания с соответствующими последствиями.
Самым простым в ремонте можно назвать сервоприводный стабилизатор, после снятия крышки устройства можно наглядно рассмотреть его поведение и попытаться выявить причину логическими выводами.
Основные и общие неисправности стабилизатора
Стабилизатор отключается. Скорее всего, в большинстве случаев, отключение защитное и срабатывает при критическом повышение или понижение напряжения. После восстановления подходящего напряжения – питание восстанавливается сразу или через 5 секунд если установлены такие настройки.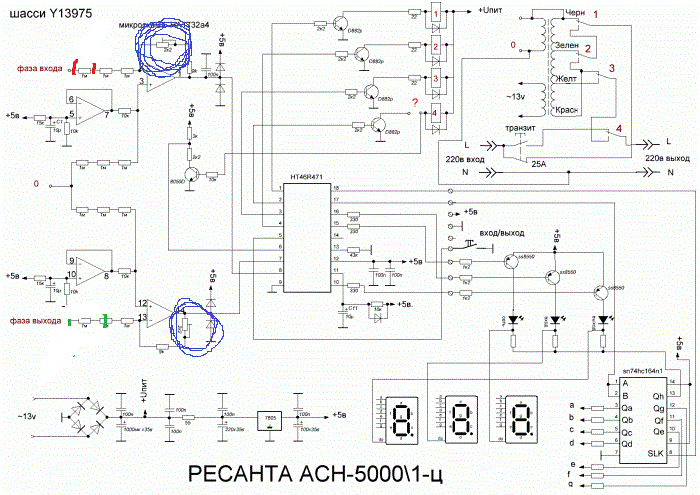
Но следует заметить что не все стабилизаторы так “следят” за нижней границей напряжения и часто при снижению напряжения до “нестабилизируемых” нижних границ напряжение падает без отключений. В таких случаях рекомендуется использование в щитке реле напряжения в котором настраивается верхний и нижний границы нужного вам напряжения, при выходе за их пределы – реле отключит нагрузку от сети.
Стабилизатор может также отключится и при превышению нагрузки (перегрузке) в таком случае оно будет сделано ступенчато, а при двукратной перегрузке будет выполнено моментальное отключение стабилизатора.
Кроме того выключится стабилизатор может при сработке термодатчика от перегрева силовых элементов или трансформатора.
Если стабилизатор часто выключается, нужно проверить входное напряжение, при его допустимых значениях – отключить нагрузку и убедится в том что в ней нет замыканий.
Если без нагрузки стабилизатор работает значит нагрузка неисправна, убедится в этом можно, подключив к стабилизатору эквивалентную нагрузку и если стабилизатор будет с ней работать то в первой нагрузке замыкание, если не будет работать с эквивалентной нагрузкой – то стабилизатор стал неисправным. Также о неисправности будет говорить тот факт если на входе напряжение будет в пределах нормы а стабилизатор не будет включатся.
Также о неисправности будет говорить тот факт если на входе напряжение будет в пределах нормы а стабилизатор не будет включатся.
Выбивает автомат при включение стабилизатора. Срабатывает защита которая ясно дает нам понять о коротком замыкание или значительной перегрузке. Впервую очередь нужно попробовать включить стабилизатор без нагрузки, тем самым сузив круг возможных причин. Если автомат выбивает без нагрузки значит стабилизатору потребуется серьезный ремонт. Прежде всего необходимо обратить внимание на мощность стабилизатора и автомат (по номиналу), может быть автомат на слишком малый ток, а стабилизатор во время включения потребляет большой ток. В некоторых (частых) случаях стабилизатор все же можно заставить работать если убрать заземление на сетевой вилке ( подключив стабилизатор с помощью переходника без заземления), но это не выход и скорее всего устройство придется ремонтировать.
Греется трансформатор стабилизатора (без нагрузки) Прежде всего нужно убедится в том что нагрузка выключена, если при этом трансформатор все же продолжает греться то возможно в трансформаторе произошло межвитковое замыкание, или что более вероятней – замыкание где то в переключателях (в зависимости от типа стабилизатора)
Например в релейном стабилизаторе следует обратить внимание на реле, а в симисторном – на силовые ключи. При пробое или замыкание (одного) силового элемента возникнет замыкание на одной из выходных обмоток, шаг напряжения на одной обмотке небольшой но все же достаточный чтоб перегреть трансформатор, а возможно и запустить защиту которая отключит устройство.
При пробое или замыкание (одного) силового элемента возникнет замыкание на одной из выходных обмоток, шаг напряжения на одной обмотке небольшой но все же достаточный чтоб перегреть трансформатор, а возможно и запустить защиту которая отключит устройство.
Реле можно осмотреть и прозвонить тестером (в выключенном состояние), убедится в отсутствие залипаний.
Симисторные или тиристорные ключи также можно проверить с помощью тестера. Между управляющим электродом и катодом сопротивление должно быть одинаковым при прямом и обратном измерении, а между анодом и катодом – стремиться к бесконечности.
В сервоприводных стабилизаторах, силовых ключей нет, но трансформатор может перегреваться из за забившихся в пространство между витками графитовых опилок, элементов гари и пыли. Такие устройства требуют периодической чистки рабочей контактной части витков трансформатора.
Поломка двигателя сервопривода или некорректная его работа, сюда же можно и причесть и обгорание и износ рабочей щетки что будет сопровождаться чрезмерным искрообразованием.
В сетях с частыми скачками напряжения двигатель сервопривода постоянно работает на износ, такое частое движение быстро вырабатывает определенный ресурс работы реверсного двигателя.
Поломка двигателя часто, за собой влечет также выход из строя выходного каскада управления сервоприводом, силовые транзисторы попросту перегорают.
В некоторых случаях двигатель можно попытаться реанимировать, разобрав и добравшись к его щеткам, очистить их от мелкой пыли и загрязнений. Собрав двигатель снова, произвести смазку редуктора и втулок на его якоре. Такое профилактическое обслуживание может значительно увеличить его ресурс работы, а к тому же уменьшить общий шум от работы сервоприводного стабилизатора.
Выход из строя реле. Часто такая поломка приводит также и к выходу из строя транзисторных ключей соответствующего реле.
В таких случаях и реле и транзистор подлежат замене на новые. В некоторых случаях изношенные контакты реле можно восстановить. Для этого разбирают корпус реле, затем снимают с пружины подвижный контакт. С помощью “нулевочной” наждачной бумаги, с контакта снимаются все нагоревшие частицы, после чего контакты протирают мягкой тряпочкой смоченной в спирте или растворителе.
С помощью “нулевочной” наждачной бумаги, с контакта снимаются все нагоревшие частицы, после чего контакты протирают мягкой тряпочкой смоченной в спирте или растворителе.
После восстановления реле, нужно обязательно убедится в исправности управляющих выходных транзисторов (типа SD882 или D882Р).
Помимо описанных выше поломок которые встречаются наиболее часто, часто можно столкнутся и с такими:
Дисплей. Хаотичное отображение на дисплее разных элементов или неполное отображение информации на дисплее может говорить о нарушение контакта между платой и дисплеем. Как правило для соединения там используют “токопроводящую резинку” которая прижимается между платой и стеклом ЖК-дисплея, в процессе постоянного нагрева стабилизатора и повышенной температуры внутри резинка пересыхает а плата может согнутся или незначительно деформироваться что вызовет потерю надежности контакта.
В сегментных дисплеях причины могут быть немножко другие.
В них зачастую причина кроется в плохой пропайке индикаторов и элементов платы. Элементы следует осмотреть на качество пайки, особое внимание уделив кварцевому резонатору и контролеру дисплея. Место соединения платы с дисплеем также осмотреть и при необходимости пропаять шлейф и контакты или очистить “токопроводящую резинку”.
Элементы следует осмотреть на качество пайки, особое внимание уделив кварцевому резонатору и контролеру дисплея. Место соединения платы с дисплеем также осмотреть и при необходимости пропаять шлейф и контакты или очистить “токопроводящую резинку”.
Поломка платы управления. Электронная плата управления у любого современного стабилизатора содержит множество радио элементов. Ее ремонт прежде всего, начинается с беглого осмотра всех элементов, их состояния и мест пропайки на плате. Обратить внимание на саму плату, почерневшие дорожки в местах перегрева и едва заметные микротрещины.
Очень часто можно заметить вздувшиеся электролитические конденсаторы. Часто конденсаторы внутри пересыхают и при этом теряют свою электрическую емкость.
Кроме того на плате можно выявить изменения оттенка радиоелементов от сильного перегрева, такие детали нужно выпаивать и проверять с помощью тестера и приборов.
Но как правило визуальный осмотр может только подсказать о масштабах случившейся неисправности, ну а сам ремонт таких плат не ограничивается заменой очевидно испорченных элементов и требует добавочной ревизии разных компонент при помощи особого оборудования. Поэтому, в случае если прозвонка силовых транзисторов и прочих элементов не обнаружила причины неисправности, ремонт платы управления лучше доверить специалистам.
Поэтому, в случае если прозвонка силовых транзисторов и прочих элементов не обнаружила причины неисправности, ремонт платы управления лучше доверить специалистам.
Стабилизатор гудит (шумит). Почти все стабилизаторы в процессе своей работы издают небольшие шумы, одни типы больше, другие меньше. Количество шума от стабилизатора будет напрямую зависеть от стабильности напряжения в сети, чем больше скачков и изменений напряжения происходит – тем больше стабилизатор должен выравнивать напряжение на выходе.
Наиболее шумными считаются сервоприводные стабилизаторы, постоянное включения реверсивного двигателя и его шум при движение графитового ползунка по обмоткам трансформатора приносят небольшой дискомфорт к которому со временем каждый владелец привыкает. Релейные стабилизаторы также издают щелчки при переключение обмоток трансформатора – тоже шум. Более благоприятными в этом плане можно считать симисторные и тиристорные стабилизаторы.
Едва слышное гудение сопровождает все стабилизаторы, источником звука есть сам преобразующий трансформатор и его гудение будет тем больше, чем больше разница входного и выходного напряжения и чем больше нагрузка в это время.
При повышенных шумах и гудению устройство лучше разобрать и осмотреть, возможно потребуется ремонт, а возможно профилактическое восстановление, например восстановление подвижной части электродвигателя сервоприводного стабилизатора.
Стабилизатор пищит. Здесь важно пищит он под нагрузкой или в холостом режиме. Отключаем нагрузку и прислушиваемся, в некоторых типах стабилизаторов (электронного типа) может быть слышен едва ощутимый писк, ето нормально.
Но если стабилизатор пищит (ощутимо) от повышения нагрузки, это может говорить о малом запасе прочности элементов конструкции аппарата, другими словами, если вы не перегружаете стабилизатор то он все же работает на пределе возможностей.
После успешного ремонта стабилизатор напряжения можно проверить с помощью ЛАТРа.
К ЛАТРу подключают проверяемый стабилизатор, а на выход стабилизатора подключают нагрузку в виде лампочки накаливания (примерно 60вт). Дальше изменяя напряжения на ЛАТРе, наблюдают за работой стабилизатора и параметрами напряжения на выходе.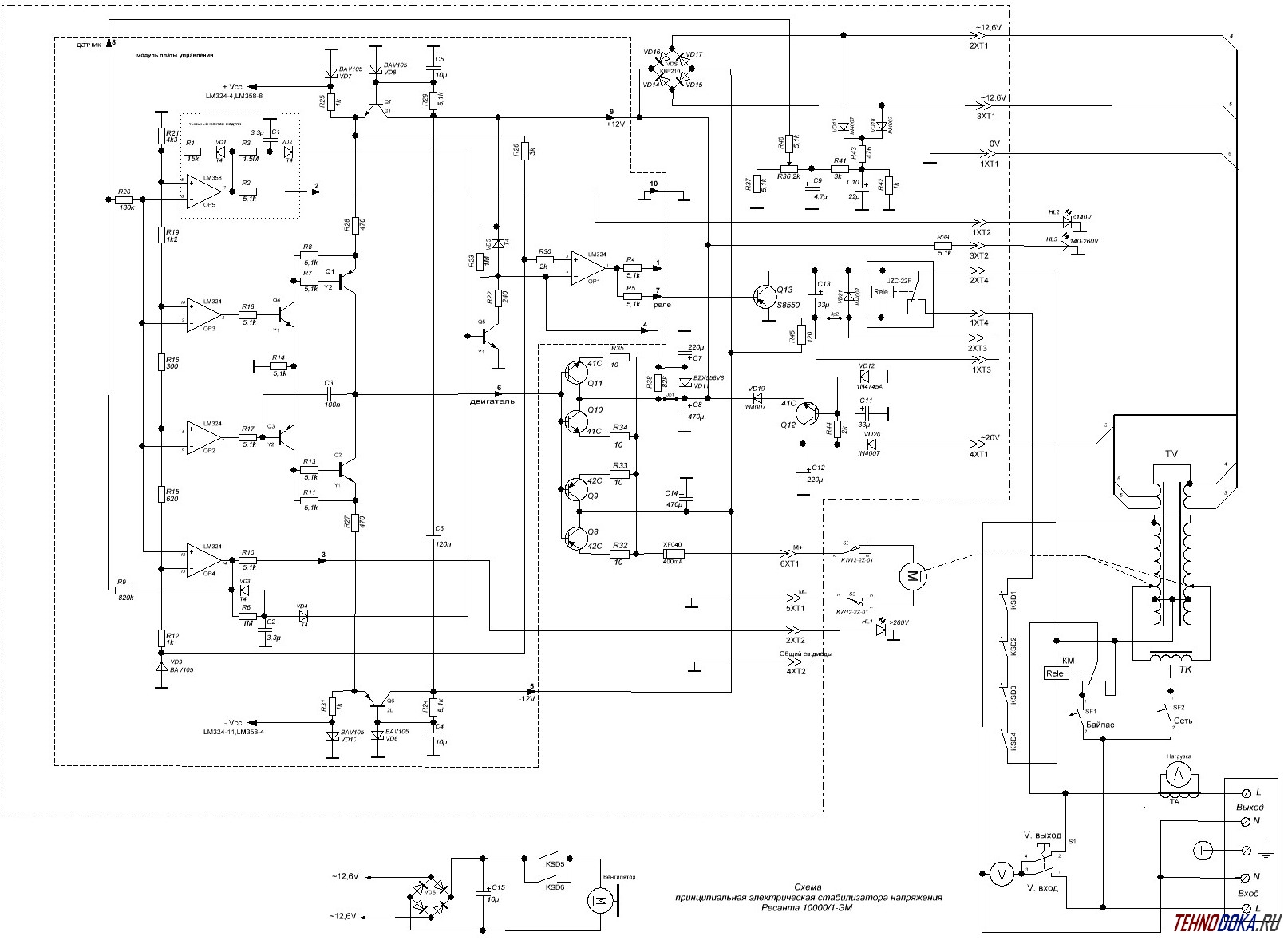
Напоследок дам несколько советов, которые помогут надолго сохранить прибор в рабочем состоянии:
- Следите за тем чтобы стабилизатор не работал долгое время при напряжение меньше 160 вольт. По крайней мере чтобы в такие моменты нагрузка на нем была сведена на минимум.
- При постоянно пониженном напряжение нужно приобретать и использовать специальные стабилизаторы, например у “Ресанта” есть некоторые модели позволяющие работать даже при 90 вольтах в сети.
- Суммарная мощность нагрузки должна быть хотя бы на 10% меньше мощности стабилизатора. При етом стараться одновременно не включать ее всю на длительное время.
- Подключая стабилизатор на весь дом необходимо оборудовать в щитке дополнительное УЗО с токовым номиналом не ниже чем у автомата на стабилизаторе.
- Очень важна правильная установка стабилизатора. Помещение где будет находится стабилизатор должно быть проветриваемым и сухим. Запрещается установка в нишах что будет нарушать воздухообмен и вызывать частый перегрев устройства.

электро схема мельницы фермер
2010-6-11 Схема технологическая Схема размещения Зерно из приемного бункера мельницы “Фермер-1” подается на вальцовый станок с крупной нарезкой рифлей (драная система).
» стержневая мельница. схема шаровой мельницы » чертеж шаровая мельница в компасе » дробилка щековая смд 506
2021-5-7 Рабочим мельницы является типовой проект мельницы Фермер – 4. Рисунок 5.1 Схема мельницы 1 – мельничный цех; 2 – склад готовой продукции в таре; 3
Возможна поставка мельницы с вашей технологической схемой. Возможна поставка отличной ржаной мельницы. В комплекте поставки электронный белизномер, лабораторный рассев.
Качаем схема электрическая газель 405 архивным файлом shema- .Электрическая схема автомобиля уаз фермер – Ищите любые файлы. где скачать электро схему по двигателю уаз 390945 фермер 409дв 2010год выпуск.
2017-11-10 Электросхема УАЗ 3160. Передний правый указатель поворота 1 1. Фары 2 3. Правый моторедуктор фароочиститсля 4 4. Прогивотуманные фары 5 6. Звуковой сигнал 7 7. Левый моторедуктор фароочистителя 8 8. Передний левый указатель поворота 9 9. Генератор 10 10.
Прогивотуманные фары 5 6. Звуковой сигнал 7 7. Левый моторедуктор фароочистителя 8 8. Передний левый указатель поворота 9 9. Генератор 10 10.
Продается мельничный комплекс: Мельницы Фермер-1 и Фермер-2, агрегат очистки РТ-АО3-3П, блок транспортировки РТ-БТ3-1, дополнительные бункеры для зерна на 7 т., для очистки и отволаживания, с суточным режимом работы.
2020-10-10 Мельница для зерна Фермер , зернодробилка, измельчитель зерн. Измельчители зерна или мельницы «Фермер» предназначены для приготовления корма домашним животным путем дробления сухого зерна и других зерновых культур в подсобных домашнем хозяйстве.
2021-6-18 Сетка на мельницах сменная, соответственно, Вы имеете возможность изменять фракцию помола, делая её по желанию крупнее или мельче. “Ураган-5К” также хорошо подойдет для дробления, например, древесного угля, щепы, ломаного печенья (делать крошку для тортов), корня девясил
Схемы электрооборудования автомобилей Газель ГАЗ-33021,2705 с двигателем 40522,4216 Схема
2020-1-28 Солододробилка своими руками – Спецтехника Зерно — один из основных видов корма для крс. Оно одинаково полезно и молочным животным, и
Оно одинаково полезно и молочным животным, и
2021-6-14 ОЛЛИ-ЮГ – продажа электропастухов OLLI и комплектующих для строительства электроизгороди Главная Компания ОЛЛИ-ЮГ – официальный представитель дистрибьютора компании Farmcomp OY в России, приветствует посетителей нашего
Молотковая дробилка: устройство и классификация, видеообзор бытового измельчителя для зерна
Мельницы-измельчители, купить в Минске. Подробная информация о товаре и поставщике. Краткое описание товара: мельницы предназначены для измельчения специй, перца, кофе, лекарственных трав.
Самодельная фреза для трактора Т-25 В сегодняшней статье мы рассмотрим один из главных вопросов агрономов, у которых есть трактор модели Т-25 – фреза.
Ножи для измельчителей Фермер 30мм -175мм. короткий 490101.. 12.00р. В корзину Сито боковое ИЗ-05/05м ИЗ-14/14М РФ д,5 190мм старый образец
Содержание мини трактора своими руками чертежи и размеры – Как сделать самодельный минитрактор своими руками – Perfil – SpegaSoft ForoКак сделать Самодельный минитрактор –
применения купить мини мельницу мельницаКупить Мини Мукомольный Завод/пшеничная Мука Мельница Цена оптом из Китая. Товары напрямую с завода производителя на при Технологическая линия по переработке угля
Товары напрямую с завода производителя на при Технологическая линия по переработке угля
Данная схема выполняется при небольших напорах от 1,5 до 4 м, реже до 6м, и небольшой мощности станции (от нескольких сотен киловатт до одного, реже двух мегаватт).
2021-1-12 Продажа – Мукомольные заводы и мельницы до 200 тыс. тонн в сутки в Краснодарском крае. электро и водоснабжение сетевое, есть скважина. В состав фермы входят: 7 коровников общей площадью
Молотковая дробилка: устройство и классификация, видеообзор бытового измельчителя для зерна
2020-1-28 Солододробилка своими руками – Спецтехника Зерно — один из основных видов корма для крс. Оно одинаково полезно и молочным животным, и
Содержание1 Как сделать муку в домашних условиях? Рецепт!1.1 Зачем делать домашнюю муку?1.2 Выбор инструмента для изготовления муки1.3 Как сделать муку в домашних условиях?2 Изготовление молотилки зерна своими руками2.1
Мельницы-измельчители, купить в Минске. Подробная информация о товаре и поставщике. Краткое описание товара: мельницы предназначены для измельчения специй, перца, кофе, лекарственных трав.
Краткое описание товара: мельницы предназначены для измельчения специй, перца, кофе, лекарственных трав.
Содержание мини трактора своими руками чертежи и размеры – Как сделать самодельный минитрактор своими руками – Perfil – SpegaSoft ForoКак сделать Самодельный минитрактор –
Посмотрите, что нашел пользователь Сергей Доведенко (wwwsdwwwsd) на Pinterest — самой большой в мире коллекции идей.
В основе транзисторных ключей модели АСН-5000/1-Ц (его схема приводится ниже) находятся транзисторы D882Р. Все эти транзисторы выпускает компания NEC. Рис. 2. Схема
Ножи для измельчителей Фермер 30мм -175мм. короткий 490101.. 12.00р. В корзину Сито боковое ИЗ-05/05м ИЗ-14/14М РФ д,5 190мм старый образец
Данная схема выполняется при небольших напорах от 1,5 до 4 м, реже до 6м, и небольшой мощности станции (от нескольких сотен киловатт до одного, реже двух мегаватт).
Организация (“Тепло Стиль”, ОГРНИП 313650105800040, ул. Ленина, 246А) является официальным дистрибьютором в Сахалинской области таких известных Южно-Корейских компаний как Naos co. , ltd и Cаммёнгтех, M.H. Korea ltd.
, ltd и Cаммёнгтех, M.H. Korea ltd.
10005708550_03
% PDF-1.6 % 1 0 объект > / OCGs [28 0 R 29 0 R 30 0 R 31 0 R 32 0 R 33 0 R 34 0 R 35 0 R] >> / Страницы 3 0 R / Тип / Каталог >> эндобдж 2 0 obj > поток application / pdf
 сделал: 549e06e6-1cec-5845-a362-1c4050bede63uuid: 5D20892493BFDB11914A8590D31508C8proof: pdfuuid: b6e5c5e5-8359-499f-980d-61e7d2e4698dxmp.did: beb75929-8ea9-6142-93e3-dc2707a93accuuid: 5D20892493BFDB11914A8590D31508C8default
сделал: 549e06e6-1cec-5845-a362-1c4050bede63uuid: 5D20892493BFDB11914A8590D31508C8proof: pdfuuid: b6e5c5e5-8359-499f-980d-61e7d2e4698dxmp.did: beb75929-8ea9-6142-93e3-dc2707a93accuuid: 5D20892493BFDB11914A8590D31508C8default 422; ttfautohint (v1.8) Ложь
422; ttfautohint (v1.8) Ложь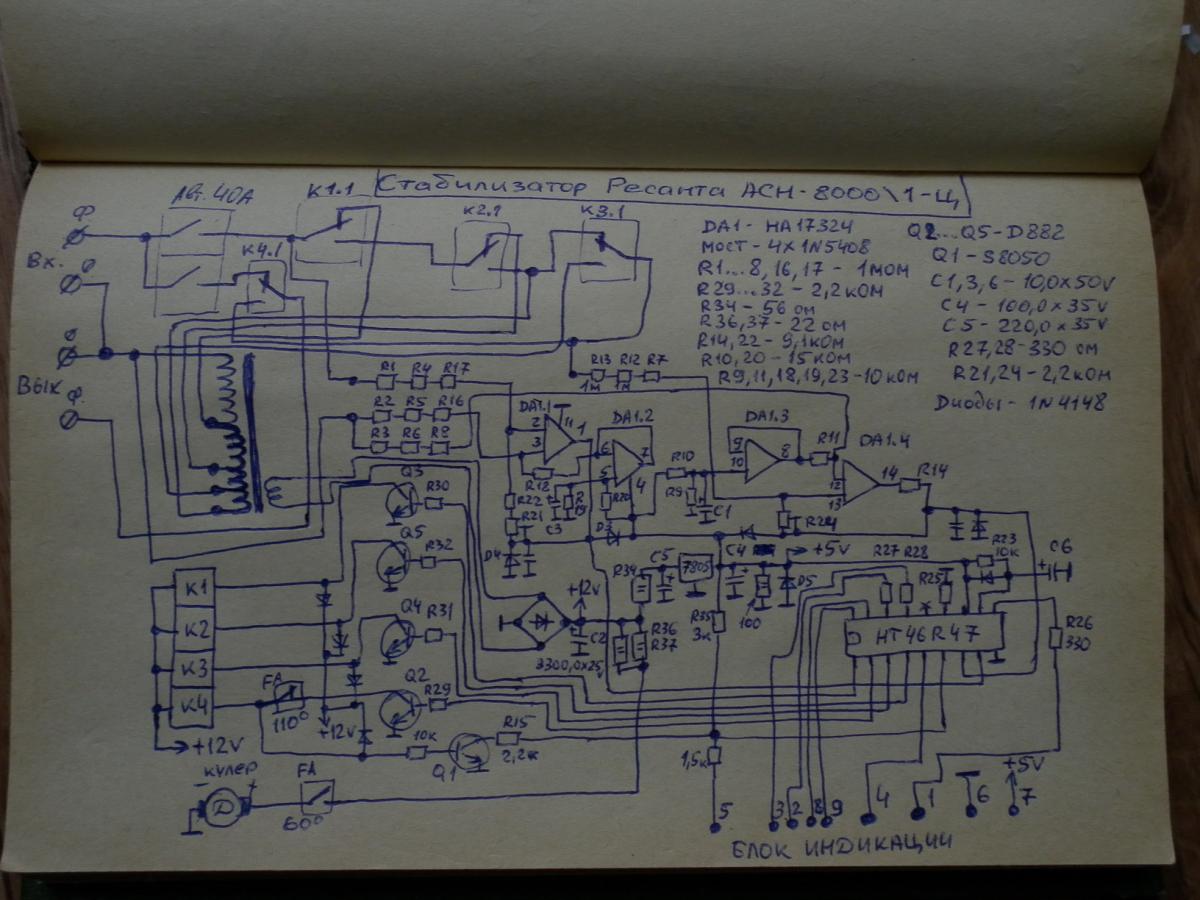 ttf
ttf 000000100.000000100.0000000.000000
000000100.000000100.0000000.000000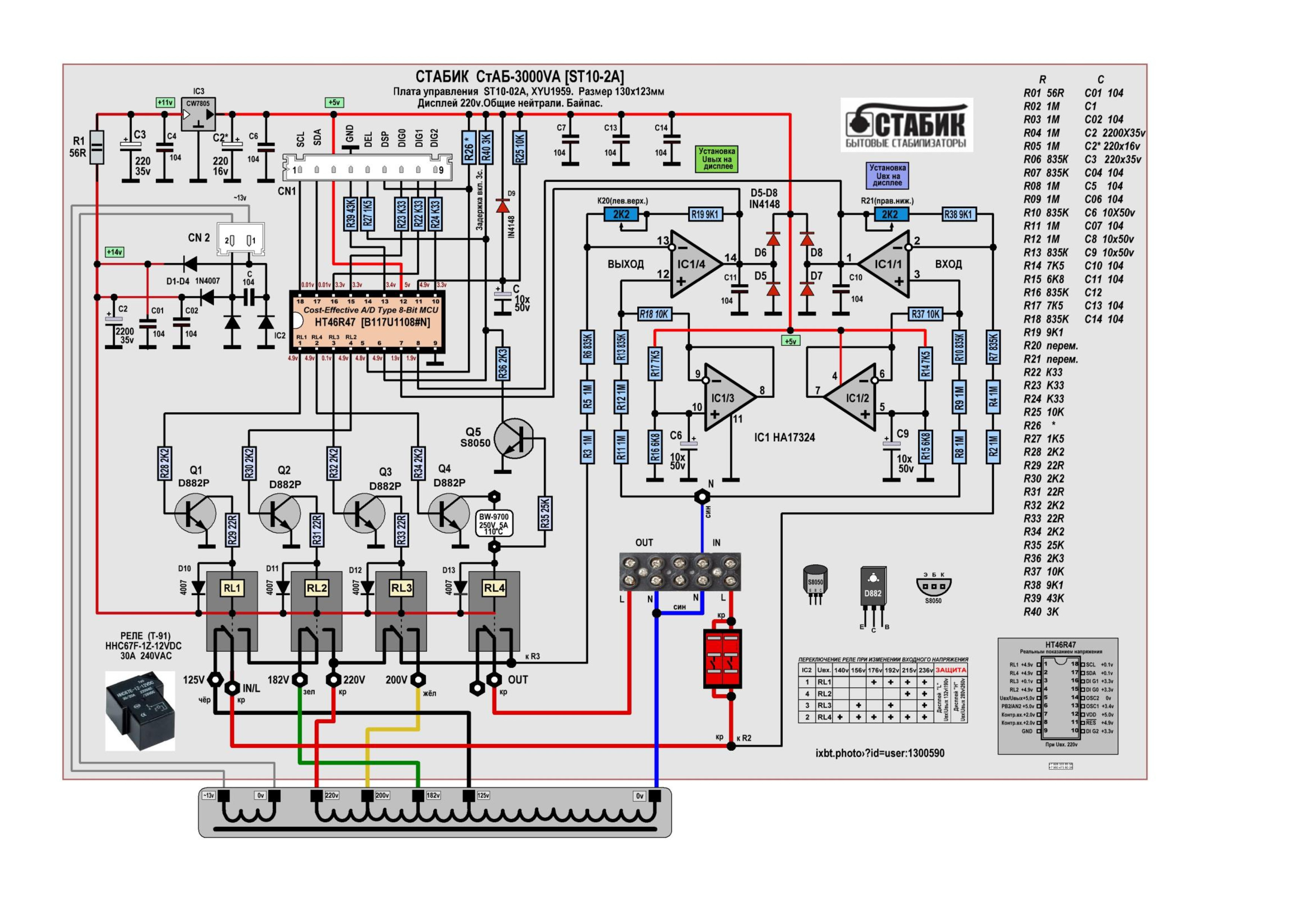 00000080.00000095.0000000.000000
00000080.00000095.0000000.000000 00000030.00000095.00000030.000000
00000030.00000095.00000030.000000 000000100.0000000.0000000.000000
000000100.0000000.0000000.000000 00000040.00000065.0000000.000000
00000040.00000065.0000000.000000 0000000.0000000.00000089.999400
0000000.0000000.00000089.999400 0000000.0000000.0000009.999100
0000000.0000000.0000009.999100 0031000.003100
0031000.003100 0 0,0 612,0 792,0] / Тип / Страница >>
эндобдж
78 0 объект
> / Ресурсы> / Шрифт> / ProcSet [/ PDF / Text] / Свойства >>> / Thumb 91 0 R / TrimBox [0.0 0.0 612.0 792.0] / Type / Page >>
эндобдж
79 0 объект
> / Ресурсы> / Шрифт> / ProcSet [/ PDF / Text] / Свойства >>> / Thumb 94 0 R / TrimBox [0.0 0.0 612.0 792.0] / Type / Page >>
эндобдж
80 0 объект
> / Ресурсы> / Шрифт> / ProcSet [/ PDF / Text] / Свойства >>> / Thumb 97 0 R / TrimBox [0.0 0.0 612.0 792.0] / Type / Page >>
эндобдж
81 0 объект
> / Ресурсы> / Шрифт> / ProcSet [/ PDF / Text] / Свойства >>> / Thumb 100 0 R / TrimBox [0.7u7wy6 @ oon> mn [m? N
0 0,0 612,0 792,0] / Тип / Страница >>
эндобдж
78 0 объект
> / Ресурсы> / Шрифт> / ProcSet [/ PDF / Text] / Свойства >>> / Thumb 91 0 R / TrimBox [0.0 0.0 612.0 792.0] / Type / Page >>
эндобдж
79 0 объект
> / Ресурсы> / Шрифт> / ProcSet [/ PDF / Text] / Свойства >>> / Thumb 94 0 R / TrimBox [0.0 0.0 612.0 792.0] / Type / Page >>
эндобдж
80 0 объект
> / Ресурсы> / Шрифт> / ProcSet [/ PDF / Text] / Свойства >>> / Thumb 97 0 R / TrimBox [0.0 0.0 612.0 792.0] / Type / Page >>
эндобдж
81 0 объект
> / Ресурсы> / Шрифт> / ProcSet [/ PDF / Text] / Свойства >>> / Thumb 100 0 R / TrimBox [0.7u7wy6 @ oon> mn [m? NДиссоциация между метаболизмом глюкозы и оксигенацией крови в сети человеческого режима по умолчанию, выявленная одновременным ПЭТ-фМРТ
% PDF-1.7 % 1 0 объект > эндобдж 4 0 obj > поток 2021-06-30T03: 08: 30 + 05: 30Arbortext Advanced Print Publisher 9.1.510 / W Unicode2021-11-07T15: 17: 38-08: 002021-11-07T15: 17: 38-08: 00 Acrobat Distiller 10.0.0 (Windows) application / pdf
 il} c۶t5e8 ~ Z% # ‘7n / ep 罻
9G} h’yb1: Աɘ] XJ -}; s
.mw; mXA; jq E5ήpM [ܗ EyK9FX ݾ 5 Ii_ ~ Ex {۬] Ƨ (hl
~ I o \ [% / QF_Y1gLY $ sT:> xI & GR2 / ly.s ݞ
il} c۶t5e8 ~ Z% # ‘7n / ep 罻
9G} h’yb1: Աɘ] XJ -}; s
.mw; mXA; jq E5ήpM [ܗ EyK9FX ݾ 5 Ii_ ~ Ex {۬] Ƨ (hl
~ I o \ [% / QF_Y1gLY $ sT:> xI & GR2 / ly.s ݞЭмоции, телесные чувства и мысли разделяются распределенными нейронными сетями
Аннотация
Ученые традиционно предполагали, что разные виды психических состояний (например, страх , отвращение, любовь, память, планирование, концентрация и т. д.) соответствуют различным психологическим способностям, которые имеют в мозгу корреляты, зависящие от предметной области. Тем не менее, растущее количество свидетельств указывает на конструкционистскую гипотезу о том, что психические состояния возникают из комбинации общих психологических процессов, которые отображаются в крупномасштабных распределенных сетях мозга.В этой статье мы сообщаем о новом исследовании, тестирующем конструкционистскую модель разума, в которой участники генерировали три типа психических состояний (эмоции, телесные чувства или мысли), в то время как мы измеряли активность в крупномасштабных распределенных сетях мозга с помощью фМРТ. Мы исследовали сходство и различия в паттернах сетевой активности в этих трех классах психических состояний. В соответствии с конструкционистской гипотезой комбинация крупномасштабных распределенных сетей способствовала эмоциям, мыслям и телесным ощущениям, хотя эти психические состояния различались по относительному вкладу этих сетей.Обсуждаются последствия для конструкционистской функциональной архитектуры различных психических состояний.
Мы исследовали сходство и различия в паттернах сетевой активности в этих трех классах психических состояний. В соответствии с конструкционистской гипотезой комбинация крупномасштабных распределенных сетей способствовала эмоциям, мыслям и телесным ощущениям, хотя эти психические состояния различались по относительному вкладу этих сетей.Обсуждаются последствия для конструкционистской функциональной архитектуры различных психических состояний.
Ключевые слова: конструкционист, эмоция, мысль, телесные чувства, внутренние сети
1. Введение
В каждый момент бодрствования человеческий разум состоит из множества психических состояний. Эти психические состояния обычно называют в терминах здравого смысла, таких как эмоции (например, страх, отвращение, любовь), познания (например, восстановление воспоминаний, планирование будущего, концентрация на задаче), восприятие (например, восприятие).ж., восприятие лица, восприятие цвета, восприятие звука) и т. д. С самого начала психологической науки исследователи предполагали, что каждое из этих слов относится к отдельной и отличной категории умственных способностей или «способности» (Lindquist & Barrett, в обзоре; Uttal, 2001). Соответственно, более века ученые искали физические корреляты этих ментальных категорий – в поведении, периферической физиологии и, в последнее время, в функционирующем мозге. Например, когнитивные нейробиологи попытались идентифицировать единую нейронную основу страха (например,g., Whalen et al., 1998), отвращение (например, Wicker et al., 2003), любовь (например, Bartels & Zeki, 2000), самость (например, Northoff & Bermpohl, 2004), эпизодическая память (Rugg , Otten, & Henson, 2002), семантическая память (например, Grossman et al., 2002), рабочая память (например, D’Esposito et al., 1998), восприятие лица (например, Kanwisher, McDermott, & Chun, 1997 ) и так далее. Двадцать лет исследований нейровизуализации, однако, показывают, что мозг не принимает во внимание категории психологии преподавателей (Barrett, 2009b; Duncan & Barrett, 2007; Gonsalves & Cohen, 2010; Lindquist & Barrett, на рассмотрении; Lindquist, Wager, Kober, Bliss , & Barrett, в печати; Pessoa, 2008; Poldrack, 2010; Uttal, 2001).
С самого начала психологической науки исследователи предполагали, что каждое из этих слов относится к отдельной и отличной категории умственных способностей или «способности» (Lindquist & Barrett, в обзоре; Uttal, 2001). Соответственно, более века ученые искали физические корреляты этих ментальных категорий – в поведении, периферической физиологии и, в последнее время, в функционирующем мозге. Например, когнитивные нейробиологи попытались идентифицировать единую нейронную основу страха (например,g., Whalen et al., 1998), отвращение (например, Wicker et al., 2003), любовь (например, Bartels & Zeki, 2000), самость (например, Northoff & Bermpohl, 2004), эпизодическая память (Rugg , Otten, & Henson, 2002), семантическая память (например, Grossman et al., 2002), рабочая память (например, D’Esposito et al., 1998), восприятие лица (например, Kanwisher, McDermott, & Chun, 1997 ) и так далее. Двадцать лет исследований нейровизуализации, однако, показывают, что мозг не принимает во внимание категории психологии преподавателей (Barrett, 2009b; Duncan & Barrett, 2007; Gonsalves & Cohen, 2010; Lindquist & Barrett, на рассмотрении; Lindquist, Wager, Kober, Bliss , & Barrett, в печати; Pessoa, 2008; Poldrack, 2010; Uttal, 2001).
Вместо того, чтобы выявить специфические для каждой умственной способности области мозга, зависящие от предметной области, растущее количество свидетельств указывает на гипотезу о том, что различные психические состояния возникают из комбинации общих психологических процессов или «ингредиентов», которые соответствуют крупномасштабным распределенным сетям. в ассоциативных областях мозга (см., Barrett, 2009b; Barrett, 2011). В психологии существует теоретическая традиция выдвигать гипотезы о том, что психические состояния возникают в результате комбинации более фундаментальных, общих психологических процессов, – это известно как конструкционистский подход.На протяжении прошлого столетия конструкционистский подход был наиболее популярен в моделях эмоций (например, Barrett, 2006, в печати; Harlow & Stagner, 1932; Lindquist, Wager, Kober, et al., В печати; Russell, 2003; Schachter). & Singer, 1962; обзор см. В Gendron & Barrett, 2009), хотя его корни можно найти в самых ранних работах по психологии (Dewey, 1895; James, 1890; Wundt, 1897/1998). Суть конструкционистского подхода состоит в том, что в каждый момент бодрствования мозг конструирует психические состояния, такие как эмоции, состояния тела и мысли, создавая ситуативные концептуализации (Barrett, 2006; Barsalou, 2009), которые объединяют три источника стимуляции. : сенсорная стимуляция из внешнего мира (экстероцептивный сенсорный массив света, вибраций, химических веществ и т. д.), сенсорные сигналы изнутри тела (соматовисцеральная стимуляция, также называемая интероцептивным сенсорным массивом или «внутренней средой») и предшествующий опыт (также называемый памятью или категориальным знанием, которое мозг частично делает доступным за счет реактивации сенсорных и моторных нейронов).Эти три источника – ощущения от мира, ощущения от тела и предшествующий опыт – доступны постоянно, и сети мозга, которые их обрабатывают, можно рассматривать как часть основных ингредиентов, формирующих всю психическую жизнь. Предполагается, что различные «рецепты» (комбинация и взвешивание ингредиентов) производят бесчисленное количество ментальных событий, которые люди дают здравым смыслом (например, «эмоции», «познания» и «восприятие»).
Суть конструкционистского подхода состоит в том, что в каждый момент бодрствования мозг конструирует психические состояния, такие как эмоции, состояния тела и мысли, создавая ситуативные концептуализации (Barrett, 2006; Barsalou, 2009), которые объединяют три источника стимуляции. : сенсорная стимуляция из внешнего мира (экстероцептивный сенсорный массив света, вибраций, химических веществ и т. д.), сенсорные сигналы изнутри тела (соматовисцеральная стимуляция, также называемая интероцептивным сенсорным массивом или «внутренней средой») и предшествующий опыт (также называемый памятью или категориальным знанием, которое мозг частично делает доступным за счет реактивации сенсорных и моторных нейронов).Эти три источника – ощущения от мира, ощущения от тела и предшествующий опыт – доступны постоянно, и сети мозга, которые их обрабатывают, можно рассматривать как часть основных ингредиентов, формирующих всю психическую жизнь. Предполагается, что различные «рецепты» (комбинация и взвешивание ингредиентов) производят бесчисленное количество ментальных событий, которые люди дают здравым смыслом (например, «эмоции», «познания» и «восприятие»). С этой точки зрения ментальные категории, такие как эмоции, познания и восприятия, заполнены разнообразным набором примеров, которые являются событиями, которые необходимо объяснить, а не конкретными причинными процессами, связанными с конкретными областями или сетями мозга.
С этой точки зрения ментальные категории, такие как эмоции, познания и восприятия, заполнены разнообразным набором примеров, которые являются событиями, которые необходимо объяснить, а не конкретными причинными процессами, связанными с конкретными областями или сетями мозга.
Есть три направления работы, которые поддерживают конструктивистскую функциональную архитектуру ментальных состояний. Во-первых, в литературе по нейровизуализации растет понимание того, что одни и те же сети усиливают активацию в самых разных областях психологических задач. Например, «сеть по умолчанию», включающая области медиальной префронтальной коры, медиальной височной доли и задней поясной коры, имеет повышенную активацию во время эмоций (например, Lindquist, Wager, Kober, et al., в печати), регулирование эмоций (например, Wager, Davidson, Hughes, Lindquist, & Ochsner, 2008), репрезентация себя (например, Kelley et al., 2002), приписывание психического состояния другим (например, Mitchell, Banaji , & Macrae, 2005), моральное рассуждение (например, Young, Scholz, & Saxe, в печати), эпизодическая память и прогнозирование (например, Addis, Wong, & Schacter, 2007), семантическая обработка (например, Binder, Desai, Graves , & Conant, 2009), и даже контекстно-зависимое визуальное восприятие (Bar et al. , 2006).«Сеть значимости», включающая кору островка и переднюю срединную часть коры головного мозга, имеет повышенную активность во время эмоций (например, Lindquist, Wager, Kober, et al., В печати), боли (например, Lamm, Decety, & Singer, 2010). тревога (например, Seeley et al., 2007), внимание, язык (см. Nelson et al., 2010) и восприятие времени (см. Craig, 2009). Даже сенсорные области мозга, которые когда-то считались унимодальными и специфичными для домена (например, первичная слуховая и зрительная кора), реагируют на другие сенсорные домены (например,g. слуховые нейроны проявляют повышенную активность при предъявлении зрительных стимулов; Бизли и Кинг, 2008; зрительные нейроны проявляют повышенную активность при предъявлении слуховых раздражителей; Cate et al., 2009). Эти результаты показывают, что отсутствие поддержки факультетской психологии – это не просто артефакт плохого пространственного или временного разрешения в методах нейровизуализации.
, 2006).«Сеть значимости», включающая кору островка и переднюю срединную часть коры головного мозга, имеет повышенную активность во время эмоций (например, Lindquist, Wager, Kober, et al., В печати), боли (например, Lamm, Decety, & Singer, 2010). тревога (например, Seeley et al., 2007), внимание, язык (см. Nelson et al., 2010) и восприятие времени (см. Craig, 2009). Даже сенсорные области мозга, которые когда-то считались унимодальными и специфичными для домена (например, первичная слуховая и зрительная кора), реагируют на другие сенсорные домены (например,g. слуховые нейроны проявляют повышенную активность при предъявлении зрительных стимулов; Бизли и Кинг, 2008; зрительные нейроны проявляют повышенную активность при предъявлении слуховых раздражителей; Cate et al., 2009). Эти результаты показывают, что отсутствие поддержки факультетской психологии – это не просто артефакт плохого пространственного или временного разрешения в методах нейровизуализации.
Фактически, метаанализы, обобщающие литературу по нейровизуализации по ментальным категориям, таким как эмоции, личность, память и т. Д.подтверждают, что области мозга демонстрируют небольшую психологическую специфичность (по крайней мере, для этих категорий или для их подчиненных категорий, таких как гнев, страх, отвращение, автобиографическое Я, обработка самореференций, автобиографическая память, семантическая память и т. д.). Например, наш метааналитический проект по эмоциям продемонстрировал, что миндалевидное тело (ранее считавшееся специфически связанным со страхом), передняя островковая часть (AI) (ранее считавшаяся связанной с отвращением), передняя средняя часть коры головного мозга (aMCC) и орбитофронтальная кора головного мозга. (OFC) (ранее считалось, что это связано с грустью и гневом соответственно), каждая из которых демонстрировала повышенную активность в переживании и восприятии множества различных эмоций, что указывает на то, что повышенная активность в этих областях не является специфической для какой-либо одной категории эмоций (Lindquist, Wager , Кобер и др., под давлением). Кроме того, во время переживания и восприятия эмоций повышалась активность в областях, обычно связанных с автобиографической памятью и поиском, языком и семантикой, а также исполнительным контролем (Barrett, Mesquita, Ochsner, & Gross, 2007; Kober, et al.
Д.подтверждают, что области мозга демонстрируют небольшую психологическую специфичность (по крайней мере, для этих категорий или для их подчиненных категорий, таких как гнев, страх, отвращение, автобиографическое Я, обработка самореференций, автобиографическая память, семантическая память и т. д.). Например, наш метааналитический проект по эмоциям продемонстрировал, что миндалевидное тело (ранее считавшееся специфически связанным со страхом), передняя островковая часть (AI) (ранее считавшаяся связанной с отвращением), передняя средняя часть коры головного мозга (aMCC) и орбитофронтальная кора головного мозга. (OFC) (ранее считалось, что это связано с грустью и гневом соответственно), каждая из которых демонстрировала повышенную активность в переживании и восприятии множества различных эмоций, что указывает на то, что повышенная активность в этих областях не является специфической для какой-либо одной категории эмоций (Lindquist, Wager , Кобер и др., под давлением). Кроме того, во время переживания и восприятия эмоций повышалась активность в областях, обычно связанных с автобиографической памятью и поиском, языком и семантикой, а также исполнительным контролем (Barrett, Mesquita, Ochsner, & Gross, 2007; Kober, et al. , 2008). ; Lindquist, Wager, Kober, et al., В печати). Мета-анализы, оценивающие нейронные корреляты других психических состояний, демонстрируют аналогичную точку зрения. Например, один метаанализ показал, что тот же набор срединных областей коры, которые составляют «сеть по умолчанию» (включая гиппокамп, медиальную префронтальную кору и заднюю часть поясной извилины), демонстрирует повышенную активность памяти, перспективу на будущее, теорию разума. , спонтанное мышление и пространственная навигация (Spreng, Mar, & Kim, 2009).Другой недавний метаанализ демонстрирует, что аналогичный набор регионов в дорсальной префронтальной и теменной корках задействован в рабочей памяти, выборе ответа, торможении ответа, переключении задач и когнитивном контроле (Lenartowicz, Kalar, Congdon, & Poldrack, 2010).
, 2008). ; Lindquist, Wager, Kober, et al., В печати). Мета-анализы, оценивающие нейронные корреляты других психических состояний, демонстрируют аналогичную точку зрения. Например, один метаанализ показал, что тот же набор срединных областей коры, которые составляют «сеть по умолчанию» (включая гиппокамп, медиальную префронтальную кору и заднюю часть поясной извилины), демонстрирует повышенную активность памяти, перспективу на будущее, теорию разума. , спонтанное мышление и пространственная навигация (Spreng, Mar, & Kim, 2009).Другой недавний метаанализ демонстрирует, что аналогичный набор регионов в дорсальной префронтальной и теменной корках задействован в рабочей памяти, выборе ответа, торможении ответа, переключении задач и когнитивном контроле (Lenartowicz, Kalar, Congdon, & Poldrack, 2010).
Вторая линия доказательств, подтверждающих жизнеспособность конструкционистского подхода к сознанию, исходит из исследований электростимуляции и повреждений. Электрическая стимуляция одного и того же участка человеческого мозга вызывает разные психические состояния в разных случаях (Halgren, Walter, Cherlow, & Crandall, 1978; Sem-Jacobson, 1968; Valenstein, 1974).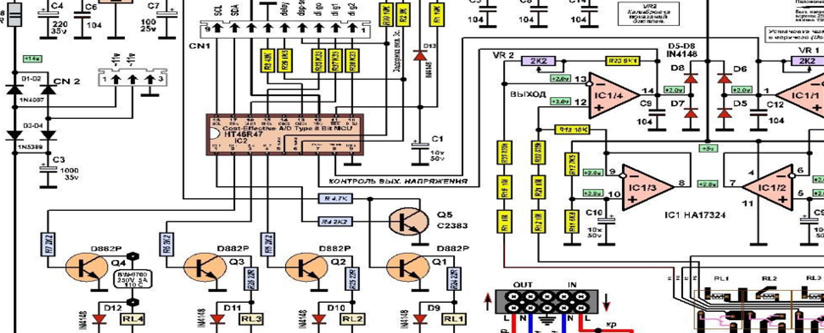 Даже исследования человеческих повреждений согласуются с идеей, что области мозга не являются специфичными для какого-либо одного психического состояния. Например, речевое расстройство, называемое афазией Брока, вызывается поражениями, выходящими за пределы области Брока, области мозга, которая, как считается, отвечает за производство речи (Mohr et al., 1978). В качестве другого примера, поражения миндалины не связаны конкретно с дефицитом обработки, связанной со страхом. Пациент с двусторонним поражением миндалины (т. Е. СМ) испытывает трудности с восприятием страха на лицах окружающих (например,g., Adolphs, Tranel, Damasio, & Damasio, 1994; Адольфс, Транель, Дамасио и Дамасио, 1995; Adolphs et al., 1999), но более поздние результаты показывают, что пациентка SM способна воспринимать страх, когда ее внимание явно направлено на глаза лица (Adolphs et al., 2005) или при просмотре карикатур на пугающие позы тела ( Аткинсон, Хеберлейн и Адольфс, 2007). Эти результаты предполагают, что миндалевидное тело может играть более общую роль в привлечении внимания к новым или мотивационно релевантным стимулам, которые способствуют возникновению страха, но что миндалевидное тело не является специфическим для страха (обсуждения см.
Даже исследования человеческих повреждений согласуются с идеей, что области мозга не являются специфичными для какого-либо одного психического состояния. Например, речевое расстройство, называемое афазией Брока, вызывается поражениями, выходящими за пределы области Брока, области мозга, которая, как считается, отвечает за производство речи (Mohr et al., 1978). В качестве другого примера, поражения миндалины не связаны конкретно с дефицитом обработки, связанной со страхом. Пациент с двусторонним поражением миндалины (т. Е. СМ) испытывает трудности с восприятием страха на лицах окружающих (например,g., Adolphs, Tranel, Damasio, & Damasio, 1994; Адольфс, Транель, Дамасио и Дамасио, 1995; Adolphs et al., 1999), но более поздние результаты показывают, что пациентка SM способна воспринимать страх, когда ее внимание явно направлено на глаза лица (Adolphs et al., 2005) или при просмотре карикатур на пугающие позы тела ( Аткинсон, Хеберлейн и Адольфс, 2007). Эти результаты предполагают, что миндалевидное тело может играть более общую роль в привлечении внимания к новым или мотивационно релевантным стимулам, которые способствуют возникновению страха, но что миндалевидное тело не является специфическим для страха (обсуждения см. В Cunningham & Brosch, в печати; Lindquist, Wager , Кобер и др., под давлением).
В Cunningham & Brosch, в печати; Lindquist, Wager , Кобер и др., под давлением).
Наконец, развивающаяся наука о «внутренних сетях» согласуется с идеей о том, что функциональная архитектура мозга содержит сети, которые соответствуют общим психологическим процессам предметной области, а не конкретным категориям психических состояний. Путем корреляции низкочастотных колебаний сигнала, зависящего от уровня оксигенации крови (ЖИРНЫЙ), в гемодинамической реакции вокселов, когда мозг находится «в состоянии покоя» (то есть, когда он не исследуется внешним стимулом или не выполняет направленную задачу), можно идентифицировать крупномасштабные распределенные сети, которые охватывают лобную, поясную, теменную, височную и затылочную коры.Эти сети хорошо воспроизводятся в исследованиях, в которых используются разные статистические методы, и наблюдаются как с помощью методов на основе семян (например, Vincent, Kahn, Snyder, Raichle, & Buckner, 2008), так и других многомерных методов (например, анализ независимых компонентов; ICA; Smith et al.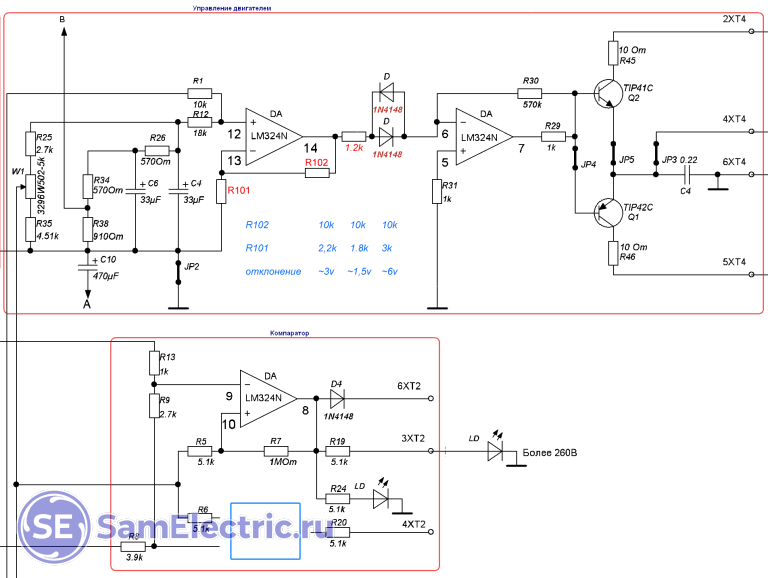 , 2009) и кластерный анализ (Yeo et al., 2011). Эти внутренние сети ограничены анатомическими связями (Buckner, 2010; Deco, Jirsa, & McIntosh, 2010; Fox & Raichle, 2007; Vincent, et al., 2008), поэтому они, кажется, раскрывают нечто фундаментальное о функциональной организации. к человеческому мозгу.Учитывая, что на внутреннюю активность приходится значительная часть метаболического бюджета мозга (Raichle & Minton, 2006), вполне возможно, что эти сети могут быть основными психологическими «ингредиентами» разума. Хотя к настоящему времени определен ряд внутренних сетей, ни одна из них, похоже, не соответствует той активности мозга, которая соответствует категориям в подходе факультетской психологии (т. Е., Похоже, не существует единой сети для «гнева» или даже «эмоции» против . «Познание»).
, 2009) и кластерный анализ (Yeo et al., 2011). Эти внутренние сети ограничены анатомическими связями (Buckner, 2010; Deco, Jirsa, & McIntosh, 2010; Fox & Raichle, 2007; Vincent, et al., 2008), поэтому они, кажется, раскрывают нечто фундаментальное о функциональной организации. к человеческому мозгу.Учитывая, что на внутреннюю активность приходится значительная часть метаболического бюджета мозга (Raichle & Minton, 2006), вполне возможно, что эти сети могут быть основными психологическими «ингредиентами» разума. Хотя к настоящему времени определен ряд внутренних сетей, ни одна из них, похоже, не соответствует той активности мозга, которая соответствует категориям в подходе факультетской психологии (т. Е., Похоже, не существует единой сети для «гнева» или даже «эмоции» против . «Познание»).
1.1 Настоящее исследование
В этой статье мы сообщаем о новом исследовании, тестирующем конструкционистскую модель разума, в котором мы измеряли активность в крупномасштабных распределенных сетях мозга с помощью фМРТ, когда участники генерировали три типа психических состояний (эмоции, телесные ощущения). , или мысли).Затем мы исследовали сходство и различия в паттернах сетевой активности в трех психических состояниях. В нашем эксперименте участники познакомились с новой техникой погружения в сценарий (Wilson-Mendenhall, Barrett, Simmons, & Barsalou, 2011), которая вызывает ментальные события, происходящие в повседневной жизни, что позволяет нам изучать уникальные субъективные переживания. человека (см. также Frith, 2007). Психология часто предполагает, что психические состояния возникают только тогда, когда человека исследуют внешние раздражители (на основе старой модели психики, заимствованной из физиологии в 19 веке; Danziger, 1997).Тем не менее, психические состояния не подчиняются этой классической модели «стимул-реакция» большую часть времени – людям не нужен стимул в физическом мире, чтобы иметь богатые и субъективно сильные эмоции, чувства или мысли (например, Killingsworth & Gilbert, 2010). . Мы попытались отдать должное этой особенности ментальной жизни, используя технику погружения в сценарий как экологически обоснованный метод, который направляет ментальное содержание, сохраняя при этом некоторую безудержность субъективного ментального опыта.
, или мысли).Затем мы исследовали сходство и различия в паттернах сетевой активности в трех психических состояниях. В нашем эксперименте участники познакомились с новой техникой погружения в сценарий (Wilson-Mendenhall, Barrett, Simmons, & Barsalou, 2011), которая вызывает ментальные события, происходящие в повседневной жизни, что позволяет нам изучать уникальные субъективные переживания. человека (см. также Frith, 2007). Психология часто предполагает, что психические состояния возникают только тогда, когда человека исследуют внешние раздражители (на основе старой модели психики, заимствованной из физиологии в 19 веке; Danziger, 1997).Тем не менее, психические состояния не подчиняются этой классической модели «стимул-реакция» большую часть времени – людям не нужен стимул в физическом мире, чтобы иметь богатые и субъективно сильные эмоции, чувства или мысли (например, Killingsworth & Gilbert, 2010). . Мы попытались отдать должное этой особенности ментальной жизни, используя технику погружения в сценарий как экологически обоснованный метод, который направляет ментальное содержание, сохраняя при этом некоторую безудержность субъективного ментального опыта.
Участники услышали насыщенные сенсорными ощущениями яркие сценарии неприятных ситуаций, и их попросили создать ситуативную концептуализацию, в ходе которой каждая ситуация переживалась как состояние тела (например,g., сердцебиение, прикосновение предмета к коже, взгляды, запахи, неприятные ощущения), эмоция (например, страх, гнев) или мысль (например, план, размышление). В начале каждого испытания участников подбирали по типу психического состояния, которое нужно было построить на этом испытании. Следуя подсказке, участники слушали сценарий по мере их построения, а затем подробно описывали состояние тела, эмоции или мысли. Этот метод визуализации был смоделирован после Аддис и его коллеги (2007), которые просили участников «конструировать», а затем «развивать» автобиографические воспоминания.Мы отдельно проанализировали этапы погружения в сценарий и этапы построения + разработки каждого испытания как два события (погружение в сценарий , опыт ).
Взяв сетевую модель разума в качестве исходного предположения, мы выдвинули гипотезу, что ментальные состояния были созданы в результате взаимодействия сетей (Fuster, 2006; Goldman-Rakic, 1988; McIntosh, 2000; Mesulam, 1998; также см. Bullmore). & Sporns, 2009), где психологическая функция набора областей мозга существует в функциональном взаимодействии этих областей.В частности, мы сосредоточились на семи внутренних сетях, недавно идентифицированных Йео и его коллегами (2011); эти сети были получены из самой большой выборки участников (N = 1000) в любом исследовании внутренней функциональной связи на сегодняшний день, а также воспроизводят сети, указанные в других опубликованных отчетах (например, Fox et al., 2005; Seeley, et al., 2007; Смит и др. 2009; Винсент и др., 2008). перечисляет области мозга, которые, как было установлено, составляют каждую сеть в исследованиях, а также ключевые документы, которые способствуют функциональному пониманию каждой сети.
Bullmore). & Sporns, 2009), где психологическая функция набора областей мозга существует в функциональном взаимодействии этих областей.В частности, мы сосредоточились на семи внутренних сетях, недавно идентифицированных Йео и его коллегами (2011); эти сети были получены из самой большой выборки участников (N = 1000) в любом исследовании внутренней функциональной связи на сегодняшний день, а также воспроизводят сети, указанные в других опубликованных отчетах (например, Fox et al., 2005; Seeley, et al., 2007; Смит и др. 2009; Винсент и др., 2008). перечисляет области мозга, которые, как было установлено, составляют каждую сеть в исследованиях, а также ключевые документы, которые способствуют функциональному пониманию каждой сети.
Таблица 1
Обзор семи представляющих интерес сетей
| Сеть | Области мозга | Области задач | Психологическое описание и гипотезы |
|---|---|---|---|
«лимбическая сеть» (Yeo et al. , 2011) , 2011) | двусторонняя передняя височная доля, медиальная височная доля, субгенуальная передняя поясная кора, медиальная и латеральная орбитофронтальная кора (хотя Yeo et al.Сеть | Поколение основного аффекта : задействование висцеромоторного контроля тела для создания основных аффективных чувств удовольствия или неудовольствия с некоторой степенью возбуждения. | |
| Гипотеза : Ощущение тела и эмоции> Мысль | |||
| «сеть значимости» (Seeley et al.  , 2007) или , 2007) или “вентральное внимание , сеть »(Yeo et al., 2011; Corbetta & Shulman, 2002) или “околопаточно-оперение сеть »(Винсент и др., 2008) | двусторонний передний срединный слой кора головного мозга (aMCC), передний островок (AI) и средний островок, лобная часть покрышки и части pars opercularis и височно-теменная соединение | внимание, направленное на тело : использование репрезентаций из тела для направления внимания и поведения.Этот ингредиент может использовать изменения гомеостатического состояния тела, чтобы сигнализировать о важных событиях в окружающей среде и регулировать поведенческие реакции. | |
| Гипотеза : Ощущение тела и эмоции> Мысль | |||
| «сеть по умолчанию» (Dosenbach et al.  , 2008; , 2008; Винсент и др., 2008; Yeo et al., 2011) | медиальная префронтальная кора, части pars triangularis, ретроспленальная зона , задняя поясная извилина кора / предклинье, медиальная височная доля (гиппокамп, энторинальная кора), двусторонняя верхняя височная борозда, части передней височной ATL), и угловая извилина |
| концептуализация : представление предшествующего опыта (т. е. памяти или категорийного знания) для придания значения ощущений от тела и мира в данный момент. |
| Гипотеза : Мысли и эмоции> Ощущение тела | |||
| «лобно-теменная сеть» (Dosenbach et al., 2008; Винсент и др., 2008; Yeo et al., 2011) или «исполнительный сеть управления »(Seeley et al., 2007) | двусторонняя дорсолатеральная префронтальная кора (dlPFC), нижняя теменная доля , нижняя теменная борозда и аспекты средней поясной извилины кора (mCC) | исполнительное внимание : модуляция активности других ингредиентов для создания единого поля сознания во время построения психического состояния (например, ).  g. выбор g. выбор некоторого концептуального содержания, когда значение состоит из ощущений, и запрещение другого содержания; выбор одних ощущений для сознательного осознания и подавления других). | |
| Никакой конкретной гипотезы не сформулировано | |||
| “дорсальное внимание , сеть »(Corbetta & Shulman, 2002; Yeo et al., 2011;) | двусторонние лобные поля глаза, дорсальная задняя теменная кора, веретенообразная извилина, область MT + | • нисходящий контроль зрительно-пространственного внимания (Corbetta et al., 2002) | зрительно-пространственное внимание : модулирование активности ингредиента для обработки визуального контента, в частности (например, выбор того, какое зрительное ощущение выбрано для осознанного осознания, и подавление других).  |
| Никакой конкретной гипотезы не сформулировано | |||
| «соматомоторная сеть» (Yeo et al.2011) | прецентральные и постцентральные гиры (сенсомоторная кора), Heschl’sgyrus (первичная слуховая кора) кора головного мозга, задний островок | Экстероцептивное сенсорное восприятие : представление слуховых и тактильных ощущений | |
| Никакой конкретной гипотезы не сформулировано | |||
«визуальная сеть» (Yeo et al. 2011) 2011) | затылочная доля | • зрение (Engel et al. 1994) | экстероцептивное сенсорное восприятие : представление зрительных ощущений |
| Никакой конкретной гипотезы не сформулировано | |||
 2009)
2009) Мы выдвинули гипотезу о том, »Поддерживает способность мозга генерировать и / или представлять соматовисцеральные изменения, которые воспринимаются как основной аффективный тон, который является общим для каждого психического состояния. Многие философы и психологи предполагали, что каждый момент психической жизни имеет какой-то аффективный аспект (например,g., Wundt, 1897), которое можно описать как сочетание гедонического удовольствия и неудовольствия с некоторой степенью возбуждения (Barrett & Bliss-Moreau, 2009; Russell & Barrett, 1999). С нашей конструкционистской точки зрения мы называем этот базовый психологический элемент «основным аффектом» (Russell, 2003). Хотя лимбическая сеть, описанная Yeo et al. (2011), ограничена относительно небольшой областью коры (охватывающей только вентромедиальную префронтальную кору и вентральные аспекты височной коры), некоторые подкорковые структуры также, вероятно, являются частью этой лимбической сети.Например, мы выдвигаем гипотезу о том, что ядра базальных ганглиев являются частью «лимбической» сети, потому что они участвуют в управлении поведением, требующим усилий (Salamone & Correa, 2002; Salamone, Correa, Farrar, & Mingote, 2007) и моторном контроле ( Grillner, Hellgren, Menard, Saitoh, & Wikstrom, 2005). Кроме того, центральное ядро миндалевидного тела и периакведуктальный серый средний мозг могут быть частью этой сети, поскольку они, соответственно, участвуют в выработке вегетативных ответов (обсуждение см. В Barrett, Mesquita, et al., 2007) и координации последовательных физиологических и поведенческих реакций (Bandler & Shipley, 1994; Van der Horst & Holstege, 1998).
С нашей конструкционистской точки зрения мы называем этот базовый психологический элемент «основным аффектом» (Russell, 2003). Хотя лимбическая сеть, описанная Yeo et al. (2011), ограничена относительно небольшой областью коры (охватывающей только вентромедиальную префронтальную кору и вентральные аспекты височной коры), некоторые подкорковые структуры также, вероятно, являются частью этой лимбической сети.Например, мы выдвигаем гипотезу о том, что ядра базальных ганглиев являются частью «лимбической» сети, потому что они участвуют в управлении поведением, требующим усилий (Salamone & Correa, 2002; Salamone, Correa, Farrar, & Mingote, 2007) и моторном контроле ( Grillner, Hellgren, Menard, Saitoh, & Wikstrom, 2005). Кроме того, центральное ядро миндалевидного тела и периакведуктальный серый средний мозг могут быть частью этой сети, поскольку они, соответственно, участвуют в выработке вегетативных ответов (обсуждение см. В Barrett, Mesquita, et al., 2007) и координации последовательных физиологических и поведенческих реакций (Bandler & Shipley, 1994; Van der Horst & Holstege, 1998). Важно отметить, что базальные ганглии, миндалина и периакведуктальный серый – все проецируются на вентромедиальную префронтальную кору (vmPFC), которая является одной из кортикальных областей в лимбической сети Yeo et al.
Важно отметить, что базальные ганглии, миндалина и периакведуктальный серый – все проецируются на вентромедиальную префронтальную кору (vmPFC), которая является одной из кортикальных областей в лимбической сети Yeo et al.
Мы предполагаем, что «сеть значимости» (именуемая «вентральным вниманием» Йео и др. 2011) использует представления аффекта для управления вниманием и поведением (см. Lindquist & Barrett, в обзоре; Medford & Critchley, 2010) .Важно отметить, что сеть значимости включает аспекты спинной передней части островковой коры и передней средней поясной извилины (aMCC), которые участвуют в исполнительном внимании (Corbetta, Kincade, & Shulman, 2002; Touroutoglou, Hollenbeck, Dickerson & Barrett, в печати). ) и интероцепции (Critchley, Elliott, Mathias, & Dolan, 2000; Critchley, Wiens, Rotshtein, Ohman, & Dolan, 2004), предполагая, что эта сеть является важным источником аффективного внимания в человеческом мозгу (Barrett & Bar, 2009 ; Дункан и Барретт, 2007).Сеть значимости также содержит аспекты вентральной передней островковой доли, которая участвует в переживании аффективных состояний (Touroutoglou, et al., В печати).
Мы предполагаем, что «сеть по умолчанию» способствует представлению или «моделированию» предыдущего опыта и извлечению категорийных знаний для создания ситуативных концептуализаций (т. Е. Придания значения соматовисцеральным изменениям в теле по отношению к непосредственному контексту) . Мы предполагаем, что эта сеть является ключом к процессу реактивации соответствующих распределенных областей мозга для поддержки категорийных знаний, воспоминаний и перспектив будущего путем управления сенсорными и моторными регионами.Задние аспекты сети по умолчанию (например, задняя поясная извилина, предклинье, гиппокамп) могут быть особенно вовлечены в интеграцию зрительно-пространственных аспектов знаний категории (Cavanna & Trimble, 2006), тогда как передние аспекты сети по умолчанию (например, mPFC) могут быть вовлечены в интеграцию аффективных, социальных и релевантных для себя аспектов категорийного знания (Gusnard et al. 2001).
«Лобно-теменная сеть» играет исполнительную роль, модулируя активность в других функциональных сетях (т.д., отдавая приоритет некоторой информации и запрещая другую информацию), чтобы помочь сконструировать пример психического состояния. «Сеть дорсального внимания» играет аналогичную роль, направляя внимание, в частности, на визуальную информацию. Мы предполагаем, что во время задачи по погружению в сценарий эти сети способствовали процессам исполнительного контроля, задействованным в выделении определенных типов информации в сознательном осознании, чтобы создать один тип психического состояния над другим. Например, хотя концептуализация важна для всех состояний, она особенно важна для переживаний эмоций и мыслей.Эти сети гарантируют, что психическое состояние воспринимается как единое (обсуждение см. В Lindquist, Wager, Bliss-Moreau, Kober, & Barrett, в печати).
Наконец, «зрительная» и «соматомоторная» сети вместе участвуют в представлении зрительных, проприоцептивных и слуховых ощущений. Мы называем их «экстероцептивными» ощущениями, потому что информация извне тела представлена в виде звуков, запахов, вкусов, проприоцепции и взглядов. Мы предполагаем, что эти ощущения важны при построении всех психических состояний.
включает наши гипотезы для участия каждой сети в телесных чувствах, эмоциях и мыслях. Во-первых, мы предположили, что телесные чувства, эмоции и мысли будут включать некоторую степень аффекта, концептуализации и исполнительного внимания ( Гипотеза 1 ). Следуя этой гипотезе, мы предсказали, что лимбическая сеть, сеть значимости, сеть по умолчанию и лобно-теменная сеть будут обычно задействованы в сочетании всех психических состояний. Во-вторых, мы предположили, что сравнение активности мозга в разных психических состояниях даст относительные различия в вкладе каждого ингредиента в каждый вид состояния (Гипотеза 2) .В частности, мы предсказали, что состояния тела и эмоции задействуют лимбические сети и сети значимости относительно больше, чем мысли (Гипотеза 2a) (см.). Мы также предсказали, что мысли и эмоции будут задействовать сеть по умолчанию относительно больше, чем состояния тела, поскольку мы полагали, что концептуализация будет играть большую роль в ментальных состояниях, где представление предшествующего опыта необходимо для придания значения телесным ощущениям в данный момент ( т. е. эмоции) или когда представление предыдущего опыта используется для руководства планами, ассоциациями и размышлениями о ситуации (т.э., мысль) (Гипотеза 2б) (см.). Наконец, у нас не было априорных предсказаний того, как лобно-теменная, дорсальная, соматомоторная и зрительная сети будут различаться в трех классах психических состояний.
2. Метод
2.1 Участники
Участниками были 21 взрослый праворукий, носитель английского языка (12 женщин, M , возраст = 26,42, SD , возраст = 5,72). Участники дали письменное информированное согласие в соответствии с наблюдательным советом организации Partners Health Care и получили за свое участие до 200 долларов США.Потенциальные участники указали, были ли у них в анамнезе нарушения обучаемости, психические заболевания, клаустрофобия, когнитивная дисфункция или злоупотребление алкоголем / наркотиками при телефонном скрининге, проведенном до включения в исследование. Участники, которые сообщили о любом из этих состояний, не были включены в исследование. Участники были также проверены на использование психоактивных или системных лекарств и на совместимость с МРТ во время этого первоначального телефонного скрининга. Одна участница была дисквалифицирована в день исследования, потому что этим утром она принимала лекарства от мигрени.Окончательная выборка состояла из 20 участников (11 женщин, M , возраст = 26,40, SD , возраст = 5,93). В день эксперимента участники также заполнили Торонтскую шкалу алекситимии из 20 пунктов (TAS-20) (Bagby, Taylor, & Parker, 1994) и несколько других показателей самооценки, которые не имеют отношения к гипотезам, обсуждаемым в этой статье. . Алекситимия – это черта, характеризующаяся внешним мышлением и трудностями в определении и обозначении собственных эмоций (Sifneos, 1973). Ни один из участников нашей выборки не набрал больше порогового значения для Alexithymia (отсечка = 61, M TAS = 38.57; SD TAS = 7,55). Это открытие исключало, что кто-либо из наших участников обладал чертами, которые могли бы затруднить выполнение нашей задачи по построению психического состояния.
2.2 Процедура
В настоящем исследовании использовался новый метод погружения в сценарий, разработанный в нашей лаборатории и ранее использовавшийся в исследованиях нейровизуализации (Wilson-Mendenhall, et al. 2011). В этом методе участникам предлагаются сценарии, которые описывают ситуации от первого лица, и их просят представить каждый сценарий так, как если бы они действительно были там.Сценарии содержат яркие сенсорные детали, чтобы участники могли представить ситуацию в мультимодальной манере, создавая впечатления с высоким уровнем субъективного реализма. Сценарии, использованные в этом исследовании, описывали физические ситуации, связанные с автомобилями, лодками, велосипедами, пешим туризмом, катанием на лыжах, едой, напитками, растениями и животными (см. Примеры). Отрицательные сценарии включают описание физического вреда, тогда как нейтральные сценарии описывают аналогичные физические действия, при которых вреда не было. Набор стимулов состоял как из полных сценариев, так и из сокращенных версий одних и тех же сценариев.Сжатые сценарии были сокращенными версиями полных сценариев, которые все еще отражали суть ситуации и были представлены во время сеанса сканирования из-за ограничений по времени (см. Примеры).
Таблица 2
| Полная версия | Полная версия |
| Вы едете домой после того, как не выпили всю ночь. Длинный отрезок дороги перед вами, кажется, продолжается вечно. Вы закрываете глаза на мгновение.Автомобиль начинает заносить. Вы просыпаетесь. Вы чувствуете скольжение рулевого колеса в ваших руках. | Вы едете по парку на велосипеде без шлема. Куда ни глянь, люди устраивают пикники и играют. Передняя шина попадает в выбоину. Вы плывете по рулю . Ваша голова врезается в бетонную поверхность . Вы можете почувствовать, как кровь стекает по вашему лицу. |
| Краткая версия | Краткая версия |
| Вы едете домой после того, как не выпили всю ночь.Вы на мгновение закрываете глаза, и машина начинает заносить. | Вы едете по парку на велосипеде без шлема. Ваша передняя шина попадает в выбоину, и вы плывете через руль . |
Эксперимент состоял из сеанса обучения вне сканера и сеанса тестирования внутри сканера (согласно Wilson-Mendenhall, et al., 2011). Оба произошли в один день. Во время сеанса обучения участники слушали сценарии, чтобы ознакомиться с полными сценариями и сжатыми сценариями, которые они позже услышат во время сеанса сканирования.Эта процедура использовалась для того, чтобы участники могли получить подробную информацию о полном сценарии при представлении сжатой версии в сканере. Во время тренинга участники также получили подробные инструкции по строительной задаче. Участникам сказали, что строительная задача была разработана для оценки мозговой основы различных умственных переживаний. Прежде чем они услышат сжатый сценарий, появится подсказка, которая проинструктирует их, какое ментальное состояние они должны создать в ответ на сценарий.В состоянии ТЕЛО участников просили создать ощущение тела в ответ на ситуацию, описанную в сценарии. Во время тренировки экспериментатор привел примеры различных внутренних соматовисцеральных ощущений и проинструктировал участников, как создавать и обращать внимание на эти ощущения в ответ на сценарий (подробные инструкции см. В дополнительных материалах). В состоянии ЭМОЦИИ участников просили создать определенную эмоцию в ответ на ситуацию, описанную в сценарии.Экспериментатор привел примеры нескольких конкретных эмоций (гнев, страх, печаль) и проинструктировал участника, как создавать и проявлять внимание к этим эмоциональным переживаниям в ответ на сценарий. В условии «МЫСЛЬ» участников просили создать мысль в ответ на ситуацию, описанную в сценарии. Экспериментатор объяснил, что мысль включает объективный анализ ситуации и рассуждения о том, что происходит, без культивирования каких-либо чувств или эмоций. Мы также включили условие ВОСПРИЯТИЯ, в котором участников просили создать сенсорное восприятие в ответ на ситуацию, описанную в сценарии.Экспериментатор проинструктировал участника обратить внимание на объекты в сцене и отношения между ними, не анализируя их, не испытывая эмоций или чувств. После того, как исследователь убедился, что участник понял инструкции задачи на этапе обучения, участники выполнили 8 практических испытаний, в которых они практиковались в построении всех четырех мысленных переживаний.
Во время сеанса сканирования участникам было представлено 36 негативных сценариев и 12 нейтральных сценариев.Каждое испытание начиналось с 2-секундной фазы сигнала , чтобы проинструктировать участников, какое психическое состояние они должны были создать в ответ на сценарий. Негативные сценарии были случайным образом объединены с тремя разными репликами, так что 12 переживались как состояния тела, 12 как эмоции и 12 как мысли. Участникам было предложено испытать 12 нейтральных сценариев в качестве восприятия, чтобы они не привыкли к негативным сценариям во многих презентациях. Данные испытаний восприятия не были включены в анализ.Задача была разбита на три 12,6-минутных прогона, каждый из которых случайным образом представлял 16 сценариев. Инструкции и сценарии были представлены через наушники; реплики и фиксирующий крест были представлены на экране, который был виден через зеркало, установленное на катушке для головы. Задача была представлена с помощью программного пакета «Презентация» (Neurobehavioral Systems, Inc.).
После фазы реплики участники слушали сокращенную версию сценария в течение 10 секунд (, фаза погружения сценария ).На этом этапе участников проинструктировали использовать сокращенную версию, чтобы «испытать сценарий с точки зрения конкретного психического состояния, с которым вы получили сигнал». После фазы погружения в сценарий участники вошли в фазу опыта , где они сконструировали и развили психическое состояние, чтобы испытать его как можно более интенсивно, не меняя его, не думая и не испытывая чего-либо еще. Участники нажимали кнопку, чтобы указать, когда психическое состояние было построено и что они начали разработку (см. Addis, et al., 2007). Фаза опыта (строительство + проработка) длилась 20 секунд. После того, как фаза опыта закончилась, участники оценили яркость своего опыта и степень, в которой они были успешны в создании заданного опыта. Рейтинги выставлялись по непрерывной шкале от 0 («совсем нет») до 10 («100%, полностью»). Интервал между стимулами был установлен на 4 секунды, в течение которых крестик фиксации отображался в центре экрана.
2.3 Детали изображений
Данные изображений были собраны в Центре биомедицинской визуализации Athinoula A. Martinos с использованием 3T Siemens Magnetom Trio и 12-канальной матричной катушки для головки. Участники были ограничены в движении головы за счет использования расширяющихся подушек из пеноматериала и носили наушники, безопасные для МРТ, через которые они слышали сценарии.
Каждый сеанс сканирования начинался с автоматического сканирования изображения и процедур подгонки. Затем были получены структурные T1-взвешенные мультиэхо-изображения MEMPRAGE (van der Kouwe, Benner, Salat, & Fischl, 2008) (TR = 2530 мс, TE1 = 1.64 мс, TE2 = 3,5 мс, TE3 = 5,36 мс, TE4 = 7,22 мс, угол поворота = 7 °, с однократным захватом с чередованием и размером вокселя 1,0 × 1,0 × 1,0 мм). После структурного прогона была получена 6-минутная функциональная последовательность состояния покоя, которая не будет обсуждаться далее в этой рукописи. Затем последовали три функциональных прогона, в каждом из которых было получено 378 функциональных объемов, взвешенных по T2 * (TR = 2000 мс, TE = 30 мс, угол поворота = 90 °, FOV = 200 мм, с получением чередующихся срезов и размером вокселя 3,1 × 3.1 × 4,0 мм). Общая продолжительность сеанса сканирования составила чуть более часа. После функциональных прогонов была получена 8-минутная последовательность изображений сенсора диффузора (DSI), которую мы не будем обсуждать далее.
2.4 Анализ данных
Функциональные и структурные данные были предварительно обработаны и проанализированы с помощью Freesurfer 5 (http://surfer.nmr.mgh.harvard.edu/). Структурная предварительная обработка следовала стандартному протоколу Freesurfer для реконструкции и парцелляции кортикальной поверхности и объема, включая автоматическое преобразование Талаираха, нормализацию интенсивности, удаление черепа, сегментацию белого вещества и регистрацию в сферическом атласе с использованием многомерного нелинейного алгоритма регистрации (Dale, Fischl , & Sereno, 1999; Fischl, Sereno, & Dale, 1999; Fischl, Sereno, Tootell, & Dale, 1999).Функциональная предварительная обработка включала коррекцию движения, пространственное сглаживание (ядро 5 мм) и регистрацию функциональных изображений на анатомическом сканировании.
После предварительной обработки отдельные временные ряды были смоделированы с использованием канонической функции гемодинамического ответа (гамма-функция, гемодинамическая задержка 2,25; дисперсия 1,25). Матрица дизайна включала регрессоры движения и полиномиальные регрессоры помех второго порядка. Временные точки с перемещением более 2 мм были исключены из анализа. Предварительный анализ показал, что этап опыта не показал каких-либо существенных различий между построением и разработкой, поэтому мы проанализировали его как единое 20-секундное событие (строительство + проработка).Мы смоделировали функцию гемодинамического ответа (HRF) для фазы сигнала, фазы погружения в сценарий, фазы опыта и фазы оценки отдельно. Впоследствии мы выполнили анализ случайных эффектов на уровне группы для расчета контрастных карт.
2.5 Кортикальная парцелляция
Чтобы идентифицировать семь внутренних сетей, представляющих интерес для нашего анализа, мы использовали поверхностные сетевые метки, представленные в Freesufer Йео и его коллегами (2011). Эти сети были получены на основе анализа данных о состоянии покоя у 1000 здоровых молодых людей.Йео и др. использовали кластерный подход для выявления сетей функционально связанных регионов коры головного мозга. Их 7-сетевое решение было очень согласованным в ходе анализа, при этом 97,4% вершин были назначены одной и той же сети через обнаружение и набор данных репликации. Разделение кортикальной поверхности на определенные регионы было основано на кортикальном атласе Desikan-Killiany (Desikan et al., 2006). В дополнение к поверхностному функциональному анализу мы также сообщаем о результатах внутри подкоркового объема с использованием подкорковых меток, предоставленных Freesurfer (Fischl et al., 2002).
2.5.1. Проверка гипотез
Чтобы исследовать гипотезу 1 о том, что все психические состояния (состояния тела, эмоции, мысли) будут включать некоторую степень представления текущего состояния тела, концептуализации и исполнительного внимания, мы оценили общее сетевое взаимодействие для эмоций, тела. чувства и мысли во время фазы переживания с анализом конъюнктуры. Мы сосредоточились конкретно на фазе опыта , потому что в отличие от фазы погружения сценария , на этой фазе не было внешнего стимула.Следовательно, соединение отражает паттерны активации, которые обычно задействуются, когда люди генерируют определенное ментальное содержание и сосредотачиваются на своем внутреннем мире без какой-либо внешней стимуляции. Анализ конъюнкции для фазы погружения в сценарий , который показывает обычную активацию во время слушания и погружения в сценарий в разных состояниях психического состояния, представлен в – в дополнительных материалах. Каждый анализ конъюнкции был реализован путем взятия минимума абсолютного значения в каждой вершине через фокус тела vs.фиксация, эмоция против фиксации и мысль против контрастов фиксации. Впоследствии мы определили кластеры в конъюнкции, которые охватывают не менее 100 непрерывных вершин, все из которых достигают порога значимости p <0,001. Мы также выполнили анализ конъюнкции объема, уделяя особое внимание подкорковым структурам, выявив кластеры, охватывающие не менее 20 непрерывных вокселей, все достигающие значимости p <0,0001 (см. В дополнительных материалах).
Таблица 3
Значимые кластеры для анализа сочетаний телесных чувств, эмоций и мыслей во время фазы опыта .
| Регион | Сеть парцелляция | Tal X | Tal Y | Tal Z | k | max | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| lh Superior frontal / SMA 902 | −24,8 | −1,3 | 41,4 | 7857 | 7.46 | |||||||||
| левый передний островок / vlPFC | S / D | −46,4 | 12,5 | 2,1 | 4682 | 5,96 | ||||||||
| правый верхний передний угол | 72322 90M | 7A32 / a 5,3 | 56,3 | 2957 | 5,96 | |||||||||
| левый прецентральный | SM | −33,5 | −19,4 | 38,8 | 1708 | 5,625 | 1708 | 5,625 | 5 −35.2−35,3 | 35,8 | 2482 | 5,47 | ||
| правая передняя островковая часть | S | 34,7 | 3 | 4,9 | 814 | 9,3 | 814 | нижняя часть | −21,6 | −57,3 | 56 | 656 | 5,28 | |
| левый супрамаргинальный / TPJ | S | −53,8 | −39,8 | 31.9 5 | ||||||||||
| правая часть оперкулярная | S | 43,3 | 10,2 | 5,6 | 568 | 4,76 | ||||||||
| лч lOFC | 9069 −1 9069 −1 902,79 11764,69 | |||||||||||||
| левая верхняя теменная | DA | −16,5 | −48,3 | 57,9 | 404 | 4,54 | ||||||||
| левосторонняя | передцентральная3 | 4,8 | 26,9 | 292 | 4,22 | |||||||||
| правая передняя часть | VA | 54,7 | 5,3 | 10,4 | 295 | передняя | 10,4 | 295 | DA2 902 32 | −9,1 | 51,8 | 581 | 4,08 | |
| правая часть треугольной формы | D | 49,2 | 27,5 | 0,4 | 220 | 3.66 | ||||||||
| л / ч dlPFC | FP | −41,1 | 35,1 | 17,5 | 486 | 3,62 | ||||||||
| л / ч dlPFC | ||||||||||||||
| 9069 FP2 | 902FP 902 902 902 3,18 | |||||||||||||
| правая затылочная доля * | V | 22,6 | −95,3 | 1,6 | 242 | −4,27 | ||||||||
Таблица 4 9020 Анализ подгруппы
и анализ контраста
| Область | Tal X | Tal Y | Tal Z | k | max | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Соединение | lh | ||||||||||
| очков опыта | lhpallidum / putamen | -43.6 | 10,7 | 0,4 | 1714 | 6,57 | |||||
| правый мозжечок | 29,7 | −58,2 | −18,1 | 279 | 279 | −57,2 | −36,7 | 96 | 5,29 | ||
| левый мозжечок | −43,6 | −54,6 | −23,3 | 39 | 5.05 | ||||||
| левый мозжечок | −29,7 | −50,7 | −23,5 | 47 | 4,98 | ||||||
| правый мозжечок 9069 −2 902,1 | 9069 −14,96 | ||||||||||
| Ощущение тела по сравнению с | lh thalamus / pallidum | −7,9 | 0 | 6,5 | 186 | 4,06 | |||||
| мысль | таламус | таламус | правый хвостик8 | −11,7 | 18,1 | 99 | 3,75 | ||||
| сценарий погружения | левый мозжечок | −37,6 | −54,7 | −25 | 55 | 3,54 | |||||
| Эмоции против мысли | правый мозжечок | 29,7 | -43,6 | -37,4 | 66 | 4,29 | |||||
| сценарий погружения | правый мозжечок | 13.8 | −44,7 | −20,5 | 22 | 4,00 | |||||
| средний мозжечок | 0 | −60,3 | −21,4 | 25 | 152 3,4 сети будут по-разному влиять на каждый вид психического состояния, мы сначала выполнили анализ области интересов (ROI), используя сетевые метки Yeo et al. Каждая из сетей Йео и др. Была преобразована в индивидуальную корковую поверхность каждого участника с использованием сферического пространства в качестве промежуточного пространства регистрации.Из полученных семи ROI, каждая из которых представляет интересующую сеть, мы извлекли бета-веса (процентное изменение сигнала) для каждого контраста, сравнивая психическое состояние с фиксацией (погружение в сценарий и опыт отдельно). Эти бета-веса были проанализированы с помощью однофакторного ( психического состояния, ) дисперсионного анализа с повторными измерениями с тремя уровнями (тело, эмоции, мысли). Если предположение о сферичности не соблюдалось, представляются скорректированные тесты Greenhouse-Geisser. Простые эффекты значимы при p <.05 представлены в тексте, когда основной эффект достиг значимости. В качестве еще одной проверки Гипотезы 2 мы провели контрасты между тремя категориями психических состояний по всей поверхности коры. Это позволило нам напрямую сравнивать психические состояния без сравнения с фиксацией. Сравнение с фиксацией могло ограничить шанс найти различия в вовлечении сети по умолчанию в анализе конъюнкции и анализе ROI (см. Stark & Squire, 2001).Кроме того, контрастирование всего мозга позволило нам проверить, отражают ли результаты ROI участие одного отдельного субрегиона в каждой сети или действительно ли результаты отражают распределенную сеть. Мы локализовали кластеры активности, используя поверхностные метки от Yeo et al. (2011) и поверхностный кортикальный атлас Desikan-Killiany (Desikan et al., 2006). Мы сравнили три категории психических состояний в сценарии погружения и фазе опыта по отдельности.Чтобы исправить множественные сравнения, мы выполнили полуавтоматическое моделирование методом Монте-Карло, реализованное в Freesurfer 5. Этот метод выполняет моделирование для получения оценки распределения максимального размера кластера при нулевой гипотезе. Следуя этому методу, мы идентифицировали кластеры с вершинным порогом p <0,01 и кластерным порогом p <0,05 (т. Е. Кластеры с вероятностью ниже порога p < .05 появиться во время моделирования нулевой гипотезы).В дополнение к кластерам на поверхности мозга мы сообщаем о подкорковых кластерах с порогом p <0,001 (нескорректированный) и размером кластера k > 20. 4. ОбсуждениеНасколько нам известно, Настоящий нейровизуализационный эксперимент является первым явным испытанием конструкционистской функциональной архитектуры разума, оценивая как сходства, так и относительные различия нейронных коррелятов телесных чувств, эмоций и мыслей. Наши результаты подтверждают конструкционистскую гипотезу о том, что психические состояния лучше всего понять, исследуя относительные различия в задействовании распределенных сетей, которые поддерживают психологические процессы, задействованные для создания различных психических состояний.Эти результаты напрямую ставят под сомнение точку зрения факультетов психологии, согласно которой разные классы психических состояний категорически различаются на уровне организации мозга. Кроме того, наш метод погружения в сценарий открывает новые возможности для использования фМРТ для понимания основных строительных блоков разума. 4.1. Психические состояния имеют общие психологические «ингредиенты»Мы обнаружили доказательства того, что телесные чувства, эмоции и мысли, хотя и различны субъективно, в каждом из них участвует одни и те же распределенные сети мозга, которые можно описать применительно к основным психологическим процессам.Там, где это возможно, мы почерпнули знания о функциях сети из литературы, но эта область находится только на самых ранних стадиях обсуждения психологических функций внутренних сетей. В результате предыдущие исследования, дающие четкое функциональное описание психологических процессов в сетях, немногочисленны (за исключением сети по умолчанию; например, Buckner & Carroll, 2007; Schacter, Addis, & Buckner, 2007; Spreng, et al. др., 2009). Однако существует множество исследований отдельных областей мозга в каждой сети.Таким образом, при обсуждении наших результатов мы обсуждаем функции отдельных регионов, составляющих сети, чтобы мотивировать понимание участия сети в целом в базовом психологическом процессе, который влияет на все психические состояния. 4.1.1 Сеть значимостиОдним из наиболее заметных открытий было общее участие сети значимости в чувствах, эмоциях и мыслях тела. Мы предположили, что сеть значимости поддерживает представления аффективных состояний, чтобы направлять внимание и поведение (Lindquist & Barrett, в обзоре; Medford & Critchley, 2010).Эта гипотеза согласуется с выводами о том, что сеть значимости демонстрирует повышенную активность в задачах, требующих распределения внимания на вызывающие воспоминания или поведенческие стимулы (Corbetta, Patel, & Shulman, 2008; Corbetta & Shulman, 2002; Nelson, et al., 2010; Сили и др., 2007). Как было определено Yeo et al., Сеть выступов состоит из передней и средней части островка, задних частей vlPFC (pars opercularis), аспектов передней средней коры головного мозга (aMCC) и передней части TPJ (супрамаргинальной извилины). ).В нашем исследовании мы обнаружили повышенную активность, в частности, в aMCC, дополнительной моторной области (SMA), дорсальной части переднего островка (AI), pars opercularis (BA 44) и левом височно-теменном соединении (TPJ) во всех областях. психические состояния. Мы обсудим каждый из этих результатов по очереди. Результат, в котором телесные чувства, эмоции и мысли коллективно задействовали SMA, согласуется с ролью SMA во внутренне обусловленных действиях (Начев, Кеннард, Хусейн, 2008). Этот паттерн активности может отражать моторные процессы, связанные с нажатием кнопки, посредством которых участники указали, что они сконструировали психическое состояние во время фазы опыта. Наш вывод о том, что TPJ задействован во всех психических состояниях, согласуется с исследованиями, которые показывают повышенную активность во время чувства свободы воли над собственным телом (обзоры см. В Decety & Grezes, 2006; Tsakiris, Constantini & Haggard, 2008) и при отображении состояния тела другого человека (как при эмпатии Lamm, et al., 2010). Действительно, повышенная активность TPJ часто наблюдается, когда один человек занимается теорией разума – или пытается понять содержание разума другого – в целом (Saxe & Kanwisher, 2003). Результат, что AI и aMCC задействованы в разных психических состояниях, согласуется с исследованиями, которые показывают активацию этих областей во время интероцепции (Critchley et al., 2003; Critchley, et al., 2004) и субъективными переживаниями в целом (Craig, 2002, 2009). Действительно, недавний метаанализ обнаружил повышенную активность в регионах, входящих в сеть значимости Йео и др. (Включая AI, aMCC, vlPFC, таламус и миндалевидное тело) во время переживания неприятного основного аффекта (Hayes & Northoff, 2011).В другой недавней статье многие из этих регионов (вентральный ИИ, таламус и миндалевидное тело) были связаны с переживанием аффективного возбуждения при просмотре негативных изображений (Touroutoglou et al. В печати). Наши результаты согласуются с обоими этими исследованиями в том смысле, что мы обнаружили активность в аналогичных регионах в пределах сети значимости для сценариев, которые были неприятными и очень возбуждающими. Таким образом, наши результаты не могут говорить о том, задействована ли сеть значимости во всех основных аффективных состояниях, поскольку мы не включали позитивные сценарии в наше исследование.Однако в предыдущем метаанализе (Вейджер, Барретт, Блисс-Моро, Линдквист, Дункан, Кобер и др., 2008 г.) мы обнаружили повышенную активность в аспектах ИИ и МКК в сторону приятных основных аффективных состояний, что согласуется с нашей гипотезой. что регионы, составляющие сеть значимости, связаны с опытом всех основных аффективных состояний. Относительная роль сети значимости в отношении неприятного и приятного основного аффекта должна стать темой будущих исследований. Что касается наших открытий, касающихся ИИ, важно отметить, что сеть значимости (названная «вентральным вниманием» Yeo et al., 2011), использованные в настоящем исследовании, охватывали как дорсальный, так и вентральный аспекты. Однако в литературе есть свидетельства того, что ИИ может быть функционально разделен на дорсальный и вентральный аспекты, при этом дорсальные аспекты функционально связаны с аМСС и относительно более вовлечены в задачи, в которых информация о теле используется для направления внимания (Kurth, Zilles, Fox, Laird, & Eickhoff, 2010; Touroutoglou, et al., В печати; Wager & Barrett, 2004). Вентральные аспекты ИИ, напротив, функционально связаны с вентромедиальной префронтальной корой и относительно больше задействованы в задачах, связанных с переживанием основных аффективных чувств (Hayes & Northoff, 2011; Touroutoglou, et al., под давлением; Wager & Barrett, 2004). Активации, которые мы наблюдали в ИИ, были в основном в спинной части, что может отражать роль этой области в устойчивом внимании к важной информации (см. Lindquist & Barrett, в обзоре; Medford & Critchley, 2010; Seeley et al., 2007). . Такая заметность могла быть особенно важной в нашей задаче по погружению, потому что участники переключали внимание между различными источниками информации (например, внутренние ощущения против внешних ощущений), когда они активно создавали психические состояния. Наши результаты показывают, что репрезентации телесных ощущений играют роль, выходящую за рамки физических ощущений в теле или аффективных состояний, таких как эмоции. В любой ситуации, когда людям представлена вызывающая воспоминания или поведенческая информация, сеть значимости будет определять направление внимания на основе телесных ощущений, независимо от того, направлены ли люди на переживание эмоции или объективное мышление о ситуации. Этот вывод согласуется с несколькими недавними предположениями в литературе о том, что телесные сигналы являются повсеместным компонентом психической жизни, в том числе в состояниях, связанных с восприятием (Barrett & Bar, 2009; Cabanac, 2002), суждением (Clore & Huntsinger, 2007), задачами, включающими усилия (Critchley, et al., 2003), и в сознании в целом (Craig, 2009; Damasio, 2000). Этот вывод также согласуется с недавними исследованиями, демонстрирующими, что представление концепций эмоций включает моделирование аффективных состояний и состояний тела (Oosterwijk, Rotteveel, Fischer, & Hess, 2009; Oosterwijk, Topper, Rotteveel, & Fischer, 2010), а также с исследованиями, которые предполагают что представления состояний тела участвуют, когда участники понимают словесные описания «когнитивных» состояний (например, мышление, запоминание или внезапное озарение) (Oosterwijk et al., 2012). 4.1.2 Лимбическая сетьВопреки нашему прогнозу, наши результаты не выявили устойчивой активации корковых аспектов лимбической сети, как было выявлено Йео и др. (2011) во время переживания телесных чувств, эмоций и мыслей. Однако наш анализ продемонстрировал вовлечение подкорковых частей лимбической сети, которые менее восприимчивы к выпадению сигнала, особенно в паллидуме и скорлупе. Исследования, предполагающие, что базальные ганглии играют роль в мотивированном поведении (Graybiel, 2005, 2008), согласуются с гипотезой о том, что эти структуры участвуют в генерации основных аффективных состояний.Например, функция базальных ганглиев показывает взаимосвязь с тяжестью симптомов расстройств, при которых генерация аффектов нарушена. В частности, степень связи базальных ганглиев с другими областями в пределах предполагаемой сети генерации основных аффектов, таких как вентромедиальная префронтальная кора головного мозга, связана с тяжестью симптомов депрессии (Marchand et al., 2012), а поражения базальных ганглиев приводят к большей частоте возникновения депрессия после инсульта, чем поражения в других частях мозга (Morris, Robinson, Raphael, & Hopwood, 1996; Vataja et al., 2004). Миндалевидное тело, еще одна подкорковая структура в нашей предполагаемой лимбической сети, не продемонстрировало повышенной активности в нашем исследовании. Хотя предполагается, что миндалевидное тело обычно участвует в эмоциях, более пристальный взгляд на литературу показывает, что активность миндалевидного тела с большей вероятностью будет происходить во время сенсорного восприятия поведенческих стимулов, чем во время внутренне сфокусированного опыта аффективных психических состояний, как это произошло в наша задача (метаанализ см. в Costafreda et al.2008; Lindquist et al. под давлением). В соответствии с нашими выводами недавнее исследование не смогло зафиксировать повышенную активность миндалины, когда участники испытывали социальный стресс в сканере (Wager et al. 2009). 4.1.3 Сеть по умолчаниюМы обнаружили общее участие сети по умолчанию в ментальных состояниях, что согласуется с гипотезой о том, что эта сеть необходима для построения ситуативной концептуализации, которая создает состояние тела, эмоции или мысли. Гипотеза о том, что сеть по умолчанию поддерживает концептуализацию, согласуется с ролью сети по умолчанию в атрибуции психического состояния (Mitchell, et al., 2005), эмоциональный опыт (Kober, et al., 2008; Lindquist, Wager, Kober, et al., В печати), воспоминания об автобиографическом прошлом (Addis, et al., 2007; Spreng & Grady, 2010), спонтанное мышление (Andrews-Hanna, Reidler, Huang, & Buckner, 2010) и семантическая и концептуальная обработка (Binder, et al., 2009; Visser, Jefferies, & Lambon Ralph, 2010). Мы предполагаем, что то, что объединяет эти разрозненные области, – это процесс концептуализации, в котором репрезентации предшествующего опыта используются для построения репрезентаций прошлого, будущего или настоящего момента.Эти области необходимы для придания значения действиям других, для придания значения собственному базовому аффективному состоянию, для вспоминания предшествующего опыта во время примеров памяти и спонтанного мышления и для представления значения концепций путем моделирования экземпляров категорий (для обсуждения, см. Lindquist & Barrett, на обзоре; Lindquist, Wager, Kober, et al., в печати). Сеть по умолчанию, как определено Йео и др., Состоит из аспектов медиальной префронтальной коры (mPFC), аспектов треугольной части и орбитальной части, верхней височной доли, угловой извилины и аспектов предклинья. .В нашем исследовании мы обнаружили общее вовлечение боковой орбитофронтальной коры и треугольной части (BA 45) через телесные чувства, эмоции и мысли. Эти области были особенно связаны с семантической обработкой (Binder et al. 2009; Visser et al. 2010) и могут отражать подсеть (см. Yeo et al. 2011, 17-network parcellation; Smith et al. В печати) внутри сеть по умолчанию, которая специально поддерживает язык (Lindquist & Barrett, на рассмотрении; Smith et al. в печати).Примечательно, что наш анализ конъюнкций не привел к увеличению активности во всей сети по умолчанию. Однако наш анализ конъюнкции не привел к какому-либо значительному снижению в основных регионах сети по умолчанию, предполагая, что телесные чувства, эмоции и мысли задействовали сеть по умолчанию в той же степени, что и спонтанные мысли, наблюдаемые во время фиксации (Эндрюс -Hanna et al.2010). Поскольку многие основные области в сети по умолчанию (например, mPFC, PCC) демонстрируют повышенную активацию во время фиксации и, таким образом, проявляются как пониженная активность во время других задач (отсюда и название сети «по умолчанию»; Raichle et al., 2001), мы, вероятно, поставили себя в невыгодное положение, обнаружив устойчивое увеличение сети по умолчанию во время построения психического состояния, сравнивая активность задачи с фиксацией. В соответствии с этой интерпретацией, мы обнаружили прогнозируемую активацию в классических областях по умолчанию (например, mPFC, PCCC) в нашем контрастном анализе (описанном ниже), когда мы сравнивали один тип психического состояния с другим. 4.1.4 Лобно-теменная сетьМы обнаружили общее участие аспекта лобно-теменной сети в состояниях тела, эмоциях и мыслях, предполагая, что все эти психические состояния связаны с исполнительным вниманием.Области мозга в этой сети, как правило, имеют повышенную активность во время «контроля внимания сверху вниз» (Corbetta & Shulman, 2002), например, при «поддержании памяти» (Cole & Schneider, 2007), «представлении правил» (Crone, Wendelken, Donohue, & Bunge, 2006) и «планирование» (Fincham, Carter, van Veen, Stenger, & Anderson, 2002). Мы предполагаем, что эта сеть модулирует активность в других функциональных сетях, чтобы помочь построить экземпляр психического состояния. Активность в этой сети может отражать когнитивный контроль, необходимый для реализации процессов концептуализации основного аффекта.Лобно-теменная сеть, как было определено Yeo et al., Состоит из аспектов дорсолатеральной префронтальной коры, аспектов поясной коры, лобного острова и аспектов нижней теменной доли и предклиния. В нашем исследовании в дорсолатеральной префронтальной коре наблюдалась повышенная активность телесных чувств, эмоций и мыслей. 4.2 Различные профили или «рецепты ингредиентов» по-разному влияют на разные психические состоянияХотя одни и те же распределенные сети мозга участвовали в телесных чувствах, эмоциях и мыслях, мы также обнаружили доказательства того, что разные сетевые профили были связаны с каждой категорией психического состояния .По сравнению с мыслями, телесные ощущения включали относительно большее увеличение активности в сети значимости (то есть, дорсальный передний островок с двух сторон, левый vlPFC, включая pars opercularis, и правый aMCC). Эмоции также продемонстрировали повышенную активацию в сети значимости по сравнению с мыслями (то есть в левом vlPFC, правом aMCC и правом TPJ), хотя и в немного разных областях. Кроме того, телесные ощущения вовлекают подкорковые области, которые, как предполагается, участвуют в основной аффективной генерации, такие как таламус, паллидум и хвостатое, в большей степени, чем мысли. По сравнению с телесными ощущениями, эмоции вовлекали относительно большее участие сети по умолчанию, особенно в левой передней височной доле (ATL). Аспекты ATL являются частью сети по умолчанию и связаны с представлением концептуальных знаний (Binder, et al., 2009; Visser, et al., 2010). Эти данные предполагают, что переживание эмоции по сравнению с состоянием тела характеризуется более сильным вовлечением процесса концептуализации для придания значения телесным ощущениям в данный момент.Эти результаты согласуются с поведенческими данными, демонстрирующими, что такие понятия, как «гнев» или «страх», используются для преобразования телесных ощущений в переживание эмоции (например, Lindquist & Barrett, 2008a). Точно так же истончение кортикального слоя при ATL из-за нейродегенеративного заболевания ухудшает способность пациентов воспринимать эмоциональные выражения лица как примеры дискретных эмоциональных чувств (например, гнев против страха) (Lindquist, Gendron, Barrett, & Dickerson, рассматривается). Мысли также продемонстрировали большее вовлечение сети по умолчанию (двусторонняя верхняя височная извилина, левая предклинье и левая мПФК) по сравнению с телесными ощущениями.Кроме того, несколько регионов в сети по умолчанию, включая левую и правую верхнюю височную извилину и левую mPFC, продемонстрировали относительно большее увеличение активности, когда участники генерировали мысли, чем когда они генерировали эмоции. Этот вывод согласуется с нашим предположением о том, что концептуализация играет большую роль в ментальных состояниях, когда людям необходимо планировать, ассоциировать и размышлять над ситуацией (мыслью), чем в ментальном состоянии, в котором основное внимание уделяется телесным ощущениям. Более того, это открытие также предполагает, что мысль может включать процесс концептуализации в большей степени, чем эмоции.Эта интерпретация согласуется с устойчивым участием сети по умолчанию в памяти и предсказании будущего (Addis, et al., 2007), теории разума (Spreng & Grady, 2010), семантической памяти (Binder, et al., 2009; Visser, et al., 2010) и другие так называемые «когнитивные» психические состояния, связанные с репрезентацией предшествующего опыта. Конкретная комбинация активации в регионах в сети по умолчанию и регионах в сети значимости (как видно из анализа конъюнкции) во время мысли и эмоций согласуется с недавним исследованием, которое продемонстрировало связь между регионами сети по умолчанию (rTPJ, vmPFC и PCC / preduneus), аспекты сети значимости (передний островок, SMA, aMCC) и первичной сенсомоторной коры во время задач ментализации (Lombardo et al., 2009). Интересно, что эта модель взаимосвязи возникла как тогда, когда участники мысленно относились к себе, так и когда они мысленно относились к другим, предполагая перекрытие между двумя процессами, которые были разделены в литературе. По мнению авторов, этот паттерн связности предполагает взаимодействие между «воплощенными / основанными на симуляции представлениями» и областями, которые поддерживают «высокоуровневую ментализацию, основанную на умозаключениях». Эта точка зрения согласуется с конструкционистской точкой зрения, согласно которой представления сенсомоторных и соматовисцеральных ощущений сочетаются с концептуализацией для создания определенного ментального содержания.В будущих исследованиях следует оценить степень, в которой сеть значимости и сеть по умолчанию различаются с точки зрения функциональной связи между эмоциями, состояниями тела и мыслями. Например, одна из гипотез состоит в том, что исходные области в сети значимости и сети по умолчанию демонстрируют большую взаимосвязь во время эмоций и мыслей (которые требуют большей концептуализации) по сравнению с телесными чувствами. Хотя у нас не было априорных гипотез об относительной вовлеченности лобно-теменной сети в психические состояния, мы наблюдали, что эта сеть была относительно более активной, когда люди сосредоточивались на чувствах своего тела, чем когда люди генерировали эмоции.Это открытие дает некоторое представление о расстройствах, которые связаны с чрезмерной бдительностью (например, паническое расстройство и симптоматика тревожности; Schmidt, Lerew, & Trakowski, 1997). Повышенная активность лобно-теменной сети также может быть истолкована как свидетельство того, что участникам было труднее создавать ощущения тела, чем создавать эмоции или мысли в нашей задаче, хотя их субъективные оценки не поддерживали эту интерпретацию. 4.3 ПоследствияНастоящие результаты представляют несколько иную функциональную архитектуру основных строительных блоков человеческого разума, чем то, что предполагается с точки зрения факультета психологии.В отличие от традиционных подходов факультетской психологии, которые были популярны в когнитивной нейробиологии в течение последних трех десятилетий, наши результаты показывают, что полезно использовать конструкционистские рамки при попытке понять, как мозг создает ментальные состояния, в которых мысли и чувства возникают из сочетание внутренних сетей. Эти сети можно описать в терминах основных, общих психологических компонентов предметной области (таких как основной аффект, концептуализация, исполнительное внимание и экстероцептивное ощущение), которые можно наблюдать как элементы множества психических состояний, составляющих человеческий разум.Наши результаты показывают, что в рамках этой функциональной архитектуры продуктивно сосредоточиться на широкомасштабных внутренних нейронных сетях при интерпретации данных функциональной нейровизуализации. Эта идея согласуется с растущим призывом к сетевому пониманию функций мозга (Bullmore & Sporns, 2009; Fuster, 2006; Goldman-Rakic, 1988; McIntosh, 2000; Mesulam, 1998; Pessoa, 2008). Наши результаты также подчеркивают важность изучения распределенных паттернов активации мозга для понимания психических состояний с разным содержанием, а не сосредоточения внимания на отдельных областях.В наиболее экстремальной версии этого подхода психологическая функция существует как функциональное взаимодействие этих областей, так что изолированные области выполняют различные психологические функции в зависимости от того, с чем они связаны в конкретном случае. В будущих направлениях будет важно использовать многомерные методы (например, McIntosh, Bookstein, Haxby, & Grady, 1996), функциональный анализ связности и другие нейроинформатические подходы, которые позволят исследователям понять, как отдельные области мозга работают вместе, для создания экземпляра основных психологические процессы при психических состояниях. Во-вторых, настоящие открытия проливают новый свет на функцию определенных областей мозга, поскольку они вносят вклад в широкомасштабные функциональные сети. Например, мы обнаружили повышенную активацию в области, известной как TPJ, во всех психических состояниях в нашем исследовании. Тем не менее, TPJ иногда называют «нейронным коррелятом» восприятия мыслей других людей, исходя из предположения, что он последовательно и специально выполняет нейронные вычисления для теории разума (Saxe & Kanwisher, 2003; Young, Cushman, Hauser, & Saxe, 2007 ).Наши результаты показывают, что TPJ вряд ли будет играть такую специфическую роль (аналогичную точку зрения см. Mitchell, 2008), поскольку его аспекты (например, надмаргинальная извилина) являются частью более крупной сети значимости, в которой мы наблюдали повышенную активность во время генерации. собственных психических состояний (а не просто размышления о чужом опыте) (другое обсуждение см. Decety & Grezes, 2006). Если сеть значимости (включая TPJ) участвует (по крайней мере частично) в представлении основного аффекта, это объясняет ее участие в задачах, связанных как с генерацией, так и с восприятием ментальных состояний.В исследованиях восприятия психического состояния другими людьми (т.е.теории разума) TPJ мог иметь повышенную активность, потому что другие люди являются значимыми стимулами (и, следовательно, представлены как имеющие основную аффективную ценность) (Mitchell, 2008). В качестве альтернативы может потребоваться моделирование основного аффективного состояния другого человека, когда он пытается понять, что он думает и чувствует (т. Е. Теория разума опирается на воплощение аффективных состояний; Ломбардо и др., 2009). Еще другие аспекты TPJ (угловая извилина) являются частью сети по умолчанию, участвующей в концептуализации.Одна из возможностей состоит в том, что роль TPJ зависит от его «нейронного контекста», в котором функция области мозга зависит от других областей, с которыми она функционально связана в данный момент времени (например, McIntosh, 2000). Другая возможность состоит в том, что существуют разные функциональные подобласти TPJ, которые по отдельности вносят вклад в основной аффект и концептуализацию. Эта основа для понимания функции области мозга также применима к другим областям, которые показали повышенную активность в нашем исследовании (например, MPFC, aMCC, островок), которые в литературе приписываются определенным психическим состояниям (например,g., самореференционная обработка; Келли и др., 2002; например, конфликт; Кернс, 2004; например, отвращение; Wicker, et al., 2003). 4.4. ОграниченияРезультаты настоящего исследования породили вопросы, которые могут стать темой будущих исследований. Например, неожиданным открытием было то, что различия между психическими состояниями возникали на разных этапах экспериментальной задачи. Например, различия в активности в сети значимости были наиболее выражены между психическими состояниями, когда участники погружались в сценарий.Напротив, фаза переживания не показала различий в пределах сети значимости между телесными чувствами, эмоциями и мыслями, предполагая, что все психические состояния в одинаковой степени включали аффективные процессы во время фазы переживания. Эти результаты можно понять, если учесть, что телесные чувства формировались быстрее, чем эмоции и мысли. Участники могли начать формирование телесных ощущений на ранней стадии фазы погружения, что соответствовало более сильному вовлечению сети значимости во время погружения и более короткому времени реакции во время фазы опыта.Тем не менее, эта интерпретация не объясняет более сильного вовлечения сети значимости во время фазы погружения в эмоции. Другая возможная интерпретация этого открытия заключается в том, что подсказка участникам испытать состояние тела или эмоции заставила их обратить внимание на тело и аффективные детали сценария за исключением других деталей, тогда как в мыслях они обращали внимание на не связанные с телом / аффективные детали. Мы не можем исключить такую интерпретацию, учитывая параметры дизайна исследования.Одним из способов прояснения этого вопроса в будущих исследованиях было бы запретить участникам видеть сигнал до тех пор, пока они не услышат сценарий. И все же третья интерпретация этого открытия заключается в том, что телесные чувства и эмоции задействуют сеть значимости больше, чем мысли, но только в начальные моменты опыта (например, когда он изначально создается), а не по мере того, как опыт сохраняется с течением времени. То есть относительное участие различных распределенных сетей во время создания психических состояний может зависеть не только от типа психического состояния – оно также может зависеть от временного хода состояния.Этот вопрос – тема будущих исследований. Еще одно открытие настоящего исследования, которое заслуживает большего экспериментального внимания, – это открытие, что состояния тела и мысли более отличаются друг от друга, чем от эмоций. Хотя возможно, что наши манипуляции эмоциями были менее устойчивыми, чем другие экспериментальные условия, или что участники не адекватно создавали эмоции в ответ на сценарии, эта интерпретация не подтверждается результатами нашего самоотчета.Участники сообщили, что они добились такого же успеха в создании эмоций, как состояния тела и мысли, и что они испытывали эмоции так же ярко, как и другие состояния. В будущих исследованиях следует попросить участников оценить специфичность, с которой они испытали каждое состояние на основе испытания за испытанием. Окончательная интерпретация этого открытия, которая была априори предсказана нашей конструкционистской точкой зрения, состоит в том, что по замыслу (а не в результате экспериментальной неудачи) эмоции разделяют многие «ингредиенты», которые также включают как состояния тела, так и мысли.Мы предположили (Barrett, 2006) и продемонстрировали (Lindquist & Barrett, 2008a) в другом месте, что эмоции – это состояния, которые одновременно являются аффективными и концептуальными. В будущих исследованиях следует продолжить изучение того, в какой степени эмоции представляют собой психические состояния, состоящие как из аффективных, так и из концептуальных компонентов. Наконец, ограничение настоящего исследования состоит в том, что в нем не измерялись индивидуальные различия в когнитивных способностях, которые могли ограничивать способность участников конструировать психические состояния.Одна из гипотез конструкционистской структуры состоит в том, что люди с большей емкостью рабочей памяти смогут лучше получить доступ к релевантной концептуальной информации и запретить нерелевантную концептуальную информацию во время построения ментальных состояний (Barrett, Tugade & Engel, 2004; Lindquist & Barrett, 2008b). Таким образом, в будущем было бы интересно смоделировать степень, в которой люди с большей емкостью рабочей памяти не только более успешны в такой задаче, но и с большей вероятностью будут иметь дискретные и устойчивые психические состояния в целом. 4.5 Будущие направленияВ данной статье мы попытались описать каждую внутреннюю сеть в психологических терминах. Важно признать, что эти психологические описания являются первой гипотезой и только отправной точкой. В будущих исследованиях будет важно продолжить определение и уточнение конструкционистской структуры, которую мы изложили в этой и других статьях (Barrett, 2006, 2009; Barrett et al., 2007; Barrett, Lindquist, & Gendron, 2007; Барретт, Мескита и др., 2007; Линдквист и Барретт, 2008a, 2008b; Линдквист, Вейджер, Блисс-Моро и др. В печати; Линдквист, Вейджер, Кобер и др. В печати; Wilson-Mendenhall и др., 2011). Лучшие психологические описания существующих сетей, несомненно, будут меняться по мере накопления данных с течением времени. Возможно, дальнейшая цель состоит в том, чтобы описать психологически самые основные «общие знаменатели», которые свяжут ряд находок в разных областях задач, которые на первый взгляд кажутся очень разными (см. Lindquist & Barrett, в обзоре).Например, определение сети значимости как функциональной группы, которая представляет основную аффективную информацию от тела для направления внимания и поведения, объясняет не только роль этой сети в эмоциях, но и в фокусе внимания, мониторинге производительности, языке, обработке сенсорных данных и т. Д. сознание в более общем плане (обзоры см. Craig, 2009; Nelson, et al., 2010). Важно отметить, что конструкционистская концепция не просто переименовывает сети, чтобы соответствовать одной конкретной теоретической точке зрения.Вместо этого он пытается предоставить функциональные описания распределенных сетей, которые содержат многолетние нейровизуализационные исследования эмоций, памяти, мышления, семантики и так далее, чтобы пролить новый свет на то, как сознание отображается на мозг. Анализ внутренних сетей остается важным направлением будущих исследований, поскольку наука пытается определить самые основные общие знаменатели психологического опыта. Например, явное моделирование времени позволило исследователям идентифицировать пространственно различные внутренние сети, которые также максимально различны с точки зрения их временной динамики (Smith et al., под давлением). Smith et al. продемонстрировать, например, что сеть по умолчанию может быть далее проанализирована на определенные во времени подсети, каждая из которых может иметь различные психологические функции (например, язык v., представляющий предыдущий опыт). Благодаря таким уточнениям исследователи будут лучше оснащены для обнаружения строительных блоков психических состояний. Наконец, хотя конструктивный подход все еще недостаточно определен, а дальнейшая проработка и уточнение имеют решающее значение, наши результаты дополняют растущий объем литературы, в которой предполагается, что факультетская психология должна быть отброшена как научная основа для понимания того, как мозг создает разум.Результаты нашего исследования показывают, что мозг не может быть разделен на разные области для разных умственных способностей, которые соответствуют уникальным умственным переживаниям индивидуально. Как возникающее явление, психические состояния с особой феноменологией, такие как телесные чувства, эмоции и мысли, не обязательно локализуются в отдельных регионах (или даже сетях) в человеческом мозгу. Люди обладают способностью испытывать самые разные психические состояния, и только изучая это разнообразие, мы сможем, наконец, понять основные строительные блоки, из которых они состоят. Уникальная структура и функция вирусных родопсиновОбщая структура OLPVRIIOLPVRII была кристаллизована с использованием подхода in meso , аналогичного тому, который использовался в наших предыдущих работах 10,14,21 . Обычно для в кристаллизации мезо пирамидальные кристаллы типа I размером до 50 мкм (дополнительный рис. 4а) появлялись в течение 3 месяцев. Кристаллы дифрагировали до 1,9 Å, а структура OLPVRII была решена при 1,9 Å. Пространственная группа кристаллов была определена как P2 1 с параметрами решетки ячеек a = 79.58 Å, b = 99,70 Å, c = 82,96 Å, α = γ = 90 ° и β = 116,95 °. Кристаллическая упаковка и примеры карт электронной плотности представлены на дополнительных рисунках 4b-e. Асимметричная элементарная ячейка OLPVRII содержит пять молекул белка, организованных в виде пентамера с бутылкообразной порой в центре, имеющей узкую шейку в цитоплазматической части (рис. 1a, d). Вокруг пентамера наблюдаются несколько липидных фрагментов с неупорядоченными полярными головками.Кроме того, хорошо упорядоченная молекула моноолеина (МО), молекула-хозяина кристаллизационной матрицы, наблюдается между спиралью D одного протомера и спиралью G ‘соседнего пентамера. МО находится глубоко внутри гидрофобной бороздки, образованной Phe197, Ile200, Tyr201, Phe205, Phe210 одного мономера и Phe103 ‘другого мономера (дополнительный рис. 5). Расположение полярной головки МО стабилизируется за счет водородной связи с основной цепью Phe210 и через Wat20 с Ala204 и Ile 209 на С-конце. Рис. 1Архитектура контактов пентамера и межпротомера OLPVRII. а Вид со стороны цитоплазмы. Поверхностное представление пентамера. Центральная пора обведена красным кружком. б Вид со стороны цитоплазмы. Мультяшное изображение пентамера. Кофактор сетчатки окрашен в голубой цвет. c Детальный вид основной области межпротомерных контактов. Протомеры C, D и E окрашены в серый цвет. d Пентамер, вид сбоку.Один протомер скрыт для ясности. Полость внутри пентамера была рассчитана с использованием HOLLOW 69 , она окрашена в голубой цвет, границы ядра гидрофобной мембраны показаны сплошными горизонтальными линиями. e , f Детальный вид межпротомерных контактов Организация пентамера OLPVRIIПентамер OLPVRII стабилизирован плотной сеткой водородных связей. Наиболее интенсивные контакты между молекулами белка в пентамере расположены на цитоплазматической стороне белка, где спираль A одного протомера вставлена в щель между спиралями A ‘и B’ другого протомера (рис.1б). Существование всей сети обусловлено наличием двух высокоупорядоченных молекул воды. Первый (Wat12) координирует водородные связи боковых цепей His37 и Asn40 одного протомера и Trp203 ‘и Glu26’ другого (рис. 1c, f). Asn40 стабилизирован также водородной связью с кислородом основной цепи Ile22 ‘. Вторая молекула воды координирует взаимодействия между Arg36 и атомами кислорода основной цепи Ala27 и Thr30 для дальнейшей стабилизации положения боковой цепи Arg29 ‘соседней молекулы белка.В центре пентамера атомы кислорода боковых цепей Leu28 и Arg29 всех пяти протомеров также связаны сильными водородными связями, опосредованными пятью молекулами воды (Wat13). Более того, Glu26 ‘второго протомера сильно взаимодействует с Arg36 из того же протомера, обеспечивая связь между кластерами опосредованных водой водородных связей. Таким образом, следующие аминокислоты играют ключевую роль в образовании пентамера: Glu26, Arg36, His37, Asn40, Trp203, и эти аминокислоты являются высококонсервативными в группе II вирусных родопсинов (дополнительный рис.1), но не в других родопсинах. Кроме того, искаженный C-конец одного протомера взаимодействует с плохо упорядоченной C-D петлей соседнего протомера, образуя несколько водородных связей между их атомами основной цепи и, таким образом, стабилизируя пентамер (Fig. 1b, e). Чтобы доказать, что наблюдаемая пентамерная сборка не является артефактом кристаллизации, мы провели эксперименты по сшиванию с белком дикого типа. С помощью эксклюзионной хроматографии белок был разделен на две фракции, соответствующие мономерам и пентамерам (дополнительный рис.6а). Эксперименты по сшиванию четко подтверждают, что мультимерные фракции, появляющиеся на эксклюзионной хроматографии, содержат пентамеры (дополнительный рис. 6b, дорожка 4). Более того, мы воссоздали мономеры солюбилизированного белка дикого типа в липосомы, и эксперименты по сшиванию показали, что в этих липосомах белок OLPVRII находится в пентамерной форме (дополнительный рис. 6b, дорожка 5). Таким образом, пентамеры сохраняют свою олигомерную форму во время очистки и очищенные в детергенте мономеры, помещенные в липидную среду, снова собираются в пентамеры, особенно во время кристаллизации.Кроме того, мы экспрессировали и очистили тройной мутант E26A / R36A / W203A с тремя мутациями высококонсервативных аминокислот, которые расположены на границе раздела между протомерами и образуют водородные связи между ними. В то время как белок дикого типа имел соотношение пентамер: мономер, равное 4: 1 в профиле элюирования, тройной мутант показал соотношение приблизительно 1: 1 (дополнительный рис. 6а). Тот факт, что тройной мутант частично сохранил пентамерную сборку, предполагает, что существуют дополнительные взаимодействия между протомерами OLPVRII, такие как образование водородных связей между петлей C-D и C-концом и гидрофобная кооперация.Принимая во внимание, что ключевые аминокислоты, образующие пентамер, являются высококонсервативными в вирусных родопсинах второй группы (дополнительный рис. 1), мы полагаем, что пентамерная сборка имеет решающее значение для функции OLPVRII. Пентамерная сборка наблюдалась также для других микробных родопсинов, таких как PR и NaR 22 (дополнительный рис. 7). Хотя относительная ориентация протомеров аналогична для OLPVRII, KR2 и BPR HOT75 (дополнительный рис.7), размер и профиль центральной поры вместе с контактами пентамеризации полностью различны в этих белках (дополнительный рис.8). Олигомеризация PR и NaR влияет на функциональность этих белков. Например, в KR2, NaR из Krokinobacter eikastus , для перекачки натрия требуется пентамеризация 23 . Таким образом, пентамеризация также может быть ключевым фактором, определяющим активность вирусных родопсинов группы II, и для выяснения его функции необходим тщательный анализ пентамера OLPVRII. Центральная пора (канал) пентамерной структурыОднако описанные выше контакты между молекулами OLPVRII необычны для микробных родопсинов.Они приводят к образованию поры в форме бутылки внутри пентамера, образованной спиралями A и B протомеров (рис. 1d, 2a). Самый узкий участок (преддверие) поры образован Phe24, Leu28 и Arg29 спирали A и имеет длину 11 Å и средний диаметр 6 Å. Примечательно, что Phe24 и Arg29 являются высококонсервативными в вирусных родопсинах группы II, тогда как Leu28 заменен другими гидрофобными остатками, Ile и Met (дополнительный рис. 1). У входа в пору, на цитоплазматической стороне пентамера, уникальное расположение боковых цепей Arg29 позволяет расположить пять соседних положительно заряженных аминокислот.В дальнейшем мы будем обозначать эту конфигурацию кольцом Arg29. Рис. 2Центральная пора внутри пентамера OLPVRII. a Общая структура центральной поры. Один протомер скрыт для ясности. Липидный фрагмент окрашен в фиолетовый цвет. b Вид с цитоплазматической стороны на пятиугольник воды и сеть взаимодействия между молекулами воды и остатками OLPVRII, выстилающими поры. c Вид на преддверие поры сбоку. Пример карты электронной плотности 2Fo-Fc показан вокруг пятиугольника воды и углеводородной цепи.Карта оконтурена на уровне 1,2 σ . Углеводородная цепь окрашена в фиолетовый цвет Глубже в пору атомы кислорода основной цепи Leu28 координируют пять молекул воды, образующих пятиугольник, стабилизированный водородными связями, со средним расстоянием между двумя соседними молекулами 2,6 ± 0,2 Å (среднее ± стандартное для разных молекулы пентамера, рис. 2б). Этот пятиугольник слабо взаимодействует с боковыми цепями Arg29, поскольку молекулы воды расположены в пределах 3,8 ± 0,3 Å (среднее ± стандартное отклонение для разных молекул пентамера) от ближайших атомов азота аргинина.Боковые цепи Leu28 и Phe24 образуют чрезвычайно гидрофобную часть преддверия (дополнительный рис. 9). Удивительно, но мы наблюдали сильную концентрацию электронов вдоль оси поры в этой области (рис. 2в). Мы отнесли его к короткой углеводородной цепи, закупоривающей поры. Боковая цепь Phe24 ориентирована перпендикулярно длинной оси поры, образующей самый узкий участок преддверия с диаметром всего 5,5 Å. Ближе к внеклеточной стороне боковая цепь Phe24 образует плечо поры, где ее диаметр резко увеличивается до 26 Å.Широкая внеклеточная часть поры выстлана остатками Thr21, Val47, Met17, Ser54, Asn55, Tyr10 и Tyr13. Плечо и следующая за ним часть поры вымощены фрагментами липидных молекул (дополнительный рис. 9). Сравнение пор внутри олигомеров, образованных микробными родопсинами, с известной структурой показывает уникальность канала в пентамере OLPVRII (дополнительный рис. 8). Примечательно, однако, что аналогичная организация пор наблюдается в трансмембранных доменах пентамерных лиганд-управляемых ионных каналов (pLGIC) (дополнительный рис.10) 24 . Действительно, кольцо из пяти отрицательно заряженных аминокислот (Glu222 в каждой субъединице) играет роль в ионной селективности структурно охарактеризованной pLGIC из Gloeobacter violaceus (GLIC). Более того, кристаллическая структура GLIC (PDB ID: 4HFI) содержит аналогичные пятиугольники молекул воды внутри трансмембранного канала, предположительно участвующие в гидратации иона проницаемости, за которыми следует чрезвычайно гидрофобная область, закрытая углеродными хвостами детергента. молекулы 24 .Преддверие поры OLPVRII имитирует трансмембранный канал GLIC (дополнительный рис. 10). Такая необычная узкая цитоплазматическая часть поры, ее сходство с трансмембранными каналами pLGIC и эволюционная консервация большинства остатков, участвующих в образовании пентамера и поры, убедительно свидетельствуют о том, что центральная пора OLPVRII функционирует как канал. В pLGIC открытие и закрытие трансмембранного канала запускается конформационными изменениями, начиная со связывания лиганда в активном центре внеклеточного домена.Напротив, структурные перестройки в микробных родопсинах инициируются поглощением света кофактором сетчатки, который находится внутри белка. Следовательно, OLPVRII, вероятно, будет функционировать как пентамерный светозависимый ионный канал. Чтобы идентифицировать возможные механизмы регуляции центрального канала в OLPVRII, мы проанализировали организацию протомеров. Карман сетчатки и внеклеточная часть OLPVRIIВ пентамере OLPVRII есть лишь небольшие различия между протомерами.Среднеквадратичное отклонение (RMSD) позиций атомов основной цепи между различными протомерами варьируется от 0,17 до 0,20 Å. Таким образом, ниже мы описываем только протомер А из модели OLPVRII. Подобно другим микробным родопсинам (дополнительный рис. 11), каждый протомер OLPVRII организован как пучок из семи трансмембранных спиралей (A-G), соединенных относительно короткими петлями. Кофактор сетчатки ковалентно связан с Lys195 через основание Шиффа (RSB), и плотности электронов показывают, что хромофор находится в конформации, полностью состоящей из – транс- (дополнительный рис.4в). OLPVRII состоит всего из 211 аминокислот и является самым маленьким среди микробных родопсинов с известной кристаллической структурой. По сравнению с другими родопсинами длина белка уменьшена из-за чрезвычайно коротких N- и C-концов, а также коротких петель, не имеющих определенной вторичной структуры. Поскольку различия в структурах OLPVRII и Halobacterium salinarum бактериородопсина (bR), вероятно, отражают функциональные различия, мы подробно сравниваем структуры bR и OLPVRII.Сборка спиралей OLPVRII в пучок отличается от таковой в bR (PDB ID 1C3W), при этом RMSD позиций атомов основной цепи составляет около 1,8 Å. Важно, что в OLPVRII спираль C значительно длиннее, тогда как спираль F, наоборот, короче, чем соответствующие спирали bR (Supplementary Fig. 12). Внеклеточная часть спирали G смещена внутрь примерно на 4 Å, а наклон и положения других спиралей также немного изменяются в OLPVRII по сравнению с bR (дополнительный рис. 12). Стенки кармана, связывающего сетчатку, гидрофобны и подобны стенкам других родопсинов, за исключением областей β-иононового кольца и RSB.Однако по сравнению с bR существуют значительные различия в соседних аминокислотах: высококонсервативный Pro186 заменен на Gly173, Ser141 на Gly137 (как в PR), а Thr142 на Phe138 (рис. 3c и дополнительный рис. 13). Есть различия также на цитоплазматической стороне RSB, где Leu93 из bR заменен на Met83 и Ala215 на Ser194. Мутация A215T является основным изменением, превращающим bR в сенсорный родопсин, тогда как Leu93 и гомологичные остатки являются функционально важными и ответственными, например, за настройку цвета в PR 25,26 .Эти мутации приводят к смещению RSB к внеклеточной части белка по сравнению с bR из-за стерического конфликта с Met83 и Ser194. Рис. 3Структура протомера OLPVRII, кармана для связывания сетчатки и внеклеточной части. a Общий вид протомера A сбоку. Спирали F и G для наглядности скрыты. Гидрофобно-гидрофильные границы мембраны показаны серыми линиями. b Детальный вид внеклеточной части. c Детальный вид кармана для связывания сетчатки.Остатки на стенках кармана окрашены в бирюзовый цвет. d Детальный вид региона RSB. Спирали A и B для наглядности скрыты. Полости внутри протеинового протомера окрашены в розовый цвет. Lys195 и ковалентно связанный сетчатка окрашены в голубой цвет Как и в bR, в OLPVRII азот RSB связан водородной связью с ключевой молекулой воды Wat1 (Wat402 в bR), которая отдает водородные связи двум анионным остаткам, Asp75 и Asp191 (Asp85 и Asp212). в bR соответственно). Такое расположение стабилизирует положительный заряд RSB и сохраняется в протонных насосах, выходящих наружу, и в большинстве других родопсинов 27 .В OLPVRII структурная организация RSB, которая включает Asp75, Asp191 и Arg72, сходна с таковой для bR, но наблюдаются некоторые различия (Fig. 3a and Supplementary Fig. 13). Структурный и мутационный анализ, описанные ниже, позволяют предположить, что Asp75 является первичным акцептором протонов от RSB, а депротонирование RSB в OLPVRII происходит аналогично тому, как это происходит для bR 28 . Пространство между Arg72 и внеклеточной поверхностью белка заметно отличается в OLPVRII по сравнению с bR (рис.3 и дополнительный рис.14). В то время как в bR боковая цепь Arg82 стабилизируется водородными связями с молекулами воды Wat403, Wat404, Wat405 и Wat407 и играет ключевую роль в переносе протонов 28 , в OLPVRII эти молекулы воды заменены тремя боковыми цепями Asn, Asn69. , Asn184 и Asn188. Важно отметить, что Asn69 и Asn184 являются высококонсервативными, а Asn188 полностью консервативными среди вирусных родопсинов группы II (дополнительный рис. 1). Хотя было продемонстрировано, что Arg82 является ключевым элементом в механизме транслокации протонов в bR 28 , а аналогичный аргинин обнаружен в большинстве микробных родопсинов, играя важную роль в их функционировании 29,30,31 , мы предполагаем, что сильная стабилизация Arg72 в OLPVRII тремя боковыми цепями Asn может влиять на его подвижность и, таким образом, влиять на функцию родопсина. Разительно отличается от bR, где внеклеточная масса отделена от группы высвобождения протонов (пара Glu194 – Glu204), в OLPVRII большая полость выступает от поверхности внеклеточного белка до Arg72 (рис. 3), аналогично Exiguobacterium sibiricum родопсин (СОЭ) и ChR2 (дополнительный рисунок 15) 5,31 . Эта гидрофильная полость заполнена молекулами воды и образована полярными аминокислотами, включая Arg183, Asp124, Tyr73, Tyr187, Gln180 и Asn69 (дополнительный рис.14). Эти аминокислоты плотно связаны водородными связями, а также связаны сетью водородных связей с Arg72 и, кроме того, с RSB. Таким образом, в OLPVRII боковая цепь Arg72 разделяет область RSB и внеклеточное пространство. Депротонирование RSB во время фотоцикла запускает структурные перестройки во внеклеточной части белка и, следовательно, может приводить к открытию ворот вокруг Arg72 подобно тому, как это может происходить в ChR2 31 . Цитоплазматическая часть OLPVRII и ее связь с вестибюлемЦитоплазматическая часть OLPVRII резко отличается от таковой всех родопсинов с известной структурой (дополнительный рис.16). Во-первых, в цитоплазматической половине спирали C (Val86 в OLPVRII в положении Asp96 в bR) нет заряженной аминокислоты, которая действовала бы в качестве донора протонов для RSB во время фотоцикла, как в большинстве случаев протонной перекачки наружу. родопсины 32 . В OLPVRII предполагаемый донор протонов Glu42 расположен в спирали B (которая соответствует Thr46 в bR) и, таким образом, соседствует с RSB. Glu42 полностью консервативен в обеих группах вирусных родопсинов, и результаты сайт-направленного мутагенеза показывают, что Glu42 действительно служит донором протонов для RSB во время фотоцикла OLPVRII (см. Ниже). Механизм репротонирования Glu42 остается неясным. Скорее всего, захват протона происходит со стороны Glu26, где цитоплазматическая поверхность вогнута внутрь протомера и достигает боковой цепи Leu39, который является единственным остатком, отделяющим Glu42 от основной массы. В этом случае Leu39 может играть роль гидрофобных ворот, аналогичных Leu93 в bR 33 (дополнительный рис. 17). Альтернативный механизм репротонирования Glu42 может включать захват протона из части протомера, занятой Asn94, Asn158, Ser157, Tyr201 и Tyr206, которые вместе образуют небольшую гидрофильную полость вблизи цитоплазматической поверхности белка. Кроме того, между RSB и боковой цепью Glu42 наблюдается большая полость. Эта полость является гидрофильной и заполнена пятью молекулами воды, стабилизированными водородными связями с Glu42, Ser36 и атомами кислорода основной цепи Ser194 и Lys195 в спирали G. Аналогично расположенная полость существует в ChR2 дикого типа, но в этом случае полость является гидрофобный, без молекул воды внутри 31 . Водородные связи выходят из Glu42 через молекулы воды Wat7, Wat8, Wat9, Wat10, Wat11 и Thr23 и боковые цепи Thr43 к Leu39, который расположен рядом с Asn40 от границы раздела пентамеризации, усиливая взаимодействия между спиралями A и B внутри протомера (рис.4). Расстояния между Glu42 и Wat9, а также между Wat8 и Wat9 чрезвычайно короткие, всего 2,24 и 2,20 Å соответственно. Донор протонов Glu42, а также His37, Asn40 и Arg36, которые образуют пентамерные контакты в OLPVRII, все расположены в пределах двух витков одной и той же спирали B. Следовательно, в OLPVRII донор протонов жестко связан с интерфейсом олигомеризации не только геометрией. и ограничения α-спирали, но также и непрерывной цепочкой сильных водородных связей через молекулы воды в гидрофильной полости и полярные остатки спирали A.Более того, плотная сеть водородных связей пентамерных контактов выходит прямо на кольцо Arg29 (рис. 4). Рис. 4Цитоплазматическая часть OLPVRII и ее связь с центральной порой. а Вид сбоку. Один протомер скрыт для ясности. б Вид со стороны цитоплазмы. Центральная пора показана синей поверхностью. Красный кружок указывает на кольцо R29 центральной поры. Коричневые стрелки показывают предполагаемую последовательность структурных перестроек, передаваемых от RSB к границе раздела пор.Полости внутри белкового протомера окрашены в розовый цвет Эта отличная архитектура цитоплазматической части вместе с сохранением среди вирусных родопсинов группы II всех аминокислот, ответственных за пентамерную сборку OLPVRII, решительно поддерживает идею о том, что структурные изменения, инициированные изомеризацией сетчатки при поглощении света распространяются непосредственно от RSB к преддверию центральной поры. Действительно, расположение протонного донора Glu42 в спирали B, близко к RSB, его дополнительная водородная связь со спиралями A и G и интерфейсом олигомеризации, а также плотные межпротомерные контакты обеспечивают почти прямое взаимодействие между RSB и Arg29. , которые расположены на расстоянии 27 Å друг от друга (рис.4). Пора образована спиралями A и B, и, соответственно, ее форма и характеристики определяются сетью водородных связей, которая включает и стабилизирует эти спирали и зависит от конформации сетчатки (в частности, ориентации RSB). Более того, наша попытка уловить активное состояние белка в кристаллах привела к драматическому разрушению кристаллов (как в случае глазного родопсина 25 и сенсорного родопсина 34 ), очевидного по падению структурного разрешения с 2 до 20 Å.Этот факт указывает на то, что большие структурные перестройки происходят в OLPVRII во время фотоцикла, и дает еще один намек на канальную активность белка, где большие структурные перестройки происходят при открытии ионного канала, как в pLGICs 35 . Рис. 5Функциональная характеристика OLPVRII. a BLM измерения OLPVRII в E. coli полярных липидных липосомах. Красная кривая показывает электрический сигнал на BLM при освещении без добавления протонофора, а синяя кривая представляет сигнал при тех же условиях, но после добавления 1799 и монензина.Начало свечения обозначается серым (свет не горит) и белым (светится) цветами. b Эксперимент BLM с временным разрешением. Электрический сигнал (серый) был зарегистрирован после лазерной вспышки и соответствовал кривым экспоненциального затухания (красный). c Схематическое изображение кинетики фотоцикла OLPVRII, реконструированного в липосомы сои. Указаны времена жизни и максимумы поглощения промежуточных состояний. d Временные кривые изменения поглощения OLPVRII, измеренные при 410, 525 и 550 нм.Исходные данные представлены в виде файла исходных данных Таким образом, детальное изучение структуры OLPVRII не только предполагает, что этот родопсин является пентамерным светозависимым ионным каналом, но также демонстрирует возможные механизмы функционирования и регуляции канала. Чтобы проверить эти прогнозы, мы провели функциональные анализы OLPVRII и моделирование молекулярной динамики. Спектроскопическая характеристика OLPVRIIСпектры поглощения в УФ-видимой области OLPVRII, солюбилизированного в DDM, показывают максимальное поглощение связанного сетчатки при 514 нм.Положение пика смещается в зависимости от pH буфера. Мы изменили pH образца OLPVRII в буфере для шести смесей от 7,58 до 1,93 и от 7,58 до 12,93 для двух идентичных образцов. При щелочном pH максимальное поглощение сетчатки изменилось с 514 до 367 нм, что указывает на депротонирование RSB (дополнительный рис. 18). Сигмоидальная аппроксимация зависимости поглощения при 514 нм от pH дала pKa RSB 10,36 ± 0,08 (точка полураспада ± стандартная ошибка). Напротив, при кислом pH мы наблюдали постепенный сдвиг максимума поглощения сетчатки с 514 до 548 нм, что указывает на протонирование противоиона RSB Asp75.Мы аппроксимировали зависимость длины волны максимума поглощения от pH сигмоидальной кривой и получили pKa акцептора протонов 3,34 ± 0,11 (точка полураспада ± стандартная ошибка). Соответствующие значения pKa для bR составляют 12,2 и 2,7 36,37 , тогда как для PR pKa акцептора протонов составляет 7,68 38 . Эти заметные различия в pKa, по-видимому, связаны, во-первых, с отсутствием в вирусных родопсинах гистидина, который консервативен в PR и образует водородную связь с акцептором протона, таким образом уменьшая его pKa, и, во-вторых, аминокислотными заменами A215M и L93S. по сравнению с bR, которые изменяют свойства RSB. Кинетику фотоцикла мономерного OLPVRII, экспрессированного в штамме C41 и восстановленного в липидных пузырьках сои, измеряли при pH 7,5. Переходные изменения поглощения после лазерного освещения показаны на рис. 5d на трех характерных длинах волн: 410, 530 и 550 нм. Набор данных лучше всего описывается семью экспонентами. Общая длина фотоцикла OLPVRII составляет около 70 мс, и были получены соответствующие постоянные времени. Результаты глобального подбора показаны на рис. 5c. Фотоцикл включает микросекундную часть, которая обычно связана с высвобождением активированного иона, проявляющимся как депротонирование RSB, и миллисекундную часть восстановления основного состояния за счет поглощения протона. Как и в большинстве родопсинов, количество разрешенных кинетических промежуточных продуктов выше, чем количество спектрально различных состояний, что указывает на присутствие спектрально молчаливых переходов 39 . Типичные абсолютные спектры промежуточных соединений показаны на дополнительном рис.19. В микросекундной части фотоцикла депротонирование RSB происходит через серию промежуточных продуктов с характерными временами τ 1 = 1 мкс, τ 2 = 16 мкс, τ 3 = 61 мкс, а τ 4 = 340 мкс. Первый промежуточный продукт состоит из одного спектрального состояния, подобного K 540 , тогда как P2 − P4 представляет собой смесь состояний типа K 540 и M 410 . Появление смещенного в синий цвет интермедиата M 410 с депротонированным RSB в диапазоне сотен микросекунд типично для микробных родопсинов.Однако, в отличие от bR 39 и многих других родопсинов, OLPVRII не формирует L-состояние, то же самое было получено для протеородопсина 38 . Затем в миллисекундном диапазоне времени промежуточный продукт, подобный M 410 , возвращается в основное состояние ( τ 5 = 10 мс, τ 6 = 25 мс и τ 7 = 73 мс) через серию состояний равновесия P5-P7, которые состоят из смеси состояний M 410 -, N 525 – и O 550 -подобных состояний. Чтобы подтвердить предложенный механизм протонной перекачки, мы проанализировали мутанты E42Q и D75N солюбилизированного OLPVRII, в которых предполагаемые отрицательно заряженные донор и акцептор протонов, соответственно, заменены полярными остатками. Мутант E42Q показал фотоцикл, очень похожий на таковой у белка дикого типа, но со значительно более длительным распадом М-состояния, как в случае мутанта D96N bR 40 , в то время как D75N не смог сформировать М-состояние (дополнительный рис. 20).Эти эксперименты показывают, что Asp75 является акцептором протонов OLPVRII, как и многие другие микробные родопсины, тогда как Glu42 является донором протонов, который расположен в спирали B, что резко контрастирует с большинством известных родопсинов. Насосная активность OLPVRIIЧтобы определить, является ли вирусный родопсин ионным насосом, мы провели прямые измерения ΔpH для оценки перекачивающей активности OLPVRII в клетках E. coli и при восстановлении в липидных липосомах сои.В E. coli не было обнаружено изменения pH внешнего растворителя при освещении, что можно объяснить низким выходом белка. Затем мы измерили ΔpH с очищенным белком, восстановленным в липидные пузырьки, где была достигнута высокая плотность белка. Однако даже в этой системе и в отличие от эталонного белка (bR, экспрессируемого в E. coli , как сообщалось до 21 ) не было обнаружено изменения pH внешнего раствора при освещении (дополнительный рис.21). Электрогенные свойства OLPVRII были дополнительно исследованы в экспериментах с черной липидной мембраной (BLM), которые были разработаны, как описано ранее. 38 . Солюбилизированный OLPVRII был восстановлен в полярных липидах E. coli в буфере 100 мМ Tris-HEPES pH 7,4 (pH был выбран, потому что он близок к естественным условиям в Organic Lake 41 ). Без добавления протонофора при освещении протеолипосом непрерывным светом наблюдались характерные фототоки (рис.5а), где быстрое отклонение вверх сопровождается последующим медленным спадом обратно до нулевого тока, в то время как стационарный ток незначителен. Добавление ионофоров 1799 и монензина 38 не оказало качественного влияния на фототок, но наблюдался положительный стационарный ток, указывающий на непрерывную накачку (рис. 5а). Липосомы были приготовлены исключительно в буфере Tris-HEPES. Перенос этих ионов маловероятен, поэтому мы заключаем, что фототок переносится протонами. Чтобы исследовать, какие этапы транспортного цикла являются электрогенными, мы провели измерения BLM с временным разрешением (рис.5б). При лазерной вспышке быстрое отклонение вверх имеет две постоянные времени: τ 1 ′ = 24 мкс и τ 2 ′ = 6 мс, тогда как медленное затухание тока характеризуется двумя константами τ 3 ′ = 90 мс и τ 4 ′ = 950 мс. Сравнивая эти постоянные времени с константами, полученными в кинетических измерениях импульсного фотолиза, мы можем отнести постоянную времени τ 1 ′ к переходу K 540 → M 410 , а постоянные времени τ 2 ′ и τ 3 ′ отражают образование поздних O 550 – и N 525 -подобных промежуточных продуктов.Постоянная времени τ 4 ′ может коррелировать с неспецифическим разрядом BLM 38 . Принимая во внимание расположение функционально важных аминокислот в белке и результаты экспериментов BLM, мы заключаем, что, по крайней мере, в липосомах, OLPVRII действует как выходящий протонный насос. Однако перенос заряда за фотоцикл невелик по сравнению с ожидаемой активностью канала. В самом деле, было показано, что ChR2 также обладает активностью по перекачке протонов наружу, которая ничтожна по сравнению с его ионным каналированием 42 . Гидрофобный вентиль ионного канала OLPVRIIЧтобы проверить, может ли OLPVRII функционировать как пентамерный светозависимый ионный канал, мы провели неуправляемое моделирование многоатомной МД белка, встроенного в липидный бислой POPC в 200 мМ водном растворе NaCl. решение (дополнительный рис. 22). Мы наблюдали спонтанный поток воды в центральную пору с обеих сторон мембраны, за исключением гидрофобной области, занятой углеводородной цепью (рис. 6а). В отсутствие этих липидов непрерывный паттерн гидратации предполагает предполагаемый путь ионной проводимости (рис.6а, б) 43 . Однако при более внимательном рассмотрении обнаруживается минимум плотности воды в области пор, выстланной Phe24 и Leu28, где в наносекундном масштабе времени происходят всплески колебаний жидкость-пар. Расчет радиусов пор показывает сужение в этой области с минимумом 1,93 Å (рис. 6c). Рис. 6Моделирование методом МД показывает закрытый путь проникновения анионов. a Усредненные значения плотности воды в отсутствие или в присутствии углеводородной цепи вдоль центральной оси по данным неуправляемого моделирования методом МД с контуром 1.2 σ вид сбоку. Спирали A и B трех субъединиц показаны на рисунках, а остатки выстилки пор в виде палочек. b Na + (синий) и Cl – (красный) для мутантных белков WT и F24A-L28A в отсутствие детергента с контуром 0,2σ. c Радиусы пор вдоль оси пор (положения относительно центра масс белка) для конформационного ансамбля, отобранного при моделировании WT без углеводородной цепи. Показаны положения в боковой цепи остатков Phe24, Leu28 и Arg29. d Распределение электростатического потенциала, рассчитанное на основе моделирования методом МД, нанесенное на доступную для растворителя поверхность OLPVRII на виде сбоку. Два передних блока скрыты для наглядности. e Потенциал профилей средней силы для воды, проницаемости Na + и Cl – вдоль поры. f Гистограммы количества молекул воды, наблюдаемых в области гидрофобных ворот для различных мутаций. Исходные данные представлены в виде файла исходных данных Arg29 создает положительный электростатический потенциал на входе цитоплазматической поры, который отделен гидрофобным сужением от широкого, частично отрицательного, внеклеточного входа (рис.6d и дополнительный рис. 23). Профили свободной энергии, рассчитанные для воды, проникновения Na + и Cl –, показывают, что цитоплазматическая сторона поры является высоко анион-селективной, тогда как внеклеточный вход является катион-селективным, с самым высоким барьером, создаваемым Phe24 и Leu28 ( Рис. 6д). Уменьшение объемов соответствующих боковых цепей за счет замены аланина увеличивает гидратацию сужения, снижает энергетический барьер для проникновения ионов и приводит к общему Cl – -селективному ионному пути в двойном мутанте.Вдоль этого пути мы наблюдали множественные спонтанные события проникновения воды и Cl – (Рис. 6f и Дополнительный Рис. 23). Наши симуляции определяют OLPVRII как анионный канал, который стерически открыт, но функционально закрыт гидрофобными воротами, образованными Phe24 и Leu28. Важно отметить, что расчеты свободной энергии демонстрируют, что изменение заряда через мутацию R29E превращает OLPVRII в закрытый катион-селективный канал, который является проводящим в мутанте F24A / L28A / R29E, предполагая аналогичный механизм стробирования, что и в белке дикого типа (дополнительная информация Инжир.23). Таким образом, стробирование OLPVRII напоминает механизм гидрофобного стробирования в pLGIC и других биологических ионных каналах 24,35,44,45,46 . После индуцированной светом изомеризации сетчатки активация OLPVRII может распространяться от RSB через сеть водородных связей к спиралям, образующим поры. На дополнительном рис. 24 мы представляем простую механистическую модель активации анионного канала OLPVRII. Возникает соблазн предположить, что открытие канала связано с вращением или наклоном спирали A, которая перемещает Phe24 и Leu28 от поры и тем самым снижает эффективную гидрофобность в поре, вызывая смачивание гидрофобных ворот и позволяя проникать Cl –.В самом деле, подобное вращение α-спирали приводит к значительному смещению гидрофобных остатков (Phe80, Phe84 и Ile76) и открытию канала в хлоридном канале, управляемом пентамерным бестрофиновым лигандом, BEST1 35 (дополнительный рис. 25). % PDF-1.7
%
2442 0 объект
>
эндобдж
xref
2442 168
0000000016 00000 н.
0000005449 00000 н.
0000005722 00000 н.
0000005759 00000 п.
0000006358 00000 п.
0000006873 00000 н.
0000007029 00000 н.
0000007186 00000 н.
0000007343 00000 п.
0000007500 00000 н.
0000007657 00000 н.
0000007814 00000 н.
0000007971 00000 н.
0000008128 00000 н.
0000008284 00000 н.
0000008441 00000 п.
0000008598 00000 н.
0000008755 00000 н.
0000008912 00000 н.
0000009069 00000 н.
0000009226 00000 п.
0000009383 00000 п.
0000009539 00000 п.
0000009696 00000 п.
0000009853 00000 п.
0000010010 00000 п.
0000010165 00000 п.
0000010322 00000 п.
0000010479 00000 п.
0000010636 00000 п.
0000010793 00000 п.
0000010950 00000 п.
0000011107 00000 п.
0000011264 00000 п.
0000011421 00000 п.
0000011578 00000 п.
0000011735 00000 п.
0000011892 00000 п.
0000012049 00000 п.
0000012206 00000 п.
0000012363 00000 п.
0000012520 00000 п.
0000012677 00000 п.
0000012834 00000 п.
0000012991 00000 п.
0000013148 00000 п.
0000013305 00000 п.
0000013462 00000 п.
0000013619 00000 п.
0000013776 00000 п.
0000013933 00000 п.
0000014090 00000 п.
0000014247 00000 п.
0000014403 00000 п.
0000014560 00000 п.
0000014717 00000 п.
0000014874 00000 п.
0000015031 00000 п.
0000015188 00000 п.
0000015345 00000 п.
0000015502 00000 п.
0000016091 00000 п.
0000016604 00000 п.
0000016643 00000 п.
0000016693 00000 п.
0000016806 00000 п.
0000016921 00000 п.
0000017544 00000 п.
0000018225 00000 п.
0000018313 00000 п.
0000018756 00000 п.
0000019302 00000 п.
0000020473 00000 п.
0000021606 00000 п.
0000022180 00000 п.
0000022724 00000 п.
0000022820 00000 н.
0000023473 00000 п.
0000023561 00000 п.
0000024039 00000 п.
0000024650 00000 п.
0000025223 00000 п.
0000026293 00000 п.
0000027100 00000 н.
0000027931 00000 н.
0000028674 00000 п.
0000029448 00000 н.
0000032198 00000 п.
0000032981 00000 п.
0000035632 00000 п.
0000035701 00000 п.
0000041057 00000 п.
0000041126 00000 п.
0000045737 00000 п.
0000045806 00000 п.
0000051775 00000 п.
0000051844 00000 п.
0000056281 00000 п.
0000056350 00000 п.
0000059357 00000 п.
0000059426 00000 п.
0000059792 00000 п.
0000059861 00000 п.
0000059930 00000 н.
0000059999 00000 н.
0000060068 00000 п.
0000060137 00000 п.
0000060206 00000 п.
0000060275 00000 п.
0000060344 00000 п.
0000060413 00000 п.
0000060482 00000 п.
0000060551 00000 п.
0000060620 00000 п.
0000060689 00000 п.
0000060758 00000 п.
0000060827 00000 п.
0000060896 00000 п.
0000060965 00000 п.
0000061034 00000 п.
0000061103 00000 п.
0000061172 00000 п.
0000061241 00000 п.
0000061310 00000 п.
0000061379 00000 п.
0000061448 00000 п.
0000061517 00000 п.
0000062038 00000 п.
0000062107 00000 п.
0000062156 00000 п.
0000062225 00000 п.
0000097794 00000 п.
0000097863 00000 п.
0000097917 00000 п.
0000097986 00000 п.
0000098388 00000 п.
0000098457 00000 п.
0000098910 00000 п.
0000098979 00000 п.
0000099354 00000 н.
0000099423 00000 п.
0000099696 00000 п.
0000099765 00000 п.
0000100101 00000 п.
0000100170 00000 н. | ||||||
