
Микросхема LM324 – счетверенный операционный усилитель
Если в схеме нужно использовать сразу несколько операционных усилителей, а особых требований например по частоте, выходному току и т.п. нету, то LM324 прекрасный кандидат: в 14 выводном корпусе размещены 4 операционных усилителя общего применения с общим питанием.
Операционные усилители серии LM324 выпускаются несколькими производителями и параметры микросхем от производителя к производителю могут отличаться. Так же разные производители выпускают модификации серии на разные температурные диапазоны и в разных корпусах:
- для монтажа в отверстия: DIP14;
- для поверхностного монтажа: SO-14, TSSOP-14, QFN16 3×3;
- для расширенного температурного диапазона в керамических корпусах.
Например все эти операционные усилители модификации LM324: LM324A, LM324E, LM124, LM224, LM2902, LM2902E, LM2902V, NCV2902.
Характеристики LM324:
- широкий диапазон питающих напряжений: от 3 до 30В;
- может работать как при однополярном, так и при двуполярном питании;
- большой коэффициент усиления по напряжению: 100дБ;
- широкий частотный диапазон: 1,3МГц;
- низкий потребляемый ток на усилитель: 375мкА;
- низкий входной ток смещения: 2нА;
- низкое входное напряжение смещения, максимум: 5мВ;
- не требует внешних цепей частотной коррекции;
- диапазон входных напряжений от 0 В.
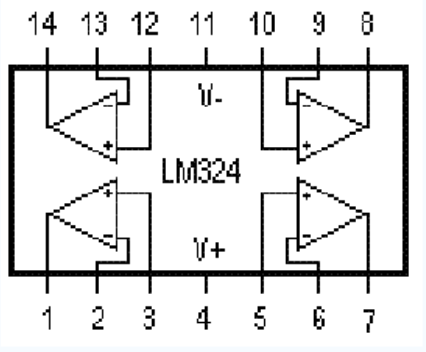
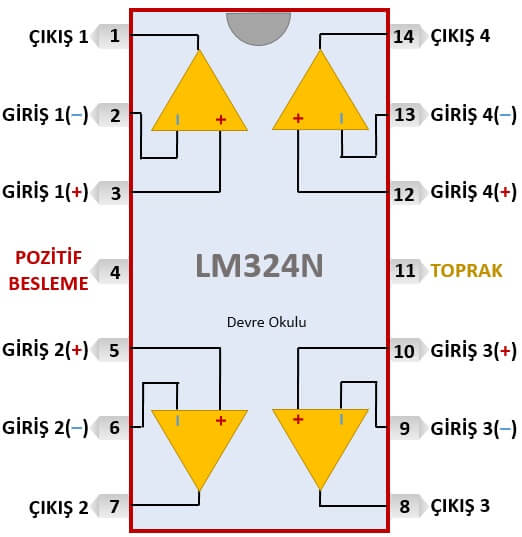
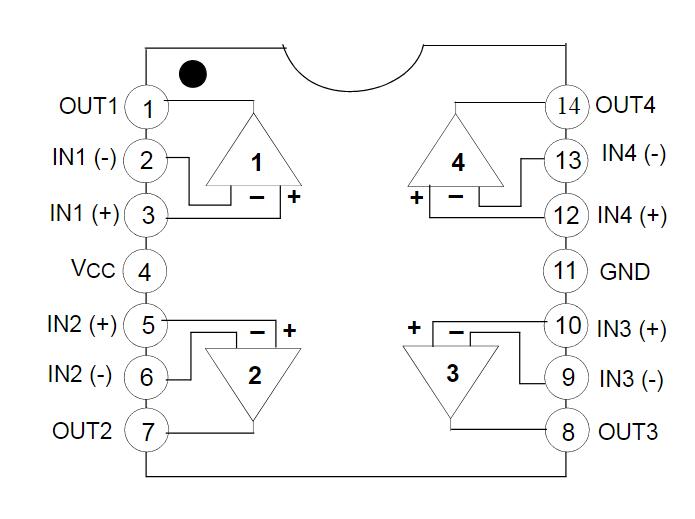
Цоколевка LM324 в DIP-14, SO-14, TSSOP-14.
Внутренняя структура одного канала:
LM324 схемы включения
Итак, где же предлагает использовать LM324 Texas Instruments:
- DVD и блюрей приводы,
- Домашние кинотеатры,
- Различные датчики,
- Мультиметры и осцилографы,
- Управление различными двигателями,
- Телевизоры,
- Весы.
Кстати TI выпускает 324-тые уже более 40 лет – с 1975.
Большое количество операционных усилителей может понадобиться как для схем с большим количеством однотипных каналов, так и в сложных схемах.
LM324 — Четырехканальный операционный усилитель — DataSheet
LM124, LM224, LM324Особенности
- Широкая полоса пропускания: 1.3 МГц
- Большое усиление по постоянному току: 100 дБ
- Широкий диапазон напряжения питания:
- Для однополярного питания: 0т +3 В до +30 В
- Для двухполярного питания: от ±1.
 5 В до ±15 В
5 В до ±15 В - Диапазон синфазного напряжения включает землю
- Большая амплитуда выходного напряжения: от 0 В до VCC -1.5 В
- Выходная мощность подходит для работы от батареи
Купить LM324
Описание
Микросхемы серии LM124, LM224 и LM324 состоят из четырех операционных усилителей с высоким коэффициентом усиления, которые работают от одного источника питания. Областью их применения являются усилители-преобразователи, усилители о все обычные схемы применения ОУ , которые можно подключить к одному источнику питания.
Тип корпуса DIP14 | Тип корпуса SO-14 |
Тип корпуса TSSOP-14 | Тип корпуса QFN16 3×3 |
Цоколевка LM324 |
Цоколевка корпус QFN16 вид сверху |
Внутренняя схема ОУ
| Обозначение | Параметр | LM124 | LM224 | LM324 | Ед. изм. изм. | ||
| VCC | Напряжение питания | ±16 или 32 | В | ||||
| Vin | Входное напряжение | от -0.3 до 32 | В | ||||
| Vid | Дифференциальное входное напряжение | 32 | В | ||||
| Длительность короткого замыкания | Неограничена | ||||||
| Iin | Входной ток: Vin отрицательное | 5 мА при постоянном токе и 5 мА при переменном(коэффициент заполнения = 10%, T=1 с) | мА | ||||
| Входной ток: Vin положительное | 0.4 | ||||||
| Toper | Диапазон рабочих температур | от -55 до +125 | от -40 до +105 | от 0 до +70 | °C | ||
| Tj | Максимальная температура p-n переходов | 150 | °C | ||||
Электрические характеристики
| Обозначение | Параметр | Мин.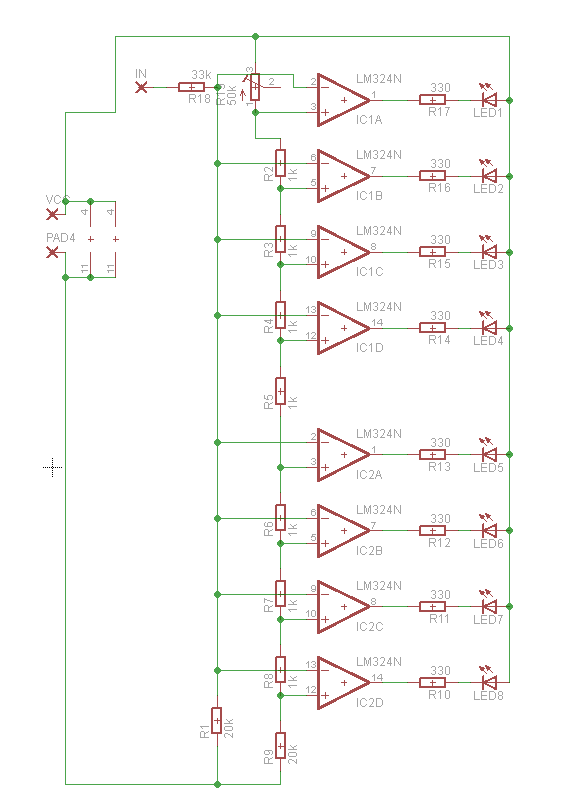 | Тип. | Макс. | Ед. изм. |
| Vio | Входное напряжение компенсации смещения нуля на выходе Tamb = +25° C LM124-LM224 | 2 | 5 | мВ | |
| LM324 | 7 | ||||
| Tmin ≤ Tamb ≤ Tmax LM124-LM224 | 7 | ||||
| LM324 | 7 | ||||
| Iio | Входной ток компенсации смещения нуля на выходе Tamb = +25° C | 2 | 30 | нА | |
| Tmin ≤ Tamb ≤ Tmax | 100 | ||||
| Iib | Входной ток смещения Tamb = +25° C | 20 | 150 | нА | |
| Tmin ≤ Tamb ≤ Tmax | 300 | ||||
| Avd | Максимальное усиление сигнала по напряжению VCC+ = +15 В, RL = 2 кОм, Vo = от 1. 4 В до 11.4 В Tamb = +25° C 4 В до 11.4 В Tamb = +25° C | 50 | 100 | В/мВ | |
| Tmin ≤ Tamb ≤ Tmax | 25 | ||||
| SVR | Коэффициент подавления помех по питанию Rs ≤ 10 кОм) VCC+ = от 5 В до 30 В Tamb = +25° C | 65 | 100 | дБ | |
| Tmin ≤ Tamb ≤ Tmax | 65 | ||||
| ICC | Потребляемый ток для всех усилителей без нагрузки Tamb = +25° C VCC = +5 В | 0.7 | 1.2 | мА | |
| VCC = +30 В | 1.5 | 3 | |||
| Tmin ≤ Tamb ≤ Tmax VCC = +5 В | 0.8 | 1.2 | |||
| VCC = +30 В | 1.5 | 3 | |||
| Vicm | Диапазон синфазного входного напряжения VCC = +30 В, Tamb = +25° C | 0 | VCC-1. 5 5 | В | |
| Tmin ≤ Tamb ≤ Tmax | 0 | VCC-2 | |||
| CMR | Коэффициент ослабления синфазного входного напряжения Rs ≤ 10 кОм) Tamb = +25° C | 70 | 80 | дБ | |
| Tmin ≤ Tamb ≤ Tmax | 60 | ||||
| VOH | Высокий уровень выходного напряжения VCC = +30 В, Tamb = +25° C, RL = 2 кОм | 26 | 27 | В | |
| Tmin ≤ Tamb ≤ Tmax | 26 | ||||
| Tamb = +25° C, RL = 10 кОм | 27 | 28 | |||
| Tmin ≤ Tamb ≤ Tmax | 27 | ||||
| VCC = +5 В, RL = 2 кОм Tamb = +25°C | 3. | ||||
| Tmin ≤ Tamb ≤ Tmax | 3 | ||||
| VOL | Низкий уровень выходного напряжения (RL = 10 кОм) Tamb = +25°C | 5 | 20 | мВ | |
| Tmin ≤ Tamb ≤ Tmax | 20 | ||||
| SR | Скорость нарастания сигнала VCC = 15 В, Vi = от 0.5 до 3 В, RL = 2 кОм, CL = 100 пФ, единичное усиление | о.4 | В/мкс | ||
| GBP | Полоса пропускания VCC = 30 В, f = 100 кГц, Vin = 10 мВ, RL = 2 кОм, CL = 100 пФ | 1.3 | МГц | ||
| THD | Суммарный коэффициент гармонических искажений f = 1 кГц, Av = 20 дБ, RL = 2 кОм, Vo = 2 Vpp, C L = 100 пФ, VCC = 30 В | 0.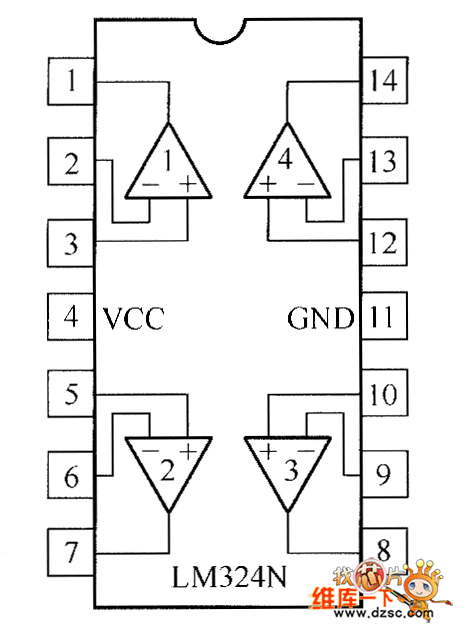 015 015 | % | ||
| Vo1/Vo2 | Разделение каналов 1 кГц ≤ f ≤ 20 кГц | 120 | дБ |
Применение (схемы включения)
Инвертирующий усилитель переменного тока | Усилитель постоянного тока с регулируемым коэффициентом усиления |
Неинвертирующий усилитель переменного тока | Суммирующий усилитель постоянного тока |
Неинвертирующий усилитель постоянного тока | Активный полосовой фильтр |
Дифференциальный усилитель постоянного тока | Использование симметричного усилителя для уменьшения входного тока |
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите
Микросхема LM324 – счетверенный операционный усилитель
- Подробности
- Категория: Components
- Просмотров: 8313
Если в схеме нужно использовать сразу несколько операционных усилителей, а особых требований например по частоте, выходному току и т. п. нету, то LM324 прекрасный кандидат: в 14 выводном корпусе размещены 4 операционных усилителя общего применения с общим питанием.
п. нету, то LM324 прекрасный кандидат: в 14 выводном корпусе размещены 4 операционных усилителя общего применения с общим питанием.
Операционные усилители серии LM324 выпускаются несколькими производителями и параметры микросхем от производителя к производителю могут отличаться. Так же разные производители выпускают модификации серии на разные температурные диапазоны и в разных корпусах:
- для монтажа в отверстия: DIP14;
- для поверхностного монтажа: SO-14, TSSOP-14, QFN16 3×3;
- для расширенного температурного диапазона в керамических корпусах.
Например все эти операционные усилители модификации LM324: LM324A, LM324E, LM124, LM224, LM2902, LM2902E, LM2902V, NCV2902.
Характеристики LM324:
- широкий диапазон питающих напряжений: от 3 до 30В;
- может работать как при однополярном, так и при двуполярном питании;
- большой коэффициент усиления по напряжению: 100дБ;
- широкий частотный диапазон: 1,3МГц;
- низкий потребляемый ток на усилитель: 375мкА;
- низкий входной ток смещения: 2нА;
- низкое входное напряжение смещения, максимум: 5мВ;
- не требует внешних цепей частотной коррекции;
- диапазон входных напряжений от 0 В.
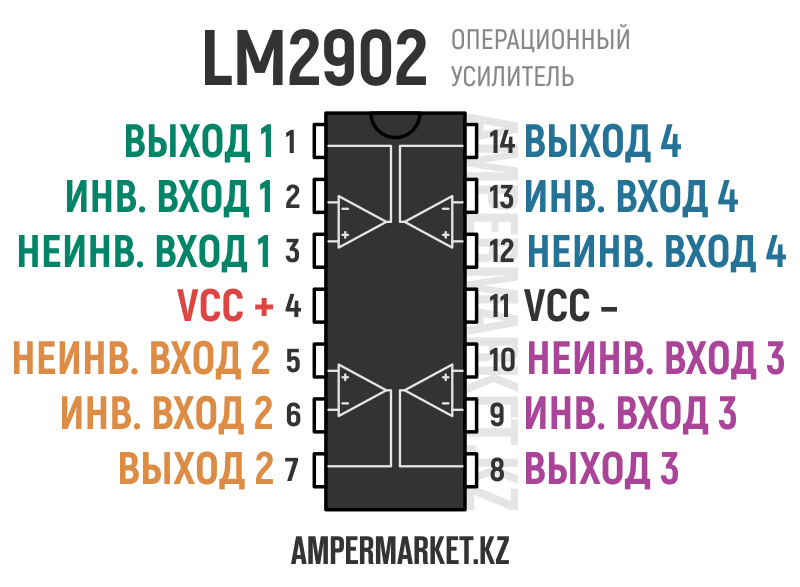
Цоколевка LM324 в DIP-14, SO-14, TSSOP-14.
Внутренняя структура одного канала:
LM324 схемы включения
Итак, где же предлагает использовать LM324 Texas Instruments:
- DVD и блюрей приводы,
- Домашние кинотеатры,
- Различные датчики,
- Мультиметры и осцилографы,
- Управление различными двигателями,
- Телевизоры,
- Весы.
Кстати TI выпускает 324-тые уже более 40 лет – с 1975.
Большое количество операционных усилителей может понадобиться как для схем с большим количеством однотипных каналов, так и в сложных схемах.
Например счетверенный LM324 пригодятся как ни кстати в схеме биквадратного фильтра.
Микросхема LM324 DIP-14 (арт. 61/50/522)
Артикул: 61/50/522
1Микросхема LM324 DIP-14 (арт. 61/50/522)
61/50/522)
LM324 – Справочник по микросхемам
Назначение, описание, аналоги
Категория Микросхемы серии L
Микросхемы серии LM124/ LM224/ LM324/ LM2902 являются недорогими операционным усилителями, имеющими прямой дифференциальный вход, внутричастотную компенсацию при единичном усилении и защиту от короткого замыкания.
В одном корпусе микросхемы расположено четыре независимых друг от друга операционных усилителя. У них имеется ряд неоспоримых преимуществ по сравнению с типовыми операционными усилителями, применяемыми в схемах с однополярным питанием. ОУ LM324 отлично работает в широком диапазоне напряжения питания: от 3 В до 32 В. Микросхема производится в корпусах типа SOIC и DIP.
Технические данные операционного усилителя LM324
- Напряжение питания:
- — однополярное: 3…32 В.
- — двухполярное: 1,5…16 В.
- Усиление по постоянному напряжению: 100 дБ.
- Собственный ток потребления: 700 мкА.
- Входной ток смещения (с температурной компенсацией): 45 нА.
- Входное напряжение смещения: 2 мВ.
- Диапазон входного синфазного напряжения содержит землю.
- Дифференциальный диапазон входного напряжения достигает напряжения питания.
- Выходного напряжение: от 0 до Uпит. – 1,5 В.
Структура операционного усилителя
Назначение выводов
Аналоги LM324
Список зарубежных и отечественных аналогов LM324:
- ULN4336N
- GL324
- LA6324
- IR3702
- HA17324
- MB3614
- NJM2902D
- SG324N
- TDB0124
- UA324
- TA75902P
- 1401УД2 (отечественный аналог)
- 1435УД2 (отечественный аналог)
Схема включения LM324
Инвертирующий усилитель по переменному току
В данном варианте усилителя коэффициент усиления будет равен: k = — R3/R1
Неинвертирующий усилитель по переменному току
Коэффициент усиления у данного типа усилителя рассчитывается по следующей формуле: k = 1 + R4/R1
Неинвертирующий усилитель постоянного тока
Усиление равно: k = 1 + R3/R2
Пиковый детектор на LM324
Пиковые детекторы используются для фиксации максимальной, за определенный промежуток времени, величины сигнала
Компаратор на LM324 с гистерезисом
Разница значений входного напряжения, при котором происходит переключение выхода компаратора (гистерезис) из одного состояния в другое, рассчитывается по следующей формуле: Н = (R1/(R1+R2))(Voh-Vol)
Примеры использования операционного усилителя LM324
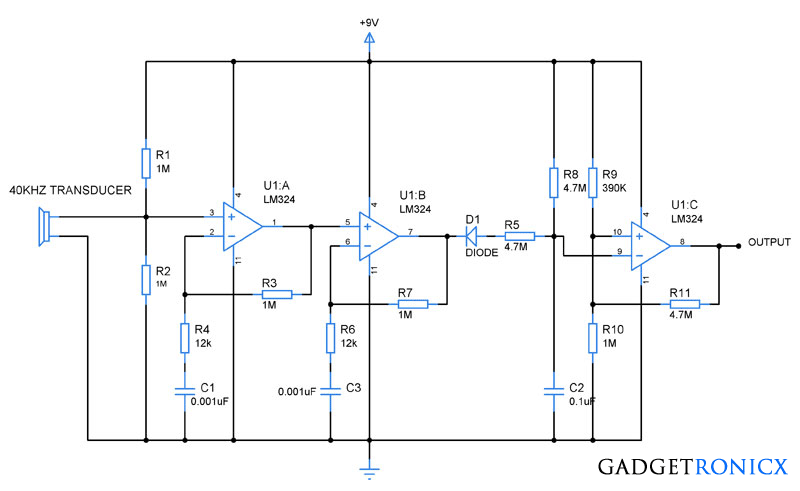
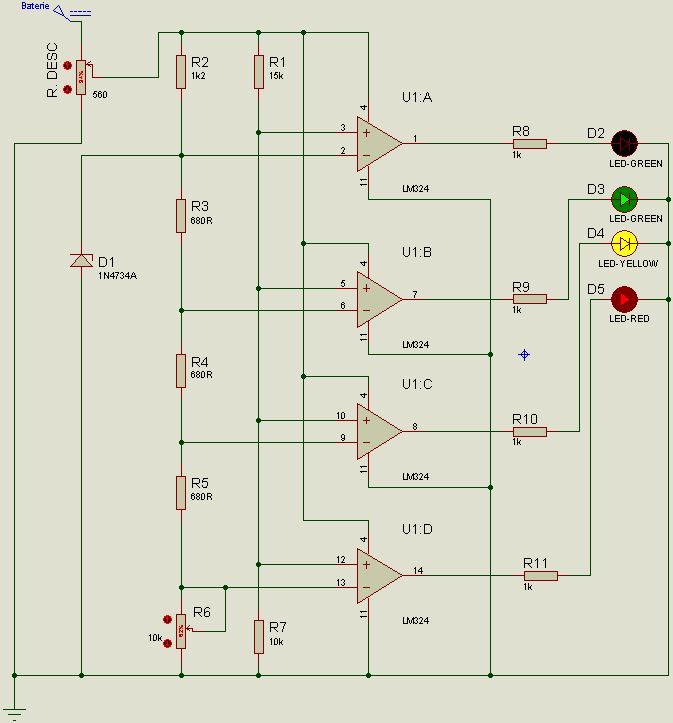
Светодиодный индикатор акустического сигнала на LM324
Низкочастотный сигнал с выхода усилителя подается на инвертирующие входы всех операционных усилителей LM324.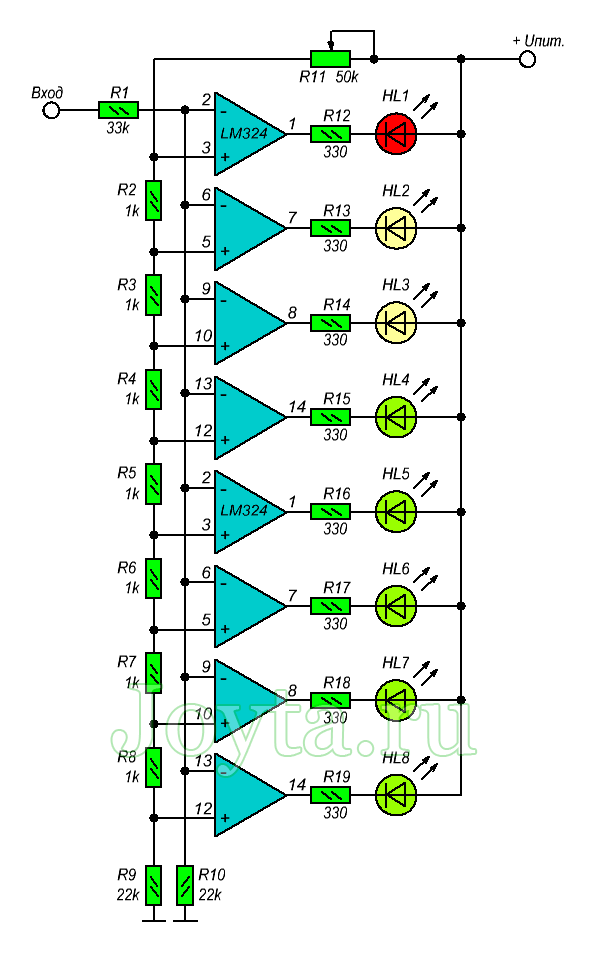 Прямые входы их подключены к делителю напряжения построенного из цепи постоянных резисторов R2…R9. Переменным резистором можно выставить необходимую чувствительность светодиодного индикатора. Сопротивления R12…R19 ограничивают максимальный ток, протекающий через светодиоды.
Прямые входы их подключены к делителю напряжения построенного из цепи постоянных резисторов R2…R9. Переменным резистором можно выставить необходимую чувствительность светодиодного индикатора. Сопротивления R12…R19 ограничивают максимальный ток, протекающий через светодиоды.
Простая светодиодная мигалка на ОУ LM324
Схема позволяет плавно поочередно включать и выключать светодиоды. Светодиодная мигалка построена на операционном усилителе LM324 и двух транзисторах разной проводимости. От сопротивления резистора R3 и емкости конденсатора C1 зависит скорость переключения светодиодов.
Микрофонный усилитель
Данная схема предназначена для усиления слабого сигнала электретного микрофона. Схема микрофонного усилителя представляет собой инвертирующий усилитель по переменному току с коэффициентом усиления 220 (R5/R3).
Даташит на микросхему
Источник: http://www.joyta.ru/
Микросхема LM324: назначение выводов, характеристики, аналоги
4-канальный операционный усилитель LM324 с однополярным питанием состоит из четырех независимых операционных усилителей и дифференциальными входами.
Базовая схема включения предполагает работу от однополярного источника питания, однако возможно подключение микросхемы к двухполярному питанию. Микросхема устойчиво работает от источника малой мощности в широком диапазоне напряжений. Сила тока потребления зависит величины напряжения источника питания.
Особенности
- Внутренняя частотная компенсация.
- Внутренняя защита выходов от короткого замыкания.
- Большой коэффициент усиления по постоянному напряжению – 100 дБ.
- Широкая полоса пропускания (единичный коэффициент усиления) –1 МГц.
- (Температурная компенсация).
- Широкий Диапазон Питания: однополярное питание – 3…32 В; двухполярное питание от ±(1,5…16) В.
- Малый ток потребления – 700 мкА (практически не зависит от напряжения питания).
- Низкий входной ток смещения – 45 нА (благодаря температурной компенсации).
- Низкие входные дифференциальные напряжение смещения – 2 мВ и ток смещения – 5 нА.
- Диапазон дифференциального входного напряжения определяется величиной напряжения питания.
Корпуса
Назначение выводов
Микросхема LM324 имеет 14 выводов. Назначение выводов для разных корпусов идентичное:
- 2,3, 5,6, 9,10, 13,12 — вход;
- 1,7,8,14 – выход;
- 4 – плюс источника питания;
- 11 – минус источника питания.
Предельно допустимые значения
Данные в таблице действительны при температуре воздуха 25°С.
| Параметр | Обозн. | Величина | Ед. изм. |
|---|---|---|---|
| Напряжение питания | Vdc | ||
| однополярное | VCC | 32 | |
| двухполярное | VCC, VEE | ±16 | |
| Диапазон входных дифференциальных напряжений | VIDR | ±32 | Vdc |
| Диапазон входного синфазного напряжения | VICR | −0,3…32 | Vdc |
| Длительность короткого замыкания на выходе | tSC | Постоянно | |
| Температура кристалла | TJ | 150 | °C |
| Тепловое сопротивление, кристалл-воздух | RθJA | °C /W | |
| Case 646 | 118 | ||
| Case 751A | 156 | ||
| Case 948G | 190 | ||
| Диапазон температур хранения | Tstg | −65…+150 | °C |
| Диапазон рабочих температур LM324, LM324A, LM324E | TA | 0…+70 | °C |
ESD – защита от электростатического разряда
- HBM (модель человеческого тела – имитирует контакт с человеком) – 2000 V.

- ММ (модель машины – имитирует контакт с оборудованием) – 200 V.
Таблица электрических параметров
Данные в таблице действительны при VCC=5.0 V, VEE=GND, TA=25°C.
| Параметр | Обозн. | Мин. | Тип. | Макс. | Ед. изм. |
|---|---|---|---|---|---|
| Разница входных напряжений смещения | VIO | mV | |||
| VCC = 5,0…30 V | |||||
| TA = 25°C | 2 | 7 | |||
| TA = Thigh | 9 | ||||
| TA = Tlow | 9 | ||||
| Средний температурный коэффициент VIO | ΔVIO/ΔT | − | 7 | − | µV/°C |
| TA = Thigh…Tlow | |||||
| Разница входных токов смещения | IIO | − | 5 | 50 | nA |
| TA = Thigh…Tlow | − | − | 150 | ||
| Средний температурный коэффициент IIO | ΔIIO/ΔT | − | 10 | − | pA/°C |
| TA = Thigh…Tlow | |||||
| Входной ток смещения | IIB | − | −90 | −250 | nA |
| TA = Thigh…Tlow | −500 | ||||
| Диапазон входного синфазного напряжения | VICR | V | |||
| TA = +25°C | 0 | 28,3 | |||
| TA = Thigh…Tlow | 0 | 28 | |||
| Диапазон входного дифференциального напряжения | VIDR | − | − | VCC | V |
| Коэффициент усиления большого сигнала по без обратной связи | AVOL | V/mV | |||
RL = 2. 0 kΩ, VCC = 15 V, 0 kΩ, VCC = 15 V, | 25 | 100 | − | ||
| TA = Thigh…Tlow | 15 | ||||
| Разделение входных каналов при 10…20 kHz | CS | − | −120 | − | dB |
| Коэффициент подавления синфазного сигнала, при RS менее 10 kΩ | CMR | 65 | 70 | − | dB |
| Коэффициент подавления помех источника питания | PSR | 65 | 100 | − | dB |
| Максимальное выходное напряжение | VOH | V | |||
| VCC = 5 V | 3,3 | 3,5 | |||
| VCC = 30 V | 27 | 28 | |||
| Минимальное выходное напряжение VCC = 5 V | VOL | 5 | 20 | mV | |
| Выходной ток VCC = 15 V, ТА=25°С | IO + | 20 | 40 | mA | |
| Выходной ток (нагрузка подключена к источнику питания) VCC = 15 V, TA = 25°C | IO − | 10 | 20 | mA | |
| Выходной ток короткозамкнутой нагрузки на землю | ISC | 40 | 60 | mA | |
| Ток источника питания при VCC = 30 V VO = 0 V | 3 | mA |
Внутренняя принципиальная схема 1-го канала ИМС LM324
LM324 содержит 4 операционных двухступенчатых усилителя с частотной компенсацией. Первый каскад – входной дифференциальный, собран на элементах Q20 и Q18 с буферными транзисторами Q21 и Q17 и дифференциального преобразователя на Q3 и Q4. Первая ступень не только усиливает входные сигналы, она определяет уровни сдвига сигналов и нормализует характеристику крутизны. Такое схемное решение позволяет применить в компенсационной линии конденсатор очень малой емкости – 5,0 пФ), что увеличивает эффективность использования полезной площади кристалла. Дифференциальный каскад на транзисторах с разделенными коллекторами Q20 и Q18 преобразует входные напряжения в ток. Другая особенность этой ступени в том, что при питании усилителя однополярным напряжением не происходит насыщения транзисторов дифференциального каскада.
Первый каскад – входной дифференциальный, собран на элементах Q20 и Q18 с буферными транзисторами Q21 и Q17 и дифференциального преобразователя на Q3 и Q4. Первая ступень не только усиливает входные сигналы, она определяет уровни сдвига сигналов и нормализует характеристику крутизны. Такое схемное решение позволяет применить в компенсационной линии конденсатор очень малой емкости – 5,0 пФ), что увеличивает эффективность использования полезной площади кристалла. Дифференциальный каскад на транзисторах с разделенными коллекторами Q20 и Q18 преобразует входные напряжения в ток. Другая особенность этой ступени в том, что при питании усилителя однополярным напряжением не происходит насыщения транзисторов дифференциального каскада.
Второй каскад — это стандартный усилительный каскад. Для нагрузки он является источником тока.
Все 4 усилителя на рабочие режимы выводятся одним узлом смещения. Благодаря этому каждый усилитель обладает хорошими показателями температурной стабильности и подавления шумов по линии питания.
Импортные и отечественные аналоги
ИМС LM324 широко применяется в радиолюбительских разработках и электронных устройствах радиотехнический промышленности. Ее отличительные особенности, – наличие дифференциальных входов и высокий коэффициент усиления используется при конструировании различных электронных схем повышенной функциональности: интегрирующих, дифференцирующих, модулирующих узлах и блоках, а также в сумматорах и вычитателях. Это только небольшая часть областей применения LM324. Кроме того, промышленность постоянно выпускает новые приборы, в которых используется данная ИМС.
Естественно, что производители электронных радиокомпонентов предлагают большой перечень микросхем-операционных усилителей, которые можно использовать для замены LM324.
| Производители | Аналоги |
|---|---|
| Импортные | ULN4336N, GL324, LA6324, IR3702, HA17324, MB3614, NJM2902D, SG324N, TDB0124, UA324, TA75902P |
| Отечественные | 1401УД2, 1435УД2 |
Типовые эксплуатационные характеристики
Зависимость входного напряжения от напряжения питания.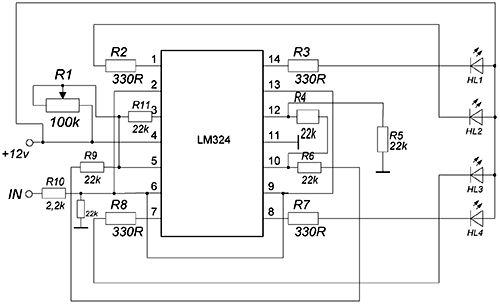
Зависимость коэффициента усиления большого сигнала с разомкнутой обратной связью от частоты.
Зависимость выходного напряжения от частоты.
График отклика выходного сигала на входной импульс.
Зависимость тока потребления от напряжения питания.
Зависимость входного тока смещения от напряжения питания.
Операционный усилитель LM324
Описание
LM324 состоит из четырех независимых, высоким коэффициентом усиления, внутренняя частотная компенсация операционных усилителей. Они работают с единым источником питания в широком диапазоне напряжений. Работа от Сплита питания также возможно и низким напряжением питания ток утечки не зависит от величины питания напряжение.
Основные параметры
| Каналов,шт | 4 |
VOS (тип. ),мВ ),мВ | 7 |
| IBIAS (тип.),нА | 250 |
| Полоса пропускания (тип.),МГц | 1 |
| Slew Rate (тип.),В/мкс | 0.5 |
| CMRR (тип.),дБ | 85 |
| Gain (тип.),дБ | 100 |
| Shutdown | Нет |
| VCC,В | от 3 до 32 |
| ICC на канал (макс.),мА | 1. |
| TA,°C | от 0 до 70 |
| Корпус | CDIP-14 LCC-20 Wafer |
 2
2
Параметры LM324 смотреть в PDF.
5962-01-324-8055 – МИКРОСХЕМА В СБОРЕ, 63A141497P006, 63D720298G1 PC 44, 63D720298G1PC44
×
Группа 85: Электрические машины, оборудование и их части; Звукозаписывающие и воспроизводящие устройства, устройства записи и воспроизведения телевизионного изображения и звука, а также их части и принадлежности
| Номер приложения Б и заголовки | Описание товара | Кол-во единиц | |
|---|---|---|---|
| 85.42 | – Электронные интегральные схемы; их части: | ||
| – – Электронные интегральные схемы: | |||
8542. 31.0000 31.0000 | – – – Процессоры и контроллеры, совмещенные или не совмещенные с запоминающими устройствами, преобразователями, логическими схемами , усилители, тактовые и временные схемы или другие схемы | No. | |
| 8542.32 | – – – Воспоминания: | ||
| – – – – Динамический произвольный доступ для чтения и записи: | |||
| 8542.32.0015 | – – – – – Не более 1 гигабита | No. | |
| 8542.32.0023 | – – – – – Более 1 гигабита | No. | |
| 8542.32.0040 | – – – – Статическое чтение-запись с произвольным доступом (SRAM) | No. | |
| 8542.32.0050 | – – – – Электрически стираемая программируемая постоянная память (EEPROM) | No. | |
| 8542.32.0060 | – – – – Стираемая (кроме электрически) программируемая постоянная память (EPROM) | № | |
8542. 32.0070 32.0070 | – – – – Другое | № | |
| 8542.33.0000 | – – – Усилители | No. | |
| 8542.39.0000 | – – – Другое | No. | |
| 8542.90.0000 | – – – Детали | X | |
5962-00-324-0368 – ЦИФРОВАЯ МИКРОСХЕМА, MC1660Sh3, MC1660SP2, SC83803
×
Группа 85: Электрические машины, оборудование и их части; Звукозаписывающие и воспроизводящие устройства, устройства записи и воспроизведения телевизионного изображения и звука, а также их части и принадлежности
| График Б №и товарные позиции | Описание товара | Кол-во единиц | |
|---|---|---|---|
| 85,42 | – Схемы электронные интегральные; их части: | ||
| – – Электронные интегральные схемы: | |||
8542. 31.0000 31.0000 | – – – Процессоры и контроллеры, совмещенные или не совмещенные с запоминающими устройствами, преобразователями, логическими схемами , усилители, тактовые и временные схемы или другие схемы | No. | |
| 8542.32 | – – – Воспоминания: | ||
| – – – – Динамический произвольный доступ для чтения и записи: | |||
| 8542.32.0015 | – – – – – Не более 1 гигабита | No. | |
| 8542.32.0023 | – – – – – Более 1 гигабита | No. | |
| 8542.32.0040 | – – – – Статическое чтение-запись с произвольным доступом (SRAM) | No. | |
| 8542.32.0050 | – – – – Электрически стираемая программируемая постоянная память (EEPROM) | No. | |
| 8542.32.0060 | – – – – Стираемая (кроме электрически) программируемая постоянная память (СППЗУ) | № | |
8542.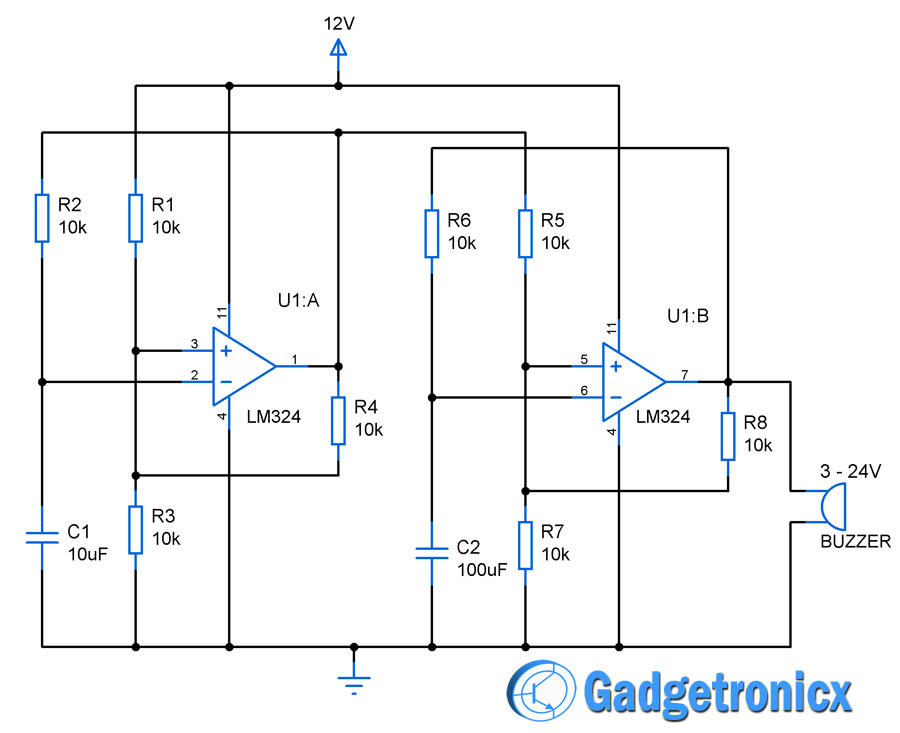 32.0070 32.0070 | – – – – Другое | Нет . | |
| 8542.33.0000 | – – – Усилители | № | |
| 8542.39.0000 | – – – Прочие | № | |
| 8542.90.0000 | – – – Детали | X | |
NSN 5962-01-324-6608 [наличие запчастей]
Особенности и характеристики
Общая ширина
Между 0.745 дюймов и 1,025 дюйма
Ширина корпуса
От 0,245 дюйма до 0,285 дюйма
Высота корпуса
От 0,045 дюйма до 0,085 дюйма
Максимальное рассеивание мощности
218,0 милливатт
Диапазон рабочих температур
-55.0 / + 125,0 градусов Цельсия
Диапазон температур хранения
-65,0 / + 165,0 градусов Цельсия
Предоставляемые функции
Монолитный и со стробоскопом
Материал приложения
Керамика
Конфигурация шкафа
Пакет плоский
Выходная логическая форма
Эмиттерная логика
Шаблон входной цепи
5 вход
Конструктивное назначение и количество
4 ворота, ор-ни
Краткое описание дела Источник и обозначение
Ф-5 мил-м-38510
Текущий рейтинг по характеристике
265. 00 наноампер обратный ток, постоянный микроампер
00 наноампер обратный ток, постоянный микроампер
Терминальная обработка поверхности
Припой
Название детали присвоено контролирующим агентством
Ecl, quad или / или ворота
Номинальное напряжение и тип согласно характеристике
Источник питания 8,0 вольт
Документ с данными испытаний
Стандарт 96906-mil-std-883 (включает отраслевые стандарты или стандарты ассоциации, стандарты отдельных производителей и т. Д.).
Тип и количество клемм
16 плоских выводов
FSC
5962 Электронные микросхемы
Цифровая микросхема НСН 5962-01-324-6609 – Электронные микросхемы
Инвентарный номер НАТО
5962-01-324-6609
ДЕМИЛ
Да – контролируется ИТАР (код D)
Общая ширина
Между 0.745 дюймов и 1,025 дюйма
Длина корпуса
0,440 дюймов
Ширина корпуса
От 0,245 дюйма до 0,285 дюйма
Высота корпуса
От 0,045 дюйма до 0,085 дюйма
Максимальное рассеивание мощности
250,0 милливатт
Диапазон рабочих температур
-55.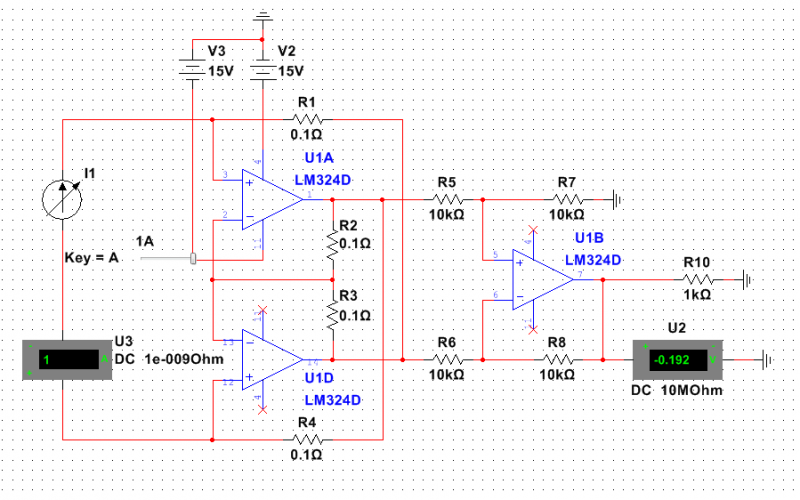 0 / + 125,0 градусов Цельсия
0 / + 125,0 градусов Цельсия
Диапазон температур хранения
-65,0 / + 165,0 градусов Цельсия
Предоставляемые функции
Монолитный
Материал приложения
Керамика
Конфигурация шкафа
Пакет плоский
Выходная логическая форма
Эмиттерная логика
Шаблон входной цепи
Четыре входа 2
Конструктивное назначение и количество
4 ворота, и
Краткое описание дела Источник и обозначение
Ф-5 мил-м-38510
Терминальная обработка поверхности
Припой
Номинальное напряжение и тип согласно характеристике
8.Источник питания 0 вольт
Документ с данными испытаний
Стандарт 96906-mil-std-883 (включает отраслевые стандарты или стандарты ассоциации, стандарты отдельных производителей и т. Д.).
Тип и количество клемм
16 плоских выводов
5962-01-324-7654, 5962013247654, MBM2732A-20 Данные.
 Получить расценки и купить 5962-01-324-7654, 5962013247654, MBM2732A-20 Данные. Получить расценки и купить
Получить расценки и купить 5962-01-324-7654, 5962013247654, MBM2732A-20 Данные. Получить расценки и купить5962-01-324-7654 (5962013247654) Информация NSN
| NSN | FSC | NIIN | Название позиции | INC |
|---|---|---|---|---|
| 5962-01-324-7654 | 013247654 | Микросхема, цифровая | 31779 |
5962-01-324-7654 Характеристики
| MRC | Параметр | Характеристики |
|---|---|---|
| AFGA | 19 Диапазон рабочих температур .0 / + 70,0 градус Цельсия||
| AGAV | Идентификация конечного элемента | Терморегистратор, alden electronic inc |
| CQSJ | Материал корпуса | Керамика |
| CQSZ | Конфигурация корпуса | Dual-in -line |
| CQWX | Выходная логическая форма | Металлооксидно-полупроводниковая логика N-типа |
5962-01-324-7654 Производственные номера деталей (SKU)
| MFG CAGE | СОСТОЯНИЕ | ISC | RNVC | RNCC | SADC | DAC | HCC | RNAAC | |
|---|---|---|---|---|---|---|---|---|---|
| MBM2732A-20 | 61271 | A | 9Z |
5962-01-324-7654 Производитель
5962-01- 324-7654 Идентификация FLIS
| PMIC | КОД ADPE | КОД CRITL | КОД DEMIL | DEMIL INTG | NIIN ASGMT | 59000 ESD|||||
|---|---|---|---|---|---|---|---|---|---|---|
| U | X | Q | 22-AUG-1990 | A | N | 85420 | 31779 |
5962-01-324-7654 Коды демилитаризации и управление
| DML | PMIC | HMIC | ADPEC | Критич. -7654 Поддержка нерасходуемых предметов -7654 Поддержка нерасходуемых предметов5962-01-324-7654 Грузовые перевозки
5962-01-324-3014, 5962013243014, LD8086-2 Данные.Получить расценки и купить5962-01-324-3014, 5962013243014, LD8086-2 Данные. Получить расценки и купить5962-01-324-3014 (5962013243014) Информация NSN
5962-01-324-3014 Характеристики
5962-01- 324-3014 Производственные номера деталей (SKU)
5962-01-324-3014 Производитель
5962-01-324-3014 Идентификация FLIS
5962-01-324-3014 Коды демилитаризации и управление
5962-01-324-3014 Опора для нерасходуемых предметов 904 895962-01-324-3014 Грузовые перевозки
границ | Модулирующие эффекты ингибирования устойчивой активности в корковой микросхеме моделиВведение Было показано, что нейроны префронтальной коры (ПФК) проявляют активность, которая часто сохраняется после окончания действия стимула, как было зарегистрировано in vivo во время периода задержки задач рабочей памяти (Goldman-Rakic, 1995). Интернейроны обладают большим разнообразием по своему распределению, связанности, нейрохимии, синаптическим связям и электрофизиологическим свойствам. FS обладают быстрым, высокочастотным и кратковременным потенциалом действия. Морфологически они были идентифицированы как нейроны люстр и корзин, и они экспрессируют кальций-связывающий белок, парвальбумин (PV). Они иннервируют сому и проксимальные дендритные компартменты пирамидных нейронов в ПФК (Wang et al., 2002; Зайцев, 2005). Интернейроны RS демонстрируют структуру возбуждения RS, в основном представляют собой двойной букет, и клетки типа Мартинотти экспрессируют белок кальбидин (CB) (Cauli et al., 1997). Было показано, что они иннервируют дистальные дендритные компартменты пирамидных нейронов в ПФК. Нейроны IS демонстрируют структуру возбуждения IS, являются в основном биполярными клетками и экспрессируют белок кальретинин (CR). Они проецируются на дендритные компартменты как PV-, так и CB-положительных клеток – предполагая, что по крайней мере некоторые типы CR-положительных клеток могут быть растормаживающими, – а также дистальные дендритные компартменты пирамидных нейронов в PFC (Cauli et al. Активность интернейронов, как было показано, вносит вклад в корковую динамику, сетевые колебания (Bartos et al., 2007), нейронная синхронизация пирамидных нейронов (Guidotti et al., 2005), сенсорная обработка (Börgers and Kopell, 2008), функция памяти (Jensen et al., 2007), целенаправленное поведение (Kvitsani et al., 2013 ) и социальное поведение (Yizhar et al., 2011). Было показано, что специфическая инактивация интернейронов PV приводит к снижению гамма-осцилляций в PFC (Sohal et al., 2009). Что касается функции рабочей памяти и постоянной активности, более ранние экспериментальные исследования показали роль опосредованного ГАМК A ингибирования в формировании полей памяти в ПФК (Rao et al., 1999). Более того, эксперименты in vitro предполагают, что активация ГАМК A предотвращает генерацию высокочастотных эпилептиформных всплесков, в то время как активация ГАМК B способствует прекращению состояний подъема и опускания, физиологического феномена, связанного с постоянной активностью (Mann et al. Однако остается много нерешенных вопросов относительно того, какие биофизические свойства или свойства связности различных типов интернейронов опосредуют индукцию постоянной активности, частотные характеристики возбуждения и синхронность нейронов. Стремясь дать некоторые ответы на эти вопросы, мы расширили недавно разработанную модель микросхемы PFC (Papoutsi et al., 2013), включив в нее три основных типа интернейронов: FS, регулярные пики (RS) и нерегулярные. -пиковые (IS) интернейроны.Мы использовали этот инструмент моделирования, чтобы проанализировать роль различных типов интернейронов в постоянной активности и определить, опосредуют ли профиль связности или физиологические свойства этих подтипов интернейронов их роли в постоянной активности. Наше моделирование показало, что (1) ток NMDA на интернейрон FS может модулировать индукцию постоянной активности, но не его поддерживающие свойства, (2) уменьшение ввода модели FS в модели пирамидных нейронов не позволяет использовать NMDA или GABA B -зависимая модуляция индукции постоянной активности и значительно увеличила частоту возбуждения, вариабельность ISI и нейронную синхронность во время постоянной активности, (3) изменения частоты возбуждения / вариабельности ISI не могли быть изменены добавлением какого-либо другого типа тормозящего воздействия на сома (опосредованная RS или IS) и (4) изменение синхронности может быть обращено вспять, но за пределами уровней контрольной сети путем добавления в сому ингибирующего входа, не относящегося к FS.В целом, наши данные предполагают, что соматическое торможение действует как стимулятор постоянной активности, при этом интернейрон FS модулирует выходной сигнал кардиостимулятора. Материалы и методы На основе известных электрофизиологических данных были построены четыре различных компартментных модельных клетки: один пирамидный нейрон и три различных интернейрона, модель FS, модель RS и модель IS. Модель пирамидального нейрона Используемая модель пирамидного нейрона была основана на модели, опубликованной в Papoutsi et al. (2013) и состоит из сомы, базального, проксимального и дистального дендритных отделов.Он включает моделирующие уравнения для 14 типов ионных механизмов, которые, как известно, присутствуют в этих нейронах, а также моделирующие уравнения для регуляции внутриклеточного кальция (то же уравнение, что и у Papoutsi et al., 2013). Пассивные и активные свойства модели пирамидного нейрона были подтверждены в соответствии с экспериментальными результатами Nasif et al. Таблица 1. Входные значения сопротивления модельных нейронов и полученные из электрофизиологических данных . Рис. 1. Проверка модели пирамидного нейрона. (A) Модель отклика на скачок тока в соме (0,17 нА). (B) Экспериментальный ответ пирамидного нейрона слоя V PFC на текущий шаговый импульс (адаптировано из Nasif et al., 2004). Интернейрон Модели Все три модели интернейронов включали ионные механизмы для быстрых токов Na + , A-типа K + и запаздывающего выпрямителя K + , а также моделирующие уравнения для регуляции механизма внутриклеточной буферизации кальция (те же уравнения как у Papoutsi et al. FS Interneuron Модель Модель интернейрона FS состояла из трех компартментов: соматического, дендритного и аксонного компартментов (дополнительная таблица 1). Соматический компартмент включал механизмы медленного тока K + (I Kslow ), высокопорогового активированного тока Ca ++ N-типа (N-тип) и катионного тока, активированного гиперполяризацией (Ih) ( Таблица 2), в дополнение к упомянутым выше.Емкость мембраны была установлена равной 1,2 мкФ / см2, а осевое сопротивление – 150 Ом / см (таблица 2). Мембранный потенциал покоя был доведен до -73 мВ, и его результирующее входное сопротивление составило 250 МОм (Kawaguchi and Kubota, 1993) (таблица 1). ПД этой модели нейрона FS имели короткую продолжительность и большую постгиперполяризацию. Таблица 2. Активные и пассивные ионные свойства интернейрона ФС модели . Рис. 2. Подтверждение электрофизиологических характеристик интернейрона FS (A), интернейрона RS (B) и модели интернейрона IS (C) . Реакция модели нейрона FS на увеличивающиеся инъекции деполяризующего тока (A1) по сравнению с экспериментальными данными из записей токового зажима ( A2 , адаптировано из Cauli et al., 1997), выявляют быстрорастущий профиль модели FS. Реакция модели нейрона RS на увеличивающиеся инъекции деполяризующего тока (B1) по сравнению с экспериментальными данными из записей токового зажима ( B2 , адаптировано из Cauli et al. RS Interneuron Модель Модель интернейрона RS состояла из трех отделов: соматического, дендритного и аксонного (дополнительная таблица 1) и включала механизмы для низкопорогового тока Ca ++ (T-тип) и Ih (таблица 3). . Мембранный потенциал был доведен до -64 мВ (Kawaguchi and Kubota, 1993). Емкость мембраны была установлена на 1,2 мкФ / см2, а осевое сопротивление – на 150 Ом / см (таблица 3). Результирующее входное сопротивление составляет 487 МОм (Таблица 1).Модельный нейрон ответил на деполяризующий импульс тока (0,05, 500 мс) 15 пиками с порогом потенциала действия -51 мВ (рис. 2В, внизу). Ток деполяризации 0,2 нА, 500 мс привел к отклику 60 Гц (рис. Таблица 3. Активные и пассивные ионные свойства интернейрона RS модели . IS Interneuron Модель Модель интернейрона IS состояла из четырех отсеков: соматического, двух дендритных и аксонального отсеков, имитирующих биполярную клетку (дополнительная таблица 1), и включала механизмы для медленного K + тока, быстрого Ca ++ -активированного K + тока и N -тип Ca ++ ток (таблица 4).Мембранный потенциал был доведен до -70 мВ (Kawaguchi and Kubota, 1993), емкость мембраны – 1,2 мкФ / см2, а осевое сопротивление – 150 Ом / см (таблица 4). Его входное сопротивление (~ 545 МОм), как показывают электрофизиологические данные (Зайцев, 2005) (Таблица 1). Типичный разряд этой ячейки в ответ на импульсы тока деполяризации состоял из излучения начального кластера из двух-шести AP, в зависимости от уровня деполяризации, за которым следовали AP, излучаемые с нерегулярной частотой (Cauli et al., 1997). Частота разряда увеличивается в зависимости от интенсивности стимуляции в соответствии с электрофизиологическими результатами (Cauli et al. Таблица 4. Активные и пассивные ионные свойства интернейрона IS модели . Модель микросхемы Мы построили микросхему из 20 моделей нейронов: 16 пирамидальных моделей, основанных на Papoutsi et al. (2013), 2 модели интернейронов FS, 1 модель интернейрона RS и 1 модель интернейрона IS, так что относительное количество интернейронов к нейронам пирамидальной модели составляло 20% (Dombrowski et al., 2001), а относительный ингибирующий вклад, исходящий от интернейронов FS, составлял 50% (Рисунок 4). Свойства связности, включая расположение и количество синаптических контактов, задержки между парами нейронов, а также электрофизиологические свойства их синаптических связей, были основаны на анатомических и электрофизиологических данных, аналогичных значениям, указанным в Papoutsi et al. (2013). В частности, модели пирамидных нейронов были полностью связаны рекуррентно (Wang et al., 2006) в их базальных дендритах с латентными периодами, взятыми из распределения Гаусса с μ = 1. Возможности подключения Аксон каждой модели пирамидного нейрона проецируется на базальный дендрит других моделей пирамидного нейрона. Модели пирамидных нейронов также проецируются на дендриты моделей FS, модели IS и модели RS. Однако специфика синаптической иннервации в неокортексе подразумевает, что рекуррентная сеть не организована случайным образом (Yoshimura and Callaway, 2005).Аксоны моделей интернейронов FS проецируются на сомы всех моделей пирамидных нейронов. Аксон модели интернейрона RS проецируется на дистальный апикальный дендрит всех моделей пирамидных нейронов (Murayama et al., 2009). Аксон модели интернейрона IS проецируется на сому модели интернейрона RS, обеспечивая тормозящий вход в микросхему, а также в дистальный апикальный дендрит всех моделей пирамидных нейронов. Кроме того, в моделях интернейронов ФС присутствуют ингибирующие аутапсы (Bacci et al. Таблица 5. Сводка синаптических связей в микросхеме . Количество синапсовОбщее количество возбуждающих синапсов для трех типов моделей интернейронов и тормозных синапсов на модели пирамидного нейрона было основано на анатомических данных (Tamás et al., 1997a, b; Markram et al., 2004). Общее количество тормозных синапсов на каждом пирамидальном модельном нейроне составляло 13% от общего числа возбуждающих синапсов (Peters et al., 2008). Сводная информация о количестве синапсов, введенных между каждым типом соединения, приведена в таблице 5. Проверка синаптических механизмов Проводимость возбуждающих и тормозных синаптических механизмов была скорректирована в соответствии с электрофизиологическими записями (Thomson and Deuchars, 1997; Angulo et al., 1999; Thomson and Destexhe, 1999; Xiang et al., 2002; Bacci et al., 2003; Woo). Рис. 3. Проверка синаптических свойств микросхемы.(A1) Текущие отклики от пирамидально-пирамидальной пары слоя 5. Когда мембранные потенциалы поддерживаются на уровне -70 мВ, токи преимущественно опосредуются рецепторами AMPA; тогда как при +60 мВ токи в значительной степени опосредуются рецепторами NMDA (адаптировано из Wang et al., 2008, Copyright 2008 National Academy of Sciences, США). (A2) Смоделированные отклики фиксации напряжения пирамидного модельного нейрона при -70 мВ (токи AMPA) и при +60 мВ (токи NMDA). (B1, B2) Экспериментальный (B1) и моделирование (B2) данных токов AMPA и NMDA в интернейронах FS ( B1 адаптировано из Wang and Gao, 2009). (C1, C2) Экспериментальный (C1) и данные моделирования (C2) токов AMPA и NMDA в интернейронах RS ( C1 адаптировано из Wang and Gao, 2009). Кроме того, были подтверждены токи, опосредованные рецептором GABA A (IPSC), между интернейроном FS и пирамидным нейроном, на основе Woo et al. (2007) и IPSC, опосредованный рецептором GABA B , был подтвержден в сравнении с экспериментальными данными Thomson et al. (1996) как у Papoutsi et al.(2013). По данным Xiang et al. (2002), амплитуда IPSC для пар FS-пирамид имела среднее значение значительно больше, чем для пар RS-пирамид. В частности, ток, опосредованный GABA A между парой RS-пирамидных нейронов, должен составлять 1/10 опосредованного GABA A тока между парой FS-пирамидных клеток (Xiang et al., 2002). Из-за отсутствия экспериментальных данных для IPSC пары IS-пирамидных нейронов, опосредованный GABA A ток этой пары оценивается как 1/10 от GABA A RS-пирамидной пары.Автоингибирование интернейронов PV намного сильнее, чем ингибирование между интернейронами разных типов, такими как пары IS-RS (Bacci et al., 2003). Аутаптические тормозные токи в интернейронах ФС вызывают относительно большой переходный ток амплитудой 0,35 мА (Bacci et al., 2003). Вышеупомянутый ток моделировался, как описано в Papoutsi et al. (2013). В различных экспериментах отношение NMDA-AMPA, равное 1,25, и соотношение GABAb-to-GABAa, равное 0,2, были взяты в качестве контрольного состояния. Фоновый шумКроме того, для наилучшего моделирования флуктуаций мембранного потенциала, наблюдаемых in vitro, из-за шума стохастического ионного канала (Linaro et al., 2011) искусственный ток с пуассоновскими характеристиками (средняя частота 0,02 Гц) вводился в сомы всех моделей нейронов. В частности, для модели нейрона IS амплитуда этого механизма была больше (средняя частота 0,035 Гц) (Golomb et al., 2007). Протокол стимуляцииПроксимальные апикальные дендриты моделей пирамидных нейронов стимулировали 120 возбуждающими синапсами (содержащими рецепторы AMPA и NMDA), которые активировались 10 раз при 20 Гц (желтые стрелки на рисунке 4) (Kuroda et al., 1998). Поскольку нейроны внутри микросхемы обладают схожими свойствами стимула (Yoshimura and Callaway, 2005; Petreanu et al., 2009), один и тот же начальный стимул был доставлен всем пирамидным нейронам. Рисунок 4. Схема микросхемы корректора коэффициента мощности . Микросхема PFC состоит из 16 моделей пирамидных нейронов (P) и 4 моделей интернейронов; 2 модели интернейронов с быстрым спайкованием (FS), 1 модель интернейрона с регулярными пиками (RS) и 1 модель интернейрона с нерегулярными пиками (IS).Все нейроны полностью связаны через повторяющиеся связи. Аксон каждой модели пирамидного нейрона проецируется на базальный дендрит других моделей пирамидного нейрона. Аксон модели интернейрона FS проецируется на сому всех моделей пирамидных нейронов. Аксоны моделей интернейронов RS и IS проецируются на дистальный апикальный дендрит всех моделей пирамидных нейронов. Аксон модели интернейрона IS проецируется на дендрит модели интернейрона RS. Пирамидный нейрон и модели интернейронов ФС также образуют аутаптические синапсы.Постоянная активность микросхемы индуцируется путем обеспечения внешнего синаптического моделирования для всех 16 пирамидных нейронов в их проксимальных апикальных дендритах. АнализАнализ данных выполнялся в Matlab (Mathworks, Inc). Интервалы между спайками (ISI) были рассчитаны для нейронного ответа каждой модели нейрона микросхемы во время стимула и во время постоянной активности. Среднее значение ISI каждого нейрона сети, а также коэффициент вариации в интервале времени 500 мс измеряли для каждого экспериментального состояния. Синхронизацию или десинхронизацию нейронов измеряли с помощью измерения расстояния SPIKE, которое чувствительно к совпадениям спайков (Kreuz et al., 2011). Для этого измерения мы получали последовательности спайков одновременно из популяции нейронов микросхемы, а затем рассчитывали интервалы времени между последовательными спайками, возникающими в любом из участвующих нейронов. Если между последовательностями импульсов нет фазовых задержек (нейроны срабатывают синхронно), индекс синхронизации будет иметь нулевые значения.В целом, небольшие значения индекса синхронизации указывают на синхронность, тогда как большие значения указывают на асинхронную пиковую активность (как у Papoutsi et al., 2013). В качестве дополнительной оценки синхронизации или десинхронизации между пиковыми нейронами в микросхеме во время каждого различного состояния мы измерили общее количество пиков, записанных во временных интервалах 1 мс, и построили график с дискретной частотой срабатывания. спектров мощности было создано на основе суммированных синаптических токов (AMPA, NMDA и GABA A ), генерируемых пирамидными нейронами в сети, усредненных для 10 испытаний, в течение 1-секундного периода постоянной постоянной активности, 3 с. после окончания раздражителя.Усредненные синаптические токи сначала были прорежены, а затем рассчитан среднеквадратичный спектр мощности с использованием метода периодограмм. Наличие моделиКод этой модели в среде моделирования NEURON будет доступен после контакта с соответствующим автором и будет размещен в базе данных ModelDB после публикации. РезультатыМы использовали модель микросхемы PFC с 20 нейронами, которая включала 16 биофизически детализированных моделей пирамидных клеток и 4 модели интернейронов: 2 FS, 1 RS и 1 IS интернейронная модель, чтобы изучить роль этих типов интернейронных клеток в устойчивых клетках. свойства возникновения и поддержания активности.Все смоделированные нейроны были проверены в сравнении с экспериментальными данными из внутриклеточных записей в срезах головного мозга (рис. 2 – подробности см. В разделе «Методы»). Кроме того, синаптические механизмы были проверены на основании экспериментальных данных (ток AMPA, отношение NMDA-AMPA, токи GABA) (рисунок 3 – подробности см. В разделе «Методы»). Постоянная активность в сети была вызвана внешним возбуждающим стимулом к апикальному дендриту (рис. 4). Подобно уменьшенной версии модели микросхемы (которая включала 7 нейронов пирамидальной модели и 2 интернейрона FS Papoutsi et al., 2013), постоянная индукция активности зависела от соотношения ГАМК B к ГАМК A и отношения NMDA к AMPA на моделях пирамидных нейронов (рис. 5A). Каждая модель нейрона имела различный образец возбуждения во время постоянной активности, в зависимости от ее собственных электрофизиологических характеристик (рис. 5В). Межспайковые интервалы (ISI) модели пирамидного нейрона во время постоянной активности составляли от 60 до 120 мс, то есть частота срабатывания 8-17 Гц (рис. 5C). Коэффициент вариации (CV) ISI, хотя и не очень высокий, как наблюдалось in vivo (Compte, 2006), был выше во время постоянной активности по сравнению с CV во время стимула (рис. 5D).Кроме того, мы обнаружили, что пиковая активность нейронов в сети была синхронизирована как во время реакции на стимул, так и во время постоянной активности, хотя синхронность во время стимула была больше по сравнению с таковой во время постоянной активности (рис. 5E). Эти свойства аналогичны соответствующим свойствам, наблюдаемым при постоянной активности во время задач с рабочей памятью (Constantinidis and Procyk, 2004). Рисунок 5. Индукция постоянной активности в сети.(A) Вероятность индукции стойкой активности (измеренная из 100 испытаний) зависела от соотношения ГАМКБ к ГАМКА и NMDA-АМРА на нейронах пирамидной модели, как было показано ранее (Papoutsi et al., 2013) . (B) Репрезентативные следы всех моделей нейронов во время стимула и во время постоянной активности. (C) График, показывающий ISI во время стимула и постоянной активности в ячейках 500 мс для всех моделей нейронов. (D) График, показывающий коэффициент вариации во время стимула и постоянной активности в ячейках 500 мс для всех моделей нейронов.ISI увеличивается во время начальной фазы постоянной активности для всех модельных нейронов по сравнению со стимулом. Это не относится к модели интернейронов FS. (E) График скорости срабатывания в дискретном времени, показывающий количество нейронов, которые срабатывают синхронно во время стимула и во время постоянной активности. (F) Изменение соотношения NMDA-AMPA на моделях пирамидных нейронов модулировало ISI, особенно во время начальных фаз постоянной активности. (G) Изменение соотношения NMDA-AMPA на моделях интернейронов FS модулирует вероятность индукции постоянной активности. (H) Изменение отношения NMDA-AMPA на моделях интернейронов FS не повлияло на ISI. Увеличение соотношения NMDA-AMPA на моделях пирамидных нейронов снизило ISI, особенно на начальных этапах постоянной активности (рис. 5F), что свидетельствует об увеличении частоты возбуждения. С другой стороны, модуляция соотношения NMDA-AMPA на моделях FS действительно модулирует% вероятности индукции постоянной активности (рис. 5G), но не ISI модели пирамидного нейрона (рис. 5H).Это согласуется с представлением о том, что регуляция рецепторов NMDA в интернейронах FS модулирует функцию PFC (Homayoun and Moghaddam, 2007), хотя медленная кинетика рецепторов NMDA может не допускать немедленного изменения срабатывания сети (Rotaru et al., 2011 ). Кроме того, изменение соотношения NMDA-AMPA либо на модели пирамидного нейрона, либо на модели FS не оказывает существенного влияния на синхронность между нейронами в микросхеме как во время стимула, так и во время постоянной активности (дополнительные таблицы 3, 4).Для остальной части исследования мы использовали следующие условия: GABA B -to-GABA A ratio = 0,2, NMDA-to-AMPA = 1,25 (модель пирамидного нейрона) и NMDA-to-AMPA = 0,5 (FS модель интернейрона). Чтобы изучить роль различных типов клеток интернейронов в постоянной активности, мы затем смоделировали сети «нокаута» для каждого подтипа интернейронов. Таким образом, мы сгенерировали микросхему PFC без моделей FS («FS KO») (рис. 6A1), микросхема без модели RS (сеть «RS KO») (рисунок 6A2) и микросхема без модели IS (сеть «IS KO») (рисунок 6A3).Мы обнаружили, что вероятность индукции стойкой активности всегда равна 1 для всех соотношений ГАМК B -к-ГАМК A и NMDA-AMPA в сети «FS KO» (рис. 6B1), хотя она не является значимой. изменено в сетевых моделях «RS KO» и «IS KO» (Рисунки 6B2,3). Таким образом, в сети «ФС КО» полностью теряется чувствительность к биофизической модуляции. Кроме того, ISI во время стимула и во время постоянной активности значительно уменьшаются в сети «FS KO» до 15 мс (т.е.е., частота близка к 80 Гц), но существенно не изменилась в сетях «RS KO» и «IR KO» (Рисунок 6C). Кроме того, CV ISI пирамидных нейронов во время стимула и во время постоянной активности значительно снижается в сети «FS KO» (рис. 6D). Это указывает на то, что скорость активации и ее вариабельность моделей пирамидных нейронов строго контролируется активностью моделей интернейронов FS, но не интернейронов RS и IR. Наконец, нейронная синхронность во время постоянной активности также значительно снижается в сети «FS KO», о чем свидетельствуют показатель индекса десинхронизации и график частоты активации в дискретном времени (Рисунки 6E, F).Это снова согласуется с другими исследованиями, предполагающими вклад пиков интернейронов FS на синхронизацию и колебания нейронов (Sohal et al., 2009). Рисунок 6. Свойства постоянной активности в различных моделях моделируемой сети «интернейрон KO» . (A) Графические изображения модели КО сети интернейрона ФС (А1) , сети КО интернейрона RS модели (А2) , сети КО интернейрона ИС модели (А3) . (B) Графики, показывающие постоянную индукцию активности по различным отношениям NMDA-к-AMPA и GABA B к ГАМК A в модели КО сети интернейронов FS модели (B1) , модели сети КО интернейронов RS (B2) , модель сети КО интернейрона IS (B3) . (C) График, показывающий ISI до и во время постоянного в ячейках 500 мс для управления и различных сетевых моделей «KO». (D) График, показывающий CV ISI модели пирамидного нейрона до и во время персистентности в ячейках 500 мс для контрольной и различных сетевых моделей «нокаут». (E) График, показывающий индекс десинхронности в управляющей и различных сетевых моделях «нокаут». (F) График скорости срабатывания в дискретном времени, показывающий синхронизацию между всеми моделями нейронов во время постоянной активности в сетевых моделях «контроль» (вверху) и «FS KO» (внизу). Поскольку сеть «FS KO» была единственной, показывающей существенные различия в отношении свойств постоянной активности, мы хотели продолжить изучение роли модели интернейронов FS. Таким образом, мы постепенно уменьшали количество ГАМКергических синапсов (как ГАМК A , так и ГАМК B ) из модели интернейрона FS на модель пирамидного нейрона, чтобы смоделировать менее серьезное и, возможно, более реалистичное нарушение в нейронной системе FS. функционирует.Мы обнаружили, что уменьшение входных данных модели FS в модель пирамидного нейрона увеличивает вероятность индукции постоянной активности по разным GABA B -to-GABA A (NMDA-to-AMPA = 1,25), тогда как при 40% или менее входов FS сохраняется, постоянная активность индуцируется во всех протестированных соотношениях GABA B -to-GABA A (фиг. 7A). Это говорит о том, что, когда теряется более 50% входов PV, микросхема PFC ведет себя так, как если бы модель FS не присутствовала, в отношении индукции постоянной активности.Это значительное увеличение индукции постоянной активности делает микросхему нечувствительной к модуляции ГАМК B . Рис. 7. Эффекты уменьшения количества ГАМКергических синаптических входов от моделей FS к моделям пирамидных нейронов. (A) Уменьшение количества синаптических входов от нейронов модели FS к нейронам пирамидной модели увеличивает диапазон соотношений GABA B -to-GABA A , в которых индуцируется постоянная активность.Когда в микросхеме присутствует менее 40% входных сигналов FS, постоянная активность индуцируется на 100% во всех соотношениях ГАМК B к ГАМК A . (B) По мере того, как количество входов FS уменьшается, ISI модели пирамидного нейрона уменьшается. (C) По мере того, как количество входов FS уменьшается, CV ISI модели пирамидного нейрона уменьшается. (D) Синхронность между всеми моделями нейронов во время постоянной активности значительно снижается, когда в микросхеме отсутствуют входы PV, в то время как она увеличивается за счет уменьшения количества синаптических входов от нейронов модели FS к нейронам пирамидальной модели. Кроме того, по мере того, как входные данные модели FS уменьшаются, ISI модели пирамидного нейрона во время постоянной активности постепенно уменьшаются, следовательно, частота возбуждения постепенно увеличивается (Рисунок 7B). Изменчивость ISI также уменьшается, когда входные данные FS уменьшаются до 60% или более, что делает этот индекс наиболее чувствительным к входным данным FS (рис. 7C). Наконец, индекс десинхронизации среди моделей нейронов в микросхеме постепенно уменьшается при уменьшении количества входов FS, но затем увеличивается, когда входы FS отсутствуют (т.е., сеть «ФС КО») (Рисунок 7D). Это говорит о том, что синхронность на самом деле увеличивается, когда процент входов FS в модели пирамидных нейронов блокируется, но затем уменьшается, когда входы отсутствуют. Многие роли нейронов FS в функциях корковой сети были приписаны его специфической связности, в частности проекции нейронов FS на сомы пирамидных нейронов (Lovett-Barron et al., 2012; Royer et al., 2012) . Однако по дизайну экспериментальные манипуляции не могут отличить целевое местоположение интернейрона от его физиологических характеристик.Итак, следующим шагом было детальное изучение роли этой специфической связи путем изменения места проекции модели нейрона FS на различные дендритные местоположения пирамидных нейронов, отличных от сомы; на базальных дендритах (сеть D0) (Рисунок 8A1), на проксимальных дендритах (сеть D1) (Рисунок 8A2), на дистальных дендритах (сеть D2) (Рисунок 8A3). Когда вход FS расположен где-то еще, кроме сомы, тогда вероятность индукции постоянной активности увеличивается до 100% (рисунок 8D), в то время как ISI и вариабельность ISI во время постоянной активности значительно снижаются (рисунки 8B, C).Кроме того, когда вход модели FS расположен в дендритных участках, а не в соме, индекс десинхронизации уменьшается, что свидетельствует об увеличении синхронности (рис. 8E). Следовательно, если модели нейронов FS не проецируются на сому, сетевая активность во время постоянной активности напоминает состояние, когда 50% меньших входов FS в сому являются активными (рис. 7). Рис. 8. Свойства постоянной активности, когда модель интернейрона FS проецируется на различные дендритные компартменты .Были сконструированы различные варианты микросхемы PFC, чтобы изучить влияние места проекции нейронов модели FS на пирамидные нейроны. (A) Графические изображения модели интернейрона FS, проецируемого на базальный (D0) (A1) , проксимальный (D1) (A2) и дистальный (D2) (A3) дендритный компартмент модель пирамидного нейрона. (B) График, показывающий ISI до и во время постоянного в ячейках 500 мс для управления и различных сетей проектируемых сайтов.ISI моделей пирамидных нейронов уменьшены во всех микросхемах, в которых нейроны модели FS проецируются на дендритные компартменты моделей пирамидных нейронов. (C) График, показывающий CV ISI до и во время сохранения в ячейках 500 мс для контрольных и различных сайтов проектирования. CV ISI моделей пирамидных нейронов снижены во всех микросхемах, в которых нейроны модели FS проецируются на дендритные компартменты моделей пирамидных нейронов. (D) График, показывающий индукцию постоянной активности в контрольной и различных сетях проектирующих сайтов FS. (E) График, показывающий индекс синхронности в сети контрольных и различных проектируемых сайтов. Соматическое торможение, обеспечиваемое интернейроном FS, по-видимому, необходимо для индукции правильной частоты возбуждения во время постоянной активности. Чтобы исключить возможность того, что тот же перисоматический ингибирующий эффект может быть достигнут другими типами интернейронов, мы изменили сеть, изменив проекцию и количество интернейронов FS на интернейрон RS (обратная сеть RS) (рис. 9A1) и с IS. интернейрон (обратная сеть IS) (рис. 9A2).Когда один интернейрон FS проецируется в дистальные дендриты пирамидных нейронов, но два интернейрона RS или два ISI обеспечивают соматическое ингибирование пирамидных нейронов, вероятность индукции постоянной активности увеличивается (рис. 9E), в то время как ISI и изменчивость ISI во время постоянной активности значительно уменьшаются (Рисунки 9B, C). Наконец, индекс десинхронизации уменьшается в обеих обратных сетях (рис. 9D). Вышеуказанные результаты оказались устойчивыми к изменениям кинетики возбуждающих синаптических механизмов моделей RS и IS и к изменениям значений проводимости тормозного синаптического механизма, которые модели RS и IS предоставляют своим связанным моделям нейронов (дополнительные рисунки 1, 2). ).Наши результаты также оказались устойчивыми к увеличению частоты (40 Гц) стимуляции, используемой для инициации постоянной активности (дополнительный рисунок 3). Эти результаты предполагают, что соматическое ингибирование, обеспечиваемое специфически FS-нейроном, необходимо для частоты возбуждения во время постоянной активности и позволяет модулировать индукцию постоянной активности. С другой стороны, синхронность микросхемы PFC может быть увеличена путем изменения либо проекции модели FS, либо изменения физиологического профиля моделей интернейронов, проецируемых на сому. Рис. 9. Различные варианты микросхемы PFC были сконструированы для того, чтобы изучить эффект пикового профиля модели нейрона, который обеспечивает соматическое ингибирование пирамидного нейрона . (A) Графические изображения микросхемы, в которой 2 модели интернейрона RS проецируются на сому, а модель интернейрона 1 FS проецируется на дистальный дендритный отсек моделей пирамидных нейронов (Reverse RS) (A1) и еще одного Микросхема, в которой 2 модели интернейрона IS проецируются на сому, а модель интернейрона 1 FS проецируется на дистальный дендритный отсек моделей пирамидных нейронов (Reverse IS) (A2) . (B) График, показывающий ISI до и во время постоянного в ячейках по 500 мс для управления и двух «обратных» состояний сети. ISI моделей пирамидных нейронов снижены во всех микросхемах, в которых нейрон модели FS проецируется на дистальные дендритные компартменты моделей пирамидных нейронов, в то время как любая из двух других моделей нейронов проецируется на сомы пирамидных нейронов. (C) График, показывающий CV ISI до и во время постоянного в ячейках 500 мс для управления и двух «обратных» состояний сети.CV ISI моделей пирамидных нейронов снижены во всех микросхемах, в которых нейрон модели FS проецируется на дистальные дендритные компартменты моделей пирамидных нейронов, в то время как любая из двух других моделей нейронов проецируется на сомы пирамидных нейронов. (D) График, показывающий индекс синхронности в управлении и два «обратных» состояния сети. (E) График, показывающий индукцию постоянной активности в контроле и двух «обратных» состояниях сети. В попытке сравнить наши результаты с доступной литературой относительно изменений в колебаниях сети при наличии дефектов ингибирования, мы проанализировали спектры мощности суммированных синаптических токов в различных модельных сетях, о которых сообщалось выше. В нашей сети управления мы наблюдаем наличие пика в спектре мощности на 20 Гц и меньшего пика на 40 Гц (рис. 10A, темно-синяя кривая). Оба пика отсутствуют в сети FS KO, что свидетельствует о значительной роли нейрона модели FS в поддержании этих колебаний.Кроме того, в сети RS KO уменьшается только пик 40 Гц, в то время как спектр мощности сети IS KO такой же, как у контрольной (рисунок 10A). Оба пика отсутствуют, когда остается 50% или менее входного сигнала FS в нейрон пирамидальной модели, однако пик на 40 Гц уже уменьшается, когда остается 80% входных сигналов FS (рис. 10B). Наконец, когда соматическое торможение обеспечивается нейроном RS или IS, пики на 20 и 40 Гц еще больше (рис. 10C). Таким образом, наши результаты показывают, что вход FS имеет решающее значение для поддержания колебаний сети, а также раскрывает новую роль модели нейрона RS в поддержании в первую очередь колебаний 40 Гц. Рис. 10. Спектры мощности суммарной синаптической активности в модельной сети. (A) Спектры мощности сетей управления, FS KO, RS KO и IS KO. (B) Спектры мощности сетей с разным процентом оставшихся входов FS на пирамидальных моделях. (C) Спектры мощности управления по сравнению с сетями обратного RS и обратного IS. ОбсуждениеВ этом исследовании мы специально определили отдельные и специфические роли интернейронов FS в свойствах постоянной активности.Во-первых, мы определили, что ввод тока NMDA в интернейроны только модулирует индукцию постоянной активности, но не ее пиковые свойства во время постоянной активности. Во-вторых, мы обнаружили, что входы FS-нейронов в пирамидные нейроны модулируют индукцию постоянной активности по принципу «все или ничего», в то время как свойства пиков во время постоянной активности градиентно. В-третьих, перемещение входов FS от сомы к другим дендритным компартментам имеет аналогичные эффекты для полного удаления нейронов FS из сети, указывая на значительную роль проектирующего сайта нейрона FS.Наконец, мы показываем, что замена соматического торможения моделями нейронов RS или IS не отменяет изменения индукции или частоты возбуждения, но изменяет синхронизацию нейронов (увеличивается за пределами контрольного состояния). NMDA-рецепторы в пирамидальном нейроне против интернейроновРоль токов NMDA пирамидных нейронов в постоянной активности и задачах рабочей памяти хорошо известна (Wang, 2001). Результаты нашего моделирования подтверждают значение токов NMDA пирамидных нейронов в индукции постоянной активности и модуляции как индукции, так и свойств возбуждения во время постоянной активности.Однако роли NMDA-рецепторов в интернейронах уделялось гораздо меньше внимания, особенно при модельных исследованиях постоянной активности. Результаты нашего моделирования показывают, что увеличение токов NMDA на интернейроны FS снижает вероятность индукции постоянной активности, в то время как уменьшение токов NMDA на интернейроны FS увеличивает вероятность индукции постоянной активности (рис. 5G). Эта двунаправленная модуляция предполагает, что рецепторы NMDA на интернейронах FS играют критическую роль в индукции постоянной активности и, следовательно, в производительности рабочей памяти.Наши результаты частично согласуются с более общей кортикальной моделью (Spencer, 2009), показывающей влияние NMDA на интернейроны FS на сетевую активность и синхронизацию. Кроме того, поскольку NMDA на интернейронах FS модулирует индукцию постоянной активности в нашей модели, мы прогнозируем, что это может ухудшить рабочую память и другие функции PFC. В самом деле, удаление функциональных NMDA рецепторов из интернейронов FS, как было показано, приводит к таким поведенческим дефектам (Belforte et al., 2009). Роль интернейронов в синхронности нейронов и гамма-частотных колебанияхБыло высказано предположение, что корковые колебания, особенно гамма-частоты, вносят значительный вклад в некоторые когнитивные функции, такие как избирательное внимание, восприятие.Считается, что эти колебания отражают синхронную активность ритмично запускаемых нейронов (Jensen et al., 2007). Активность интернейронов PV / FS, как было обнаружено в нескольких исследованиях, как экспериментальных, так и вычислительных, играет значительную роль в поддержании вышеуказанных колебаний и синхронизации нейронов (Borgers et al., 2008; Cardin et al., 2009; Sohal et al. , 2009; Vierling-Claassen et al., 2010). Гамма-колебания, как было показано, увеличивают взаимную информацию между входящей синаптической частотой и выходом потенциалов действия (Sohal et al., 2009). В нашей модели существует двунаправленная модуляция нейрональной синхронности интернейроном FS. Уменьшение входного сигнала FS приводит к увеличению синхронности, в то время как смоделированное состояние «KO» приводит к снижению синхронности. Замена входа FS на нейроны RS или IR увеличила синхронность, но вышла за уровни управляющей сети. Это говорит о том, что влияние ввода FS на синхронность нейронов сложное. Небольшое сокращение входных сигналов FS (20%) приводит как к увеличению синхронности, так и к небольшому отклонению в скорости срабатывания и изменчивости ISI, предполагая, что это может быть полезно для сетевой активности и, возможно, может привести к улучшению рабочей памяти.Однако снижение на 40% или больше синаптических входов, опосредованных FS, приводит не только к увеличению синхронности, но также к увеличению частоты возбуждения и снижению вариабельности ISI, указывая на возможный дефект, который может двигать сетевую активность в сторону эпилептиформного поведения. Следовательно, как упоминалось в Yu et al. (2004), «это не более слабое или сильное, но соответствующее синхронное состояние может иметь большее функциональное значение при сенсорном кодировании». Изменения интернейронов и болезниСовпадающие экспериментальные и клинические данные свидетельствуют о том, что дисфункция ГАМКергической системы и связанный с этим дисбаланс между возбуждением и торможением в коре головного мозга лежит в основе, по крайней мере, части патофизиологии нескольких нейропсихиатрических расстройств, таких как шизофрения, эпилепсия и аутизм (Marín, 2012). В частности, дефекты интернейронов очень сильно связаны с шизофренией (Lewis et al., 2005). Было показано, что у больных шизофренией снижены уровни синтезирующего ГАМК фермента, GAD67 и PV (Akbarian, 1995; Volk et al., 2001). Кроме того, GAD67 и PV также снижены в нескольких моделях шизофрении на животных (Braun et al., 2007; Lodge et al., 2009). Кроме того, при шизофрении были обнаружены как снижение нагрузки на рабочую память, так и снижение мощности гамма-осцилляторной активности в дорсолатеральном ПФК (Gonzalez-Burgos and Lewis, 2008), что также наблюдается при инактивации интернейронов ЛВ светом (Sohal et al., 2009). Снижение маркеров ингибирующей передачи, таких как количество дифференцированных интернейронов PV и CB, обнаруживается в моделях аутизма на животных (Eagleson et al., 2010; Fu et al., 2012), в то время как снижение GAD67 (маркер ингибирующей передачи) также снижается у людей с аутизмом (Fatemi et al., 2002). Кроме того, снижение гамма-ответа в затылочной коре также было обнаружено в исследовании на людях (Wright et al., 2012). Эпилепсия – еще одно заболевание, связанное с уменьшением популяции интернейронов, о чем в основном свидетельствует появление эпилептического поведения на животных моделях с уменьшенным количеством интернейронов (Cobos et al., 2005; Батт и др., 2008; Гант и др., 2009; Peñagarikano et al., 2011). Более того, эпилепсия характеризуется чрезмерной синхронностью нейронов (Traub and Wong, 1982), предполагая, что снижение функции интернейронов также может быть связано с повышенной синхронностью. Наши результаты предсказывают, что уменьшение более чем на 50% количества интернейронов или ГАМКергических синапсов приводит к нарушению индукции устойчивой активности, специфичной для стимула. Следовательно, любой тип стимула, независимо от нейромодуляции, может привести к постоянной активности, состоянию, которое должно значительно ухудшить выполнение задач с рабочей памятью и других когнитивных функций, зависимых от PFC, таких как внимание и поведенческая гибкость. Связь и физиологические свойстваВсе три различных подтипа интернейронов различаются по своим физиологическим характеристикам, а также по расположению синаптических мишеней. В частности, для интернейрона FS он имеет FS, высокочастотный физиологический профиль и в первую очередь нацелен на соматическую область пирамидных нейронов (Markram et al., 2004). Было высказано предположение, что наблюдаемая функция конкретных подтипов интернейронов в основном связана с расположением их синаптических мишеней (Wang et al., 2004). Значение проекционного сайта в пирамидных нейронах также было показано в нашем исследовании, поскольку различия в индукции постоянной активности, частоте возбуждения и вариабельности ISI видны, когда интернейрон FS проецируется в любой дендритный компартмент, а не в сому. Однако наши симуляции также показывают, что физиологический профиль FS также необходим, поскольку замена модели нейрона FS на модели нейронов RS или IR в соме не отменяет изменения индукции, частоты возбуждения и вариабельности ISI.Вместо этого изменения синхронности обращаются, но не до базовых уровней. В частности, когда соматическое торможение обеспечивается нейронами RS или IS, синхронность и сетевые колебания становятся сильнее. Это говорит о том, что соматическое торможение, обеспечиваемое интернейронами с активностью FS, абсолютно необходимо для различных свойств постоянной активности. Однако любой тип ингибирования, опосредованный моделями нейронов RS или IR, может поддерживать и даже дополнительно увеличивать синхронность нейронов.Таким образом, общее соматическое торможение может служить в качестве водителя ритма постоянной активности, но соматическое торможение, опосредованное ФС, необходимо для правильного проявления свойств устойчивой активности. Ограничения моделиНаша модельная микросхема включает в себя разные типы модельных нейронов (пирамидный нейрон и 3 разных типа интернейронов). Как видно из методов, модельная сеть, используемая в этой работе, сильно ограничена доступными экспериментальными данными. Однако источники неточности могут быть внесены из-за вариабельности препаратов, используемых для получения экспериментальных данных, а также из-за ограниченной доступности данных в отношении конкретной области мозга нашего исследования (префронтальная кора) и определенного слоя (слой V). .По возможности данные, используемые для проверки моделей, были взяты из исследований пирамидных нейронов слоя V или различных типов интернейронов в префронтальной коре (Зайцев, 2005; Peters et al., 2008; Wang et al., 2008; Wang and Гао, 2009). Однако в случаях, когда не было доступных данных из определенной области и определенного слоя, данные из неспецифической лобной коры (Kawaguchi and Kubota, 1993; Lubke et al., 1996; Thomson and Lamy, 2007; Woo et al., 2007) ) или специфические первичные сенсорные области либо для пирамидных нейронов, либо для интернейронов (Cauli et al., 1997; Tamás et al., 1997a, b; Xiang et al., 2002; Toledo-Rodriguez et al., 2005) и были использованы. Еще одна проблема, связанная с доступными данными, используемыми для ограничения модели, – это возраст животных, использованных в экспериментальных исследованиях. Большинство приведенных выше исследований, использованных для проверки наших моделей, проводилось на грызунах очень молодого возраста (до месяца), хотя некоторые из них были, например, на взрослых животных (Wang and Gao, 2009). Поскольку становится очевидным, что возраст играет очень важную роль в клеточной физиологии и лежащих в основе клеточных механизмах (McCutcheon and Marinelli, 2009), наши выводы ограничиваются использованием конкретных доступных данных.Если станут доступны более конкретные данные из префронтальной коры, предпочтительно от взрослых, конкретные или будущие модели могут быть ограничены более строгим образом. Кроме того, имеющиеся данные также изменчивы в отношении конкретного исследованного слоя. Следовательно, исследования с использованием приматов предполагают, что рекуррентные сети в основном на уровне III и в меньшей степени на уровне V PFC опосредуют постоянную активность, наблюдаемую во время задач рабочей памяти (Wang et al., 2013). Несколько моделей рабочей памяти в литературе (Compte et al., 2000; Wang et al., 2004) моделируют рекуррентные сети уровня III (Kritzer and Goldman-Rakic, 1995) в большем масштабе по сравнению с моделью, представленной здесь. Однако основные результаты, касающиеся вклада, например, токов NMDA, аналогичны. Даже модели слоя III ограничивают свои биофизические параметры, используя электрофизиологические записи слоя V (Seamans et al., 1997) или записи, не ограниченные конкретным слоем (Connors and Gutnick, 1990; Hammond and Crepel, 1992). В свете недавних данных о различиях в биофизических свойствах пирамидных нейронов слоя III и V (Bremaud et al., 2007; de Kock, 2013), будущие исследования по моделированию могут быть еще более ограничены записями, специфичными для слоя, возможно, раскрывая специфичное для слоя кодирование информации или свойства постоянной активности. Прогнозы моделей и будущее использованиеНаша модель генерирует два важных прогноза, которые можно проверить экспериментально. Во-первых, он предсказывает, что модуляция тока NMDA на интернейронах модулирует только индукцию постоянной активности, а не возбудимость или синхронность нейронов во время постоянной активности.Таким образом, блокада рецепторов NMDA, в частности, в интернейронах, может увеличивать возникновение состояний вверх и вниз в экспериментах in vitro или увеличивать постоянную активность неселективных стимулов в задачах in vivo . Во-вторых, он предсказывает, что постепенное снижение процента ФС-опосредованных ГАМКергических синапсов значительно увеличит скорость активации во время постоянной активности и уменьшит вариабельность ISI. Например, в моделях животных с пониженным содержанием ГАМКергических нейронов (Peñagarikano et al., 2011; Vidaki et al., 2012), можно было бы ожидать очень значительного увеличения возникновения состояний вверх и вниз, во время которых частота активации нейронов будет увеличиваться, а вариабельность ISI уменьшаться. Кроме того, из-за уровня биофизической детализации и обширных проверок, модельная сеть может использоваться в качестве инструмента для дальнейшего определения роли интернейронов в постоянной активности и активности, зависимой от стимулов. Некоторые примеры исследований, которые могут использовать и / или расширять модель, включают, помимо прочего, (а) определение роли двух других типов интернейронов (модели RS и IS) при различных протоколах стимуляции, (б) изучение роли специфических биофизических механизмов на интернейронах при постоянной активности или активности, зависимой от стимула, и (c) расширение модельной сети для включения правил пластичности, специфичных для различных типов интернейронов. Заявление о конфликте интересовАвторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов. БлагодарностиЭта работа была поддержана грантом Мари-Кюри IOF (FEAR MEMORY TRACE-253388) и наградой NARSAD для молодых исследователей Кириаки Сидиропулу и стартовым грантом ERC Панайоте Пойрази («dEMORY», ERC-2012-StG-311435). Дополнительные материалыДополнительные материалы к этой статье можно найти в Интернете по адресу: http: // www.frontiersin.org/journal/10.3389/fncir.2014.00007/abstract Список литературыАкбарян С. (1995). Экспрессия гена декарбоксилазы глутаминовой кислоты снижается без потери нейронов в префронтальной коре головного мозга шизофреников. Arch. Gen. Psychiatry 52, 258–266. DOI: 10.1001 / archpsyc.1995.03950160008002 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Ангуло М.С., Россье Дж. И Аудинат Э. (1999). Постсинаптические рецепторы глутамата и интегративные свойства быстрых интернейронов в неокортексе крыс. J. Neurophysiol . 82, 1295–1302. Pubmed Реферат | Pubmed Полный текст Баччи А., Хугенард Дж. Р. и Принс Д. А. (2003). Функциональная аутаптическая нейротрансмиссия в быстрых интернейронах: новая форма подавления обратной связи в неокортексе. Дж. Neurosci . 23, 859–866. Pubmed Реферат | Pubmed Полный текст Белфорте, Дж. Э., Зсирос, В., Склар, Э. Р., Цзян, З., Ю, Г., Ли, Ю. и др. (2009). Постнатальная абляция рецепторов NMDA в кортиколимбических интернейронах придает фенотипы, подобные шизофрении. Nat. Neurosci . 13, 76–83. DOI: 10.1038 / nn.2447 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Боргерс К., Эпштейн С. и Копелл Н. Дж. (2008). Гамма-колебания опосредуют конкуренцию стимулов и выбор внимания в модели корковой сети. Proc. Natl. Акад. Sci. США . 105, 18023–18028. DOI: 10.1073 / pnas.0809511105 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Браун, И., Гений, Дж., Грюнзе, Х., Бендер, А., Моллер, Х.-Дж., и Ружеску, Д. (2007). Изменения гиппокампа и префронтальных ГАМКергических интернейронов на животной модели психоза, вызванного антагонизмом к рецепторам NMDA. Schizophr. Res . 97, 254–263. DOI: 10.1016 / j.schres.2007.05.005 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Бремо А., Уэст Д. К. и Томсон А. М. (2007). Биномиальные параметры различаются между слоями неокортекса и разными классами связей в неокортексе взрослых крыс и кошек. Proc. Natl. Акад. Sci. США . 104, 14134–14139. DOI: 10.1073 / pnas.0705661104 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Батт, С. Дж. Б., Соуза, В. Х., Фуччилло, М. В., Хьерлинг-Леффлер, Дж., Миёси, Г., Кимура, С. и др. (2008). Требование Nkx2-1 во временной спецификации подтипов корковых интернейронов. Нейрон 59, 722–732. DOI: 10.1016 / j.neuron.2008.07.031 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Кардин, Дж.A., Carlen, M., Meletis, K., Knoblich, U., Zhang, F., Deisseroth, K., et al. (2009). Управление быстропротекающими клетками вызывает гамма-ритм и контролирует сенсорные реакции. Природа 459, 663–667. DOI: 10.1038 / nature08002 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Cauli, B., Audinat, E., Lambolez, B., Angulo, M.C., Ropert, N., Tsuzuki, K., et al. (1997). Молекулярное и физиологическое разнообразие корковых непирамидных клеток. Дж. Neurosci .17, 3894–3906. Pubmed Реферат | Pubmed Полный текст Cobos, I., Calcagnotto, M. E., Vilaythong, A. J., Thwin, M. T., Noebels, J. L., Baraban, S. C., et al. (2005). Мыши, лишенные Dlx1, демонстрируют специфическую для подтипа потерю интернейронов, снижение ингибирования и эпилепсию. Nat. Neurosci . 8, 1059–1068. DOI: 10.1038 / nn1499 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Compte, A. (2006). Вычислительные и in vitro исследования постоянной активности: переход к клеточным и синаптическим механизмам рабочей памяти. Neuroscience 139, 135–151. DOI: 10.1016 / j.neuroscience.2005.06.011 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Компте, А., Брунель, Н., Гольдман-Ракич, П. С. и Ван, X. J. (2000). Синаптические механизмы и сетевая динамика, лежащие в основе пространственной рабочей памяти в модели корковой сети. Cereb. Cortex 10, 910–923. DOI: 10.1093 / cercor / 10.9.910 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Домбровский, С.М., Хилгетаг, К.С., и Барбас, Х. (2001). Количественная архитектура отличает префронтальные корковые системы макаки-резуса. Cereb. Cortex 11, 975–988. DOI: 10.1093 / cercor / 11.10.975 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Иглсон, К. Л., Гравиелле, М. К., Шулютер МакФадьен-Кетчум, Л. Дж., Рассек, С. Дж., Фарб, Д. Х., и Левитт, П. (2010). Генетическое нарушение гена риска расстройств аутистического спектра PLAUR вызывает изменения субъединиц рецептора ГАМК. Неврология 168, 797–810. DOI: 10.1016 / j.neuroscience.2010.03.066 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Фатеми, С. Х., Халт, А. Р., Стэри, Дж. М., Канодиа, Р., Шульц, С. К., и Реалмуто, Г. Р. (2002). Белки декарбоксилазы глутаминовой кислоты 65 и 67 кДа снижены в париетальной и мозжечковой коре головного мозга аутистов. Biol. Психиатрия 52, 805–810. DOI: 10.1016 / S0006-3223 (02) 01430-0 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Fu, C., Cawthon, B., Clinkscales, W., Bruce, A., Winzenburger, P., and Ess, K.C. (2012). Развитие и функция ГАМКергических интернейронов модулируются геном Tsc1. Cereb. Cortex 22, 2111–2119. DOI: 10.1093 / cercor / bhr300 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Gant, J. C., Thibault, O., Blalock, E. M., Yang, J., Bachstetter, A., Kotick, J., et al. (2009). Уменьшение количества интернейронов и увеличение припадков у мышей с дефицитом нейропилина 2: последствия для аутизма и эпилепсии. Эпилепсия 50, 629–645. DOI: 10.1111 / j.1528-1167.2008.01725.x Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Голомб Д., Доннер К., Шахам Л., Шлосберг Д., Амитаи Ю. и Хансель Д. (2007). Механизмы возбуждающих паттернов в корковых интернейронах с быстрым спайком. PLoS Comput. Биол . 3: e156. DOI: 10.1371 / journal.pcbi.0030156 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Гонсалес-Бургос, Г., и Льюис, Д.А. (2008). ГАМК-нейроны и механизмы сетевых колебаний: значение для понимания корковой дисфункции при шизофрении. Schizophr. Бык . 34, 944–961. DOI: 10.1093 / schbul / sbn070 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Guidotti, A., Auta, J., Davis, J. M., Dong, E., Grayson, D. R., Veldic, M., et al. (2005). ГАМКергическая дисфункция при шизофрении: новые стратегии лечения на горизонте. Психофармакология 180, 191–205.DOI: 10.1007 / s00213-005-2212-8 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Homayoun, H., and Moghaddam, B. (2007). Гипофункция рецептора NMDA оказывает противоположное действие на интернейроны префронтальной коры и пирамидные нейроны. Дж. Neurosci . 27, 11496–11500. DOI: 10.1523 / JNEUROSCI.2213-07.2007 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Кавагути Ю. и Кубота Ю. (1993). Корреляция физиологических подгрупп непирамидных клеток с парвальбумин- и кальбиндин (D28k) -иммунореактивными нейронами в слое v лобной коры головного мозга крысы. J. Neurophysiol . 70, 387–396. Pubmed Реферат | Pubmed Полный текст Кройц, Т., Чичарро, Д., Грешнер, М., и Анджеяк, Р. Г. (2011). Временные и масштабные адаптивные меры синхронности спайковых поездов. J. Neurosci. Методы 195, 92–106. DOI: 10.1016 / j.jneumeth.2010.11.020 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Критцер, М. Ф., и Гольдман-Ракич, П. С. (1995). Организация внутренних цепей основных слоев и подслоев дорсолатеральной префронтальной коры у макаки-резуса. J. Comp. Neurol . 359, 131–143. DOI: 10.1002 / cne.9035 | Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Курода М., Йокофудзита Дж. И Мураками К. (1998). Ультраструктурное исследование нервной цепи между префронтальной корой и медиодорсальным ядром таламуса. Prog. Нейробиол . 54, 417–458. DOI: 10.1016 / S0301-0082 (97) 00070-1 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Квицани, Д., Ранаде, С., Хангья, Б., Танигучи, Х., Хуанг, Дж. З., и Кепек, А. (2013). Четкие поведенческие и сетевые корреляты двух типов интернейронов в префронтальной коре. Природа 498, 363–369. DOI: 10.1038 / nature12176 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Линаро Д., Стораче М. и Джульяно М. (2011). Точное и быстрое моделирование канального шума в модельных нейронах на основе проводимости с помощью диффузионного приближения. PLoS Comput. Биол .7: e1001102. DOI: 10.1371 / journal.pcbi.1001102 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Лодж, Д. Дж., Беренс, М. М., и Грейс, А. А. (2009). Потеря парвальбуминсодержащих интернейронов связана со снижением осцилляторной активности на животной модели шизофрении. Дж. Neurosci . 29, 2344–2354. DOI: 10.1523 / JNEUROSCI.5419-08.2009 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Ловетт-Бэррон, М., Turi, G.F., Kaifosh, P., Lee, P.H., Bolze, F., Sun, X.-H., et al. (2012). Регулирование нейрональных входных преобразований с помощью настраиваемого дендритного торможения. Nat. Neurosci . 15, 423–430. DOI: 10.1038 / nn.3024 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Lubke, J., Markram, H., Frotscher, M., and Sakmann, B. (1996). Частота и дендритное распределение аутапсов, установленных пирамидными нейронами 5 слоя в развивающемся неокортексе крысы: сравнение с синаптической иннервацией соседних нейронов того же класса. Дж. Neurosci . 16, 3209–3218. Pubmed Реферат | Pubmed Полный текст Манн, Э. О., Коль, М. М., и Паульсен, О. (2009). Различная роль рецепторов GABAA и GABAB в балансировании и прекращении постоянной корковой активности. Дж. Neurosci . 29, 7513–7518. DOI: 10.1523 / JNEUROSCI.6162-08.2009 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Маркрам, Х., Толедо-Родригес, М., Ван, Ю., Гупта, А., Зильберберг, Г., и Ву, К.(2004). Интернейроны тормозной системы неокортекса. Nat. Преподобный Neurosci . 5, 793–807. DOI: 10.1038 / nrn1519 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Мураяма, М., Перес-Гарси, Э., Невиан, Т., Бок, Т., Сенн, В., и Ларкум, М. Э. (2009). Дендритное кодирование сенсорных стимулов, контролируемых глубокими корковыми интернейронами. Природа 457, 1137–1141. DOI: 10.1038 / nature07663 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Насиф, Ф.Дж., Сидиропулу К. и Уайт Ф. Дж. (2004). Повторное введение кокаина увеличивает возбудимость мембран пирамидных нейронов медиальной префронтальной коры головного мозга крыс. J. Pharmacol. Exp. Ther . 312, 1305–1313. DOI: 10.1124 / jpet.104.075184 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Невиан Т., Ларкум М. Э., Польски А. и Шиллер Дж. (2007). Свойства базальных дендритов пирамидных нейронов слоя 5: прямое исследование записи патч-кламп. Nat. Neurosci . 10, 206–214. DOI: 10.1038 / nn1826 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Папуци, А., Сидиропулу, К., Куцуридис, В., и Пойрази, П. (2013). Индукция и модуляция постоянной активности в модели микросхемы V PFC. Фронт. Нейронные цепи . 7: 161. DOI: 10.3389 / fncir.2013.00161. Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Пеньягарикано, О., Абрахамс, Б.С., Herman, E. I., Winden, K. D., Gdalyahu, A., Dong, H., et al. (2011). Отсутствие CNTNAP2 приводит к эпилепсии, аномалиям миграции нейронов и основным дефицитам, связанным с аутизмом. Cell 147, 235–246. DOI: 10.1016 / j.cell.2011.08.040 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Рао, С. Г., Уильямс, Г. В., и Гольдман-Ракич, П. С. (1999). Разрушение и создание пространственной настройки растормаживанием: ГАМК. Дж. Neurosci . 20, 485–494. Pubmed Реферат | Pubmed Полный текст Ротару, Д.К., Йошино, Х., Льюис, Д. А., Эрментроут, Г. Б., и Гонсалес-Бургос, Г. (2011). Подтипы глутаматных рецепторов, опосредующие синаптическую активацию нейронов префронтальной коры: актуальность для шизофрении. Дж. Neurosci . 31, 142–156. DOI: 10.1523 / JNEUROSCI.1970-10.2011 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Ройер, С., Земельман, Б.В., Лошонци, А., Ким, Дж., Чанс, Ф., Маги, Дж. К. и др. (2012). Контроль времени, скорости и всплесков клеток места гиппокампа с помощью дендритного и соматического торможения. Nat. Neurosci . 15, 769–775. DOI: 10.1038 / nn.3077 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Симанс, Дж. К., Горелова, Н., Янг, К. Р. (1997). Вклад потенциалзависимых Са-каналов в проксимальных и дистальных дендритах в синаптическую интеграцию в префронтальных кортикальных нейронах. Дж. Neurosci . 17, 5936–5948. Pubmed Реферат | Pubmed Полный текст Сидиропулу, К., Лу, Ф.-М., Фаулер, М.А., Сяо, Р., Филлипс К., Озкан Э. Д. и др. (2009). Дофамин модулирует опосредованную mGluR5 деполяризацию, лежащую в основе персистирующей префронтальной активности. Nat. Neurosci . 12, 190–199. DOI: 10.1038 / nn.2245 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Спенсер, К. (2009). Функциональные последствия аномалий коркового контура на гамма-колебания при шизофрении: выводы из компьютерного моделирования. Фронт. Гм. Neurosci . 3:33. DOI: 10.3389 / нейро.09.033.2009 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Tamás, G., Buhl, E.H., and Somogyi, P. (1997a). Быстрые IPSPs, вызываемые через несколько сайтов синаптического высвобождения различными типами ГАМКергических нейронов в зрительной коре головного мозга кошки. J. Physiol . 500 (Pt 3), 715–738. Pubmed Реферат | Pubmed Полный текст Tamás, G., Buhl, E.H., and Somogyi, P. (1997b). Массивная аутаптическая самоиннервация ГАМКергических нейронов зрительной коры головного мозга кошек. Дж. Neurosci . 17, 6352–6364. Pubmed Реферат | Pubmed Полный текст Томсон А. М., Уэст Д. К., Хан Дж. И Дюшарс Дж. (1996). IPSP с одним аксоном, индуцированные в пирамидных клетках тремя классами интернейронов в срезах неокортекса крысы. J. Physiol . 496, 81–102. Pubmed Реферат | Pubmed Полный текст Томсон А. М. и Дестекс А. (1999). Двойные внутриклеточные записи и вычислительные модели медленных тормозных постсинаптических потенциалов в неокортикальных и гиппокампальных срезах крыс. Neuroscience 92, 1193–1215. DOI: 10.1016 / S0306-4522 (99) 00021-4 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Томсон А. М. и Дюшарс Дж. (1997). Синаптические взаимодействия в неокортикальных локальных цепях: двойные внутриклеточные записи in vitro . Cereb. Cortex 7, 510–522. DOI: 10.1093 / cercor / 7.6.510 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Толедо-Родригес, М., Гудман, П., Иллич, М., Ву, К., и Маркрам, Х. (2005). Профили экспрессии генов нейропептидов и кальций-связывающих белков позволяют прогнозировать анатомический тип нейронов у молодых крыс. J. Physiol . 567, 401–413. DOI: 10.1113 / jphysiol.2005.089250 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Видаки М., Тиводар С., Доулгераки К., Тибулевич В., Кессарис Н., Пахнис В. и др. (2012). Rac1-зависимый выход из клеточного цикла предшественников MGE и миграция ГАМКергических интернейронов в кору. Cereb. Cortex 22, 680–692. DOI: 10.1093 / cercor / bhr145 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Фирлинг-Клаассен, Д., Кардин, Дж. А., Мур, К. И., и Джонс, С. Р. (2010). Вычислительное моделирование отдельных неокортикальных колебаний, вызванных избирательным оптогенетическим приводом клеточного типа: отдельные резонансные контуры, управляемые низкопороговыми импульсами и интернейронами с быстрым выбросом. Фронт. Гм. Neurosci . 4: 198. DOI: 10.3389 / fnhum.2010.00198 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Волк Д. В., Остин М. К., Пьерри Дж. Н., Сэмпсон А. Р. и Льюис Д. А. (2001). МРНК транспортера-1 ГАМК в префронтальной коре при шизофрении: снижение экспрессии в подмножестве нейронов. Am. J. Psychiatry 158, 256–265. DOI: 10.1176 / appi.ajp.158.2.256 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Wang, H.-X., and Gao, W.-J. (2009). Клеточно-специфическое развитие рецепторов NMDA в интернейронах префронтальной коры головного мозга крыс. Нейропсихофармакология 34, 2028–2040. DOI: 10.1038 / npp.2009.20 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Wang, H.-X., Stradtman, G.G., Wang, X.-J., and Gao, W.-J. (2008). Специализированная функция рецептора NMDA в повторяющейся микросхеме 5-го слоя префронтальной коры взрослой крысы. Proc. Natl. Акад. Sci. США . 105, 16791–16796. DOI: 10.1073 / pnas.0804318105 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Ван, М., Yang, Y., Wang, C.-J., Gamo, N.J., Jin, L.E., Mazer, J.A., et al. (2013). Рецепторы NMDA обеспечивают постоянное возбуждение нейронов во время рабочей памяти в дорсолатеральной префронтальной коре. Neuron 77, 736–749. DOI: 10.1016 / j.neuron.2012.12.032 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Ван, X.-J. (1999). Синаптическая основа постоянной активности коры: важность рецепторов NMDA для рабочей памяти. Дж. Neurosci . 19, 9587–9603. Pubmed Реферат | Pubmed Полный текст Ван Х. Дж., Тегнер Дж., Константинидис К. и Гольдман-Ракич П. С. (2004). Разделение труда между отдельными подтипами тормозных нейронов в корковой микросхеме рабочей памяти. Proc. Natl. Акад. Sci. США . 101, 1368–1373. DOI: 10.1073 / pnas.0305337101 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Ван, Ю., Гупта, А., Толедо-Родригес, М., Ву, К., и Маркрам, Х. (2002).Анатомические, физиологические, молекулярные и схемные свойства клеток гнездовой корзины в развивающейся соматосенсорной коре. Cereb. Cortex 12, 395–410. DOI: 10.1093 / cercor / 12.4.395 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Ван Й., Маркрам Х., Гудман П. Х., Бергер Т. К., Ма Дж. И Гольдман-Ракич П. С. (2006). Неоднородность пирамидальной сети медиальной префронтальной коры. Nat. Neurosci . 9, 534–542. DOI: 10.1038 / nn1670 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Ву, Р.-S., Li, X.-M., Tao, Y., Carpenter-Hyland, E., Huang, Y.Z., Weber, J., et al. (2007). Нейрегулин-1 усиливает вызванное деполяризацией высвобождение ГАМК. Neuron 54, 599–610. DOI: 10.1016 / j.neuron.2007.04.009 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Райт, Б., Олдерсон-Дей, Б., Прендергаст, Г., Беннет, С., Джордан, Дж., Уиттон, К. и др. (2012). Гамма-активация у молодых людей с расстройствами аутистического спектра и типично развивающимися элементами управления при просмотре эмоций на лицах. PLoS ONE 7: e41326. DOI: 10.1371 / journal.pone.0041326 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Сян З., Хугенард Дж. Р. и Принс Д. А. (2002). Синаптическое ингибирование пирамидных клеток, вызванное различными межнейрональными подтипами в слое V зрительной коры головного мозга крысы. J. Neurophysiol . 88, 740–750. DOI: 10.1152 / jn.00635.2001 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Ижар О., Фенно Л.E., Prigge, M., Schneider, F., Davidson, T.J., O’Shea, D.J., et al. (2011). Неокортикальный баланс возбуждения / торможения при обработке информации и социальная дисфункция. Природа 477, 171–178. DOI: 10.1038 / природа10360 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Йошида М., Хассельмо М. Э. (2009). Постоянное возбуждение поддерживается внутренним клеточным механизмом в компоненте системы направления головы. Дж. Neurosci . 29, 4945–4952.DOI: 10.1523 / JNEUROSCI.5154-08.2009 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст Зайцев А.В. (2005). Локализация кальцийсвязывающих белков в физиологически и морфологически охарактеризованных интернейронах дорсолатеральной префронтальной коры головного мозга обезьян. Cereb. Cortex 15, 1178–1186. DOI: 10.1093 / cercor / bhh318 Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст .Больше новостей |
|---|
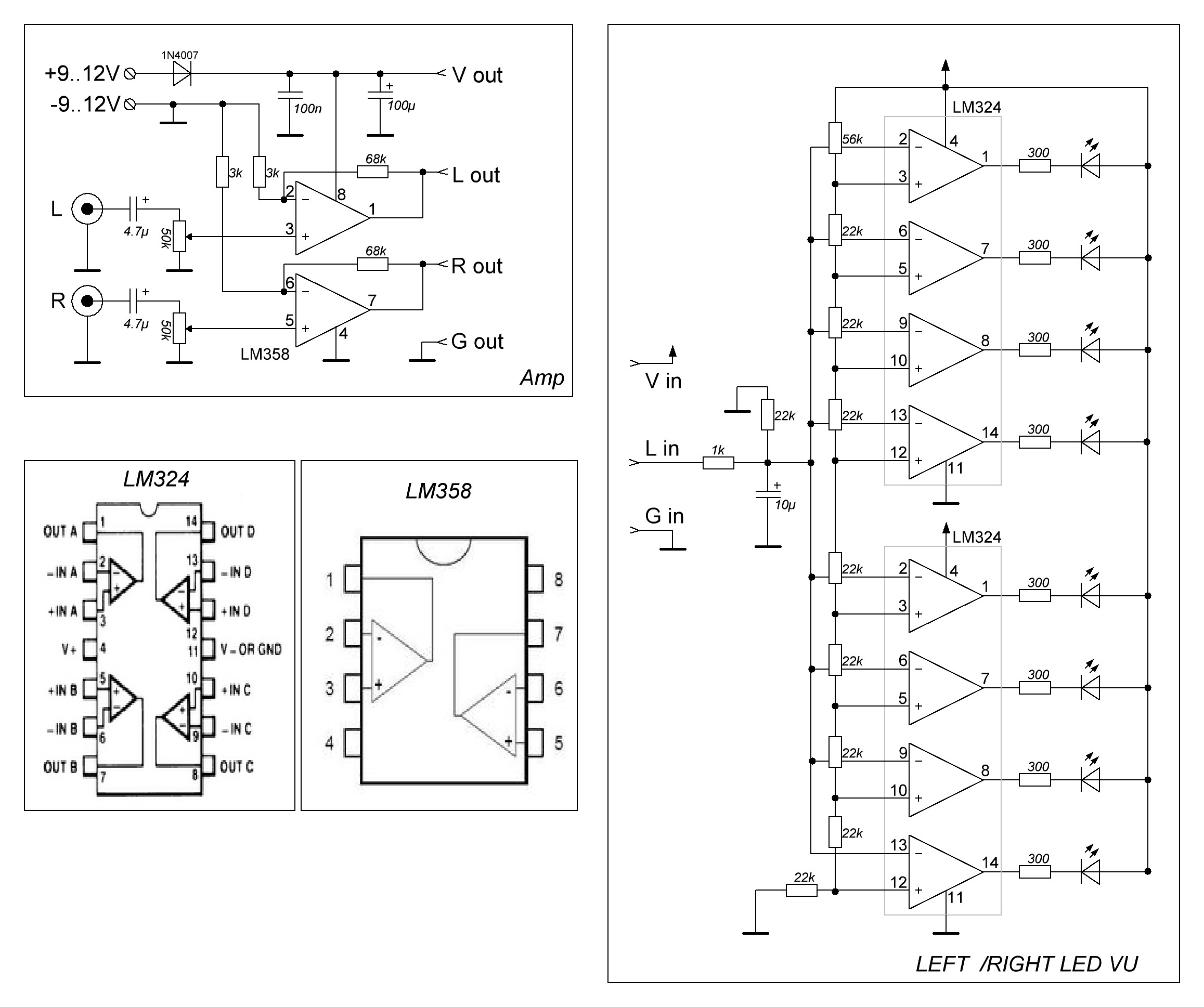
 Эта постоянная активность соответствует представлению памяти в режиме онлайн в течение короткого периода времени. Было показано, что его появление зависит от баланса возбуждения, обеспечиваемого глутаматергическими нейронами, и торможения, обеспечиваемого ГАМКергическими интернейронами (Goldman-Rakic, 1995; Compte, 2006), а также от динамики отдельных нейронов (Sidiropoulou et al., 2009). ; Йошида, Хассельмо, 2009). В частности, на основании компьютерных исследований было показано, что активация рецепторов глутамата NMDA играет очень важную роль в поддержании стабильной постоянной активности (Wang, 1999; Compte et al., 2000), in vitro экспериментов с срезами мозга (McCormick, 2003) и записей in vivo на обезьянах (Wang et al., 2013). Однако относительно немного исследований изучали, как структура и физиология интернейронов вносят вклад в физиологическую функцию префронтальной коры (PFC) (Rao et al., 1999; Wang et al., 2004).
Эта постоянная активность соответствует представлению памяти в режиме онлайн в течение короткого периода времени. Было показано, что его появление зависит от баланса возбуждения, обеспечиваемого глутаматергическими нейронами, и торможения, обеспечиваемого ГАМКергическими интернейронами (Goldman-Rakic, 1995; Compte, 2006), а также от динамики отдельных нейронов (Sidiropoulou et al., 2009). ; Йошида, Хассельмо, 2009). В частности, на основании компьютерных исследований было показано, что активация рецепторов глутамата NMDA играет очень важную роль в поддержании стабильной постоянной активности (Wang, 1999; Compte et al., 2000), in vitro экспериментов с срезами мозга (McCormick, 2003) и записей in vivo на обезьянах (Wang et al., 2013). Однако относительно немного исследований изучали, как структура и физиология интернейронов вносят вклад в физиологическую функцию префронтальной коры (PFC) (Rao et al., 1999; Wang et al., 2004). На основании их электрофизиологических характеристик были идентифицированы три основных класса, а именно: FS, интернейроны с регулярными пиками (RS) и нерегулярными пиками (IS) (Markram et al., 2004).
На основании их электрофизиологических характеристик были идентифицированы три основных класса, а именно: FS, интернейроны с регулярными пиками (RS) и нерегулярными пиками (IS) (Markram et al., 2004).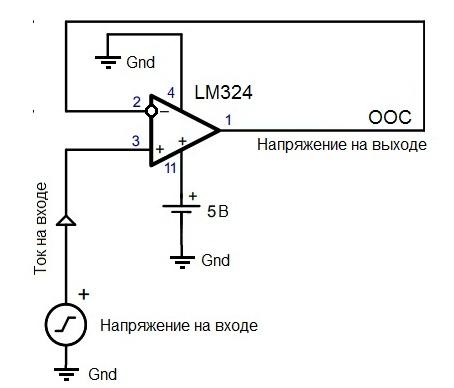 , 1997).
, 1997). др., 2009). Наконец, компьютерные исследования показали, что активность интернейронов PV / FS в индукции постоянной активности, а интернейронов CB / RS – в опосредовании устойчивости к дистракторам, которые могут преждевременно прекращать постоянную активность (Wang et al., 2004).
др., 2009). Наконец, компьютерные исследования показали, что активность интернейронов PV / FS в индукции постоянной активности, а интернейронов CB / RS – в опосредовании устойчивости к дистракторам, которые могут преждевременно прекращать постоянную активность (Wang et al., 2004).
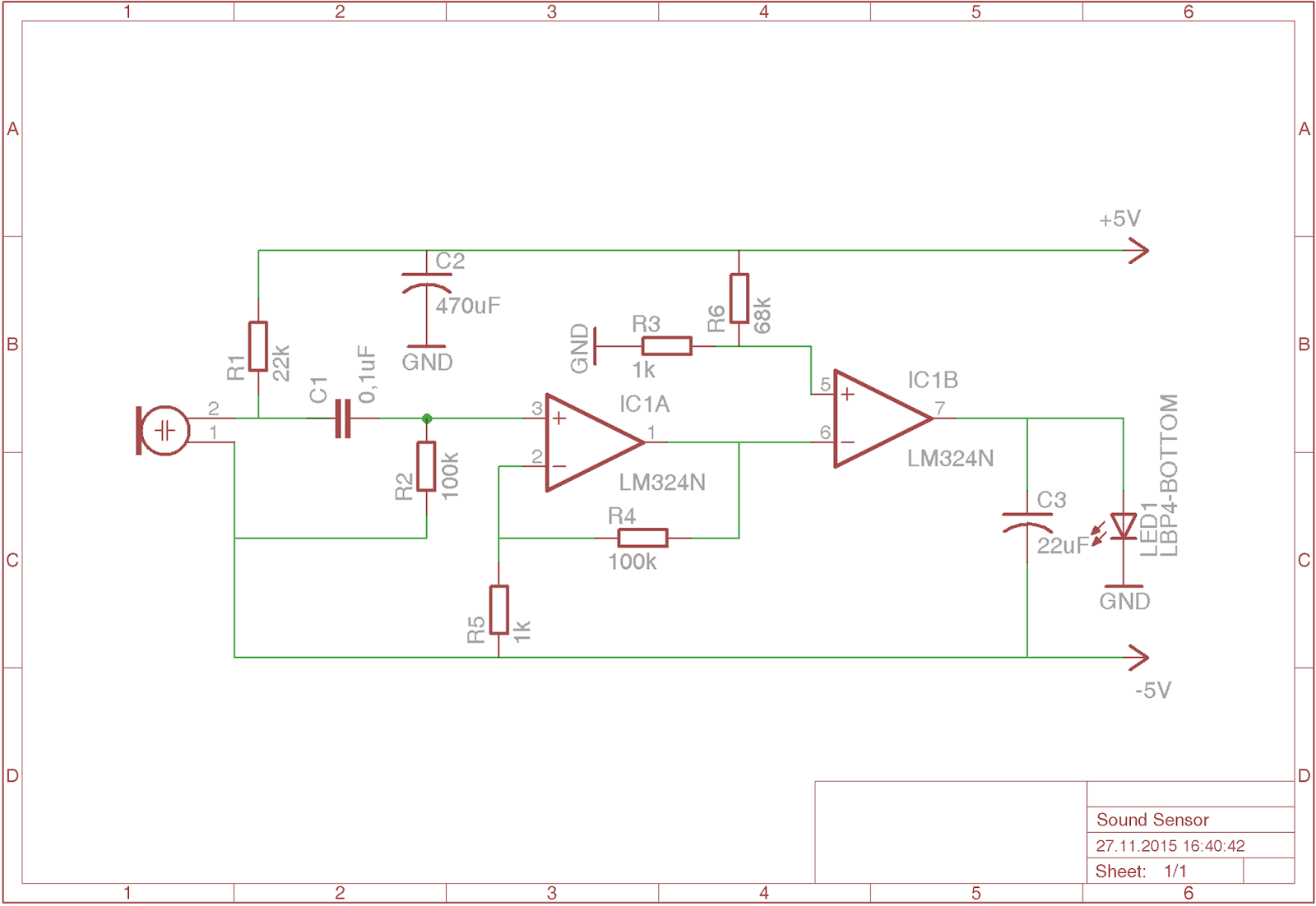 Они были объединены в сеть, которая включала 16 пирамидальных моделей и 4 модели интернейронов (2 модели FS, 1 модель RS и 1 модель IS).Связь между модельными нейронами была основана на экспериментальных анатомических и электрофизиологических данных, как описано ниже. Все модели реализованы в среде моделирования Neuron (Hines and Carnevale, 2001), а моделирование было выполнено на кластере xeon (8-ядерные процессоры xeon).
Они были объединены в сеть, которая включала 16 пирамидальных моделей и 4 модели интернейронов (2 модели FS, 1 модель RS и 1 модель IS).Связь между модельными нейронами была основана на экспериментальных анатомических и электрофизиологических данных, как описано ниже. Все модели реализованы в среде моделирования Neuron (Hines and Carnevale, 2001), а моделирование было выполнено на кластере xeon (8-ядерные процессоры xeon). (2004) (Таблица 1 и Рисунок 1). Размеры соматического, аксонного и дендритного компартментов пирамидной модельной клетки, а также пассивные и активные параметры модельного нейрона перечислены в дополнительном тексте (Дополнительные таблицы 1, 2).
(2004) (Таблица 1 и Рисунок 1). Размеры соматического, аксонного и дендритного компартментов пирамидной модельной клетки, а также пассивные и активные параметры модельного нейрона перечислены в дополнительном тексте (Дополнительные таблицы 1, 2). , 2013). Кроме того, каждый отдельный подтип модели интернейрона включает дополнительные ионные механизмы, которые, как известно, присутствуют в каждом типе (Toledo-Rodriguez et al., 2005), как подробно описано в следующих параграфах.
, 2013). Кроме того, каждый отдельный подтип модели интернейрона включает дополнительные ионные механизмы, которые, как известно, присутствуют в каждом типе (Toledo-Rodriguez et al., 2005), как подробно описано в следующих параграфах. Он ответил на деполяризующий импульс тока (0,05 нА, 500 мс) шестью пиками, как показано на рисунке 2A (вверху), с порогом потенциала действия -53 мВ. Деполяризующий ток 0.2 нА, 500 мс привели к отклику 100 Гц (10 всплесков 100 мс) (рисунок 2A, внизу).
Он ответил на деполяризующий импульс тока (0,05 нА, 500 мс) шестью пиками, как показано на рисунке 2A (вверху), с порогом потенциала действия -53 мВ. Деполяризующий ток 0.2 нА, 500 мс привели к отклику 100 Гц (10 всплесков 100 мс) (рисунок 2A, внизу). , 1997), показывает регулярный всплеск модели RS. Реакция модели нейрона IS на инъекции деполяризующего тока (C1) по сравнению с экспериментальными данными из записей токового зажима ( C2 , адаптировано из Cauli et al., 1997), выявляют нерегулярные всплески модели IS.
, 1997), показывает регулярный всплеск модели RS. Реакция модели нейрона IS на инъекции деполяризующего тока (C1) по сравнению с экспериментальными данными из записей токового зажима ( C2 , адаптировано из Cauli et al., 1997), выявляют нерегулярные всплески модели IS. 2В, вверху).
2В, вверху). , 1997) (рис. 2C).
, 1997) (рис. 2C). 7 мс и σ = 0,9 (Thomson, Lamy, 2007). Также были включены аутаптические контакты, которые были адаптированы к 1/3 возбуждающих связей (Lubke et al., 1996).
7 мс и σ = 0,9 (Thomson, Lamy, 2007). Также были включены аутаптические контакты, которые были адаптированы к 1/3 возбуждающих связей (Lubke et al., 1996). , 2003). Сводка синаптических связей, присутствующих в микросхеме, приведена в Таблице 5.
, 2003). Сводка синаптических связей, присутствующих в микросхеме, приведена в Таблице 5. et al., 2007; Wang et al., 2008; Wang, Gao, 2009). Проводимость одного синапса AMPA-R на модели пирамидного нейрона была отрегулирована так, чтобы он генерировал ответ напряжения 0.1 мВ в соме (Nevian et al., 2007). Ток NMDA был подтвержден протоколом фиксации напряжения, чтобы воспроизвести результаты Wang et al. (2008) (Рисунок 3A). Согласно Wang и Gao (2009), токи, опосредованные AMPA и NMDA, были зарегистрированы на уровне -70 мВ и +60 мВ, соответственно, в моделях нейронов FS и RS. Наши результаты соответствуют экспериментальным данным, как показано на рисунках 3B, C. Относительная доля синаптических компонентов, опосредованных рецепторами NMDA и AMPA, в моделях FS стандартизована на уровне 0.5 (Ван и Гао, 2009). Относительная доля синаптических компонентов, опосредованных рецепторами NMDA и AMPA, в моделях RS стандартизована на уровне 0,8 (Wang and Gao, 2009). Из-за отсутствия экспериментальных данных для модели нейрона IS, его AMPA- и NMDA-опосредованные токи также были смоделированы, чтобы соответствовать моделям нейронов FS и RS, тогда как соотношение NMDA-AMPA было адаптировано так, чтобы модель интернейрона IS могла потенциалы действия огня во время воздействия.
et al., 2007; Wang et al., 2008; Wang, Gao, 2009). Проводимость одного синапса AMPA-R на модели пирамидного нейрона была отрегулирована так, чтобы он генерировал ответ напряжения 0.1 мВ в соме (Nevian et al., 2007). Ток NMDA был подтвержден протоколом фиксации напряжения, чтобы воспроизвести результаты Wang et al. (2008) (Рисунок 3A). Согласно Wang и Gao (2009), токи, опосредованные AMPA и NMDA, были зарегистрированы на уровне -70 мВ и +60 мВ, соответственно, в моделях нейронов FS и RS. Наши результаты соответствуют экспериментальным данным, как показано на рисунках 3B, C. Относительная доля синаптических компонентов, опосредованных рецепторами NMDA и AMPA, в моделях FS стандартизована на уровне 0.5 (Ван и Гао, 2009). Относительная доля синаптических компонентов, опосредованных рецепторами NMDA и AMPA, в моделях RS стандартизована на уровне 0,8 (Wang and Gao, 2009). Из-за отсутствия экспериментальных данных для модели нейрона IS, его AMPA- и NMDA-опосредованные токи также были смоделированы, чтобы соответствовать моделям нейронов FS и RS, тогда как соотношение NMDA-AMPA было адаптировано так, чтобы модель интернейрона IS могла потенциалы действия огня во время воздействия.